白背飛虱細胞色素P450還原酶基因的分子特征與表達模式分析
2016-08-15 05:25:05余航劉蘇朱晴子周文武梁慶梅史肖肖祝梓杰祝增榮
浙江大學學報(農業與生命科學版) 2016年4期
余航, 劉蘇, 朱晴子, 周文武, 梁慶梅, 史肖肖, 祝梓杰, 祝增榮
(浙江大學昆蟲科學研究所/水稻生物學國家重點實驗室/農業部農業昆蟲學重點實驗室,杭州 310058)
?

白背飛虱細胞色素P450還原酶基因的分子特征與表達模式分析
余航, 劉蘇, 朱晴子, 周文武, 梁慶梅, 史肖肖, 祝梓杰, 祝增榮*
(浙江大學昆蟲科學研究所/水稻生物學國家重點實驗室/農業部農業昆蟲學重點實驗室,杭州 310058)
從水稻重要害蟲白背飛虱(Sogatellafurcifera)中克隆了一條編碼細胞色素P450還原酶(cytochrome P450 reductase,CPR)的全長cDNA,命名為SfCPR,其開放閱讀框全長為2 034 bp,編碼一個含有677個氨基酸殘基的蛋白質。SfCPR蛋白質的2級結構具有CPR的典型特征,例如N端跨膜區、黃素單核苷酸結合域、黃素腺嘌呤二核苷酸結合域和煙酰胺腺嘌呤二核苷酸磷酸結合域,以及保守的催化殘基。系統進化分析結果顯示,SfCPR與灰飛虱和褐飛虱的CPRs聚在同一分支。熒光定量聚合酶鏈式反應結果顯示:SfCPR基因在白背飛虱各齡期均有表達,其中以成蟲表達量最高;在白背飛虱成蟲各組織中均能夠檢測到SfCPR基因的表達,其中,以腹部表達量最高。使用亞致死劑量的溴氰菊酯、吡蟲啉和噻嗪酮處理白背飛虱3齡若蟲后,試蟲的SfCPR表達水平均顯著上升。上述結果為進一步研究該基因的生理功能奠定了基礎。
細胞色素P450還原酶; 白背飛虱; 殺蟲劑; 表達模式
Summary Nicotinamide adenine dinucleotide phosphate (NADPH)-cytochrome P450 reductase (CPR) is an electron supplier for various cytochrome P450 monooxygenases. Most P450-mediated catalytic reactions in insects require involvement of CPR, such as detoxification of insecticides and plant secondary metabolites. So far, CPR genes have been identified and characterized from many model and non-model insect species. Since insect P450 system is one of the most important metabolic adaptive traits involved in the degradation of xenobiotics and regulation of endogenous substrates. CPR, as the indispensable electron donor of P450 system, has attracted increasing attention as a potential candidate to develop novel chemical inhibitor to manage target insect pests. Rice planthoppers, such asNilaparvatalugens,LaodelphaxstriatellusandSogatellafurcifera, are considered to be the most serious pests of rice. Previously, some studies onL.striatellusandN.lugensfound that silencing the CPR gene by RNAi technology could increase their sensitivity to insecticides, but little was known aboutS.furcifera. This work firstly reports the identification of CPR gene inS.furciferaand up-regulation of the CPR transcription by chemical insecticides. The research will facilitate further study on the function of CPR inS.furcifera.
In this study, a full-length cDNA encoding CPR was cloned fromS.furcifera. The phylogenetic relationships ofSfCPRwith other insect CPRs were estimated by neighbor-joining method, and its distribution in various tissues and different developmental stages were analyzed by real-time quantitative polymerase chain reaction (qPCR). Finally, after exposure of deltamethrin, buprofezin and imidacloprid at sublethal concentrations for 6, 12 and 24 h, the relative expression levels ofSfCPRin the third-instar nymphs were investigated.
By searching the transcriptome data sets ofS.furcifera, a cDNA fragment encoding putative CPR (namedSfCPR) was identified. This cDNA fragment was then amplified by PCR and sequenced in order to confirm that the sequence was not chimeric. TheSfCPRcDNA contained a complete open reading frame (ORF) of 2 034 bp nucleotides, encoding a protein of 677 amino acid residues. The theoretical isoelectric point (pI) and calculated molecular mass of SfCPR protein are 5.48 and 76.762 ku, respectively. The secondary structure of SfCPR protein showed the marked features of typical CPRs, such as N-terminal transmembrane region, FMN-, FAD- and NADPH-binding domains and conserved catalytic residues. In addition, a transmembrane anchor was predicted in the N-terminus of the protein, indicating that SfCPR is an endoplasmic reticulum-located protein. Phylogenic analysis was used to gain insight into the phylogenetic relationships among CPR proteins from diverse insect species. We found that SfCPR and CPRs from other two planthoppers were clustered together. Real-time quantitative PCR showed that the expression ofSfCPRwas detectable in all developmental stages and the level in adults was the highest. TheSfCPRtranscripts were also expressed in all the tested tissues of the adults, and most were strongly expressed in the abdomen. The exposure at sublethal concentrations of deltamethrin, buprofezin and imidacloprid led to significantly elevated expression ofSfCPR. The expression ofSfCPRwas activated soon (6 h) after treatment with buprofezin and imidacloprid, while the response ofSfCPRexpression to deltamethrin was relatively slow (12 h).
In conclusion, this work is the first report about the cDNA sequence information, secondary structure and transcription profiles of CPR gene in theS.furcifera. These findings provide foundation for further research on the physiological function ofSfCPR.
煙酰胺腺嘌呤二核苷酸磷酸-細胞色素P450還原酶(cytochrome P450 reductase,CPR)是細胞色素P450單加氧酶(cytochrome P450 monooxygenases,CYP)的電子供體,在P450介導的諸多催化反應中起著非常重要的作用[1]。對于昆蟲而言,大多數P450介導的對殺蟲劑和植物次生代謝物的解毒過程中都需要有CPR的參與[2-3]。CPR也會參與昆蟲的其他生理途徑,例如蛻皮激素和表皮碳氫化合物的合成與代謝等[4-5]。
盡管昆蟲CPR的晶體結構目前尚無報道,但是對大鼠和酵母CPR的3維結構研究表明,該酶由3個功能結構域組成:黃素單核苷酸(flavin mononucleotide,FMN)結合域、黃素腺嘌呤二核苷酸(flavin adenine dinucleotide,FAD)結合域和煙酰胺腺嘌呤二核苷酸磷酸(nicotinamide adenine dinucleotide phosphate,NADPH)結合域,它們均參與電子穿梭過程[6-7]。此外,CPR在N端有一較短的疏水性跨膜區,該區域有助于CPR定位于細胞內質網上[2,6]。結構生物學研究表明,CPR在電子傳遞中自身結構會發生變化,以促進與NADPH的結合和電子轉移[8-9]。
目前,多種模式和非模式昆蟲的CPR基因已被鑒定,如黑腹果蠅(Drosophilamelanogaster)[5,10]、岡比亞按蚊(Anophelesgambiae)[11]、床虱(Cimexlectularius)[12]、二化螟(Chilosuppressalis)[13]、棉鈴蟲(Helicoverpaarmigera)[14]和小菜蛾(Plutellaxylostella)[15]等,這對深入研究CPR的生理功能和利用CPR來開發新型的害蟲無公害管理技術均具有重要意義。由于CPR是P450s的電子供體,所以目前利用RNA干擾技術(RNA interference,RNAi)沉默昆蟲的CPR基因進而抑制P450s的活性也受到越來越多的關注。例如:當注射雙鏈RNA(double-stranded RNA,dsRNA)沉默岡比亞按蚊和床虱的CPR基因后,試蟲對擬除蟲菊酯類殺蟲劑的敏感性均顯著上升[11-12];當黑腹果蠅的CPR基因被沉默后,試蟲表皮碳氫化合物合成代謝受阻,試蟲的存活率和羽化率均顯著下降[10]。
稻飛虱是水稻的一類重大害蟲。近10年來,褐飛虱(Nilaparvatalugens)、灰飛虱(Laodelphaxstriatellus)和白背飛虱(Sogatellafurcifera)及其傳播的病毒病相繼大規模爆發[16-18]。目前,防治這類害蟲仍然依賴大量使用化學殺蟲劑;而殺蟲劑的無節制使用,不僅造成環境污染,而且還誘使飛虱產生抗藥性[19]。此前,針對褐飛虱和灰飛虱CPR基因的研究發現,通過RNAi技術沉默該基因后,試蟲對殺蟲劑敏感性顯著上升,因此,可以利用該基因開發新型的無公害防治技術[20-21]。但白背飛虱的CPR基因至今尚無報道。
本研究首先克隆了重要水稻害蟲白背飛虱的CPR基因(SfCPR)全長cDNA序列,并對SfCPR基因的分子特征進行了分析;其次,使用實時熒光定量聚合酶鏈式反應(real-time quantitative polymerase chain reaction,qPCR)檢測了該基因在白背飛虱不同組織和不同發育階段的表達模式;最后,檢測了SfCPR基因在受到3種殺蟲劑(溴氰菊酯、吡蟲啉和噻嗪酮)脅迫后表達水平的變化。本研究結果可為進一步揭示SfCPR的生理功能奠定基礎。
1 材料與方法
1.1供試蟲源
本試驗所用白背飛虱種群采集自浙江大學華家池校區(杭州),用感蟲水稻品種TN1(Taichung Native 1)在人工氣候室內飼養繁殖35代以上。飼養條件:溫度(25±1) °C,相對濕度80%,光周期16 h光照/8 h黑夜。
1.2RNA提取和第1鏈cDNA合成
取白背飛虱3齡若蟲100頭,用Trizol試劑(美國Invitrogen公司)提取總RNA。提取的RNA樣本用無RNA酶的DNA酶(大連寶生物工程有限公司)處理,以消除可能的基因組DNA污染。RNA的濃度和質量用Nanodrop 2000微量分光光度計(美國Thermo Scientific公司)測定。取1 μg總RNA,使用PrimeScript試劑盒(大連寶生物工程有限公司)合成第1鏈cDNA。
1.3PCR擴增
通過搜索已公開的白背飛虱轉錄組數據庫[22],得到一條編碼CPR的cDNA序列,命名為SfCPR。根據SfCPR的序列信息設計1對基因特異性引物(表1),擴增其開放閱讀框(open reading frame,ORF)區域。使用KOD FX DNA聚合酶(日本東洋紡公司)進行PCR擴增。PCR產物經瓊脂糖凝膠電泳鑒定,用凝膠回收試劑盒(美國Axygen公司)回收純化,克隆入pMD18-T載體(大連寶生物工程有限公司),轉化大腸桿菌DH5α菌株,并測序驗證。
1.4序列分析
使用ExPASy服務器(http://www.expasy.org/tools/protparam.html)計算蛋白質理論分子質量和等電點。使用NCBI的BLAST服務器(http://blast.ncbi.nlm.nih.gov/Blast.cgi)搜索序列一致性及同源基因。使用TMHMM程序(http://www.cbs.dtu.dk/services/TMHMM)預測跨膜區。通過搜索NCBI的保守結構域數據庫(http://www.ncbi.nlm.nih.gov/structure/cdd/cdd.shtml)確定保守結構域及關鍵催化殘基。使用ClustalW 2.0程序(http://www.ebi.ac.uk/tools/msa/clustalw2/)連配SfCPR與其他昆蟲CPR的氨基酸序列,連配結果導入MEGA 5.05軟件[23],以鄰接法構建系統發育樹,各分支置信度通過自舉法進行1 000次重復檢驗獲取。
1.5實時熒光定量聚合酶鏈式反應
使用實時熒光定量聚合酶鏈式反應(qPCR)檢測SfCPR基因在白背飛虱不同發育階段(1~5齡若蟲和成蟲)以及短翅型成蟲不同組織(頭、胸、腹、足)中的相對表達水平。試驗設3個生物學重復,每個重復含試蟲50頭。提取總RNA的方法同1.2節,使用Revertra Ace qPCR 反轉錄試劑盒(日本東洋紡公司)合成第1鏈cDNA,并用超純水稀釋至10 ng/μL。qPCR擴增體系為20 μL,包含1 μL (10 ng) cDNA模板,10 μL SYBR Green Realtime PCR Master Mix試劑(日本東洋紡公司),上下游引物各0.2 μL (0.2 μmol/L),超純水8.6 μL。選擇白背飛虱的18S rRNA作為內參基因。qPCR所用引物見表1。qPCR所用儀器為Bio-Rad CFX96型實時定量系統(美國Bio-Rad Laboratories公司)。PCR反應采用2步法,熱循環參數:95 ℃,2 min,1個循環;95 ℃,10 s,60 ℃,20 s,40個循環。基因的相對表達水平使用2-ΔΔCT方法[24]計算。
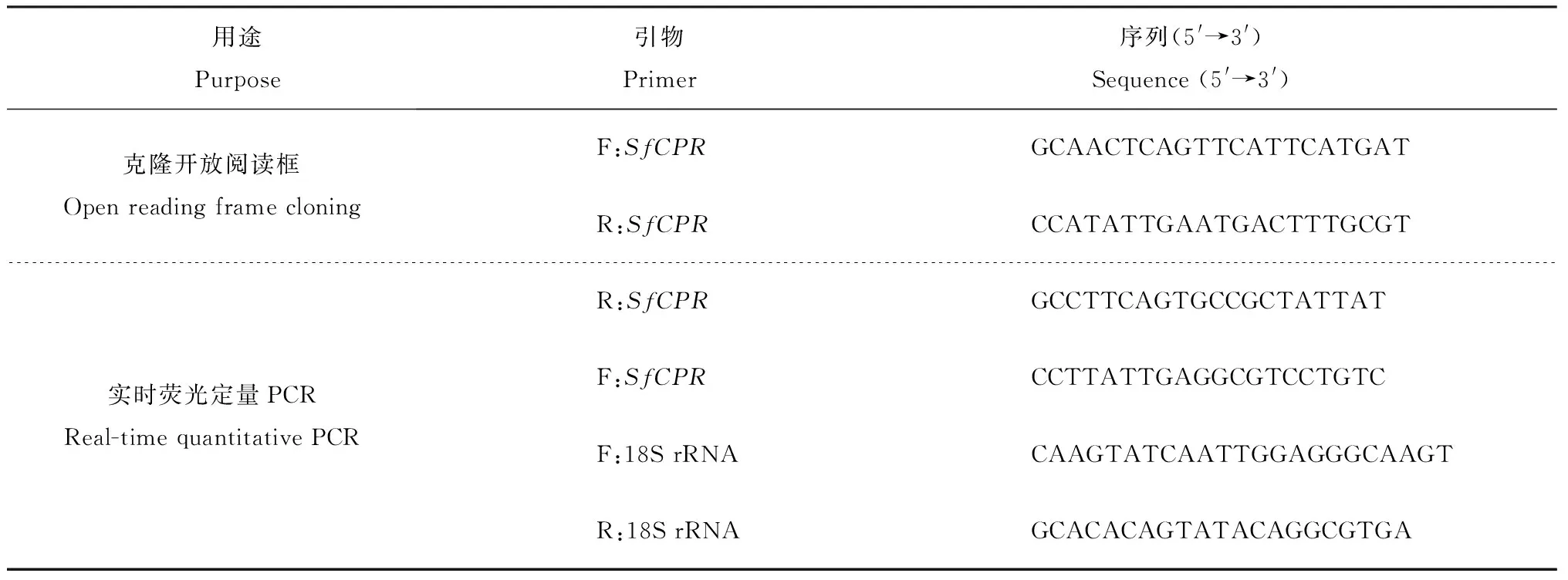
表1 研究所用引物
1.6殺蟲劑處理
溴氰菊酯、吡蟲啉和噻嗪酮購自浙江省臺州市新農公司,純度≥95%。使用時先用丙酮配制成儲存液,再逐級稀釋至亞致死濃度(溴氰菊酯、吡蟲啉和噻嗪酮均為0.5 ng/μL)。使用手動微量點滴儀(美國Hamilton公司)對白背飛虱3齡若蟲進行殺蟲劑處理,每試蟲滴定0.1 μL。處理6、12和24 h后,隨機選擇30頭試蟲提取RNA,檢測SfCPR基因表達水平的變化。以丙酮處理的試蟲作為對照。試驗設3個生物學重復。qPCR步驟同1.5節。
1.7數據統計
使用DPS V9.50軟件[25]對試驗數據進行統計分析。原始數據經反正弦轉換,用單因素方差分析進行處理,并用最小顯著差別法比較多個樣本之間是否存在顯著差異(顯著性水平設為P<0.05)。
2 結果
2.1序列分析
通過搜索白背飛虱轉錄組數據庫,獲得了1條編碼SfCPR的cDNA序列。該序列經PCR擴增和測序予以驗證。SfCPR的ORF區域全長為2 034 bp,編碼一個由677個氨基酸殘基組成的蛋白質。SfCPR的理論分子質量為76.762 ku,理論等電點為5.48。該蛋白質與半翅目昆蟲CPR的氨基酸序列具有非常高的一致性(68%~97%),與非半翅目昆蟲CPR的一致性也較高(65%~72%)(表2)。SfCPR序列已登錄至GenBank數據庫,登錄號為KJ017970。
為了確定白背飛虱是否還存在其他種類的CPR旁系同源基因,以模式昆蟲的CPR氨基酸序列為模板,使用本地BLAST程序在白背飛虱轉錄組數據庫中搜索,結果沒有發現新的與CPR同源的蛋白質序列。這說明在白背飛虱體內,在轉錄水平上只有1種CPR表達形式。為分析SfCPR蛋白質的2級結構特征,將SfCPR與其他幾種昆蟲的CPR進行連配。結果(圖1)表明,SfCPR具有CPR的典型特征,例如N端有一個由23個氨基酸殘基組成的跨膜區,保守的FMN、FAD和NADPH結合區域,以及關鍵催化殘基。
2.2系統進化分析
為了探知昆蟲CPRs之間的進化關系,從GenBank下載半翅目(Hemiptera)、膜翅目(Hymenoptera)、鞘翅目(Coleoptera)、鱗翅目(Lepidoptera)和雙翅目(Diptera)昆蟲已被注釋的CPR氨基酸序列,連同SfCPR一起構建系統發育樹。結果(圖2)顯示,雖然這些蛋白質的同源性很高,但不同物種的CPR仍被分在不同的進化支上。其中,白背飛虱(S.furcifera)、灰飛虱(L.striatellus)和褐飛虱(N.lugens)的CPRs聚在同一分支上:表明它們之間的親緣關系最接近。

表2 白背飛虱和其他昆蟲CPRs氨基酸序列的一致性百分數

Sfur:白背飛虱;Lstr:灰飛虱;Nlug:褐飛虱;Agam:岡比亞按蚊;Bmor:家蠶;Clec:床虱;Mdom:家蠅。實線方框內為預測的跨膜區;灰色、黃色和紫色分別表示構成FMN、FAD和NADPH結合域的關鍵氨基酸殘基;虛線框內為FAD結合域的精氨酸、酪氨酸和絲氨酸;黑色箭頭處為絲氨酸、半胱氨酸、天冬氨酸和色氨酸4個催化殘基。Sfur: Sogatella furcifera; Lstr: Laodelphax striatellus; Nlug: Nilaparvata lugens; Agam: Anopheles gambiae; Bmor: Bombyx mori; Clec: Cimex lectularius; Mdom: Musca domestica. The putative transmembrane region is boxed, and the amino acid residues constituting the binding site in the FMN-, FAD- and NADPH-binding domains are highlighted in gray, yellow and purple, respectively. FAD-binding motifs (Arg-X-Tyr-Ser) are indicated by dotted box, and four catalytic residues (Ser, Cys, Asp and Trp) are marked by black arrows. 圖1 白背飛虱和其他昆蟲的CPRs氨基酸多序列比對Fig.1 Multiple alignment of amino-acid sequences of CPRs from S. furcifera and other insect species

下劃線處為白背飛虱的CPR;各序列的GenBank登錄號見表2。標尺表示在不同昆蟲的CPR蛋白質中5%的氨基酸殘基有差異。 CPRs from S. furcifera are underlined. GenBank accession numbers of the sequences used are listed in Table 2. Scale represents 5% of the amino acid residues changed in the CPR proteins from different insect species.圖2 基于鄰接法的昆蟲CPRs系統進化分析Fig.2 Phylogenic analysis of insect CPRs based on neighbor-joining method
2.3組織和發育階段SfCPR表達模式分布
通過qPCR分析發現:SfCPR基因在白背飛虱短翅型成蟲的各組織(頭、胸、腹、足)中均有表達,其中以腹部的表達量最高,顯著高于頭部和足部(P<0.05),但與胸部表達水平間的差異無統計學意義(P>0.05);SfCPR基因在足部的表達水平最低(圖3A)。同樣,在白背飛虱的各個發育期均能檢測到SfCPR基因的表達,其中成蟲期的SfCPR表達量最高,2齡若蟲次之,4齡若蟲表達量最低,且4齡若蟲SfCPR的表達水平與2齡若蟲和成蟲之間差異有統計學意義(P<0.05)(圖3B)。
2.4殺蟲劑處理對SfCPR表達的影響
使用亞致死濃度的溴氰菊酯、吡蟲啉和噻嗪酮處理白背飛虱若蟲后,SfCPR基因的表達水平均有不同程度的上調(圖4):SfCPR對吡蟲啉和噻嗪酮的響應較迅速,使用這2種殺蟲劑處理6 h后,SfCPR的表達水平即顯著上升,較之丙酮處理組分別提高了16.04倍和5.72倍;但SfCPR對溴氰菊酯的響應較慢,處理12 h后,該基因的表達水平才顯著升高;用3種殺蟲劑處理24 h后,sfCPR基因的表達水平仍然顯著高于丙酮對照組。

短柵上的不同小寫字母表示在P<0.05水平差異有統計學意義。Different lowercase letters above bars indicate statistically significant differences at the 0.05 probability level. 圖3 SfCPR在白背飛虱不同組織(A)和不同發育階段(B)的相對表達水平Fig.3 Tissue-specific (A) and developmental stage-dependent (B) expression of S. furcifera CPR gene SfCPR
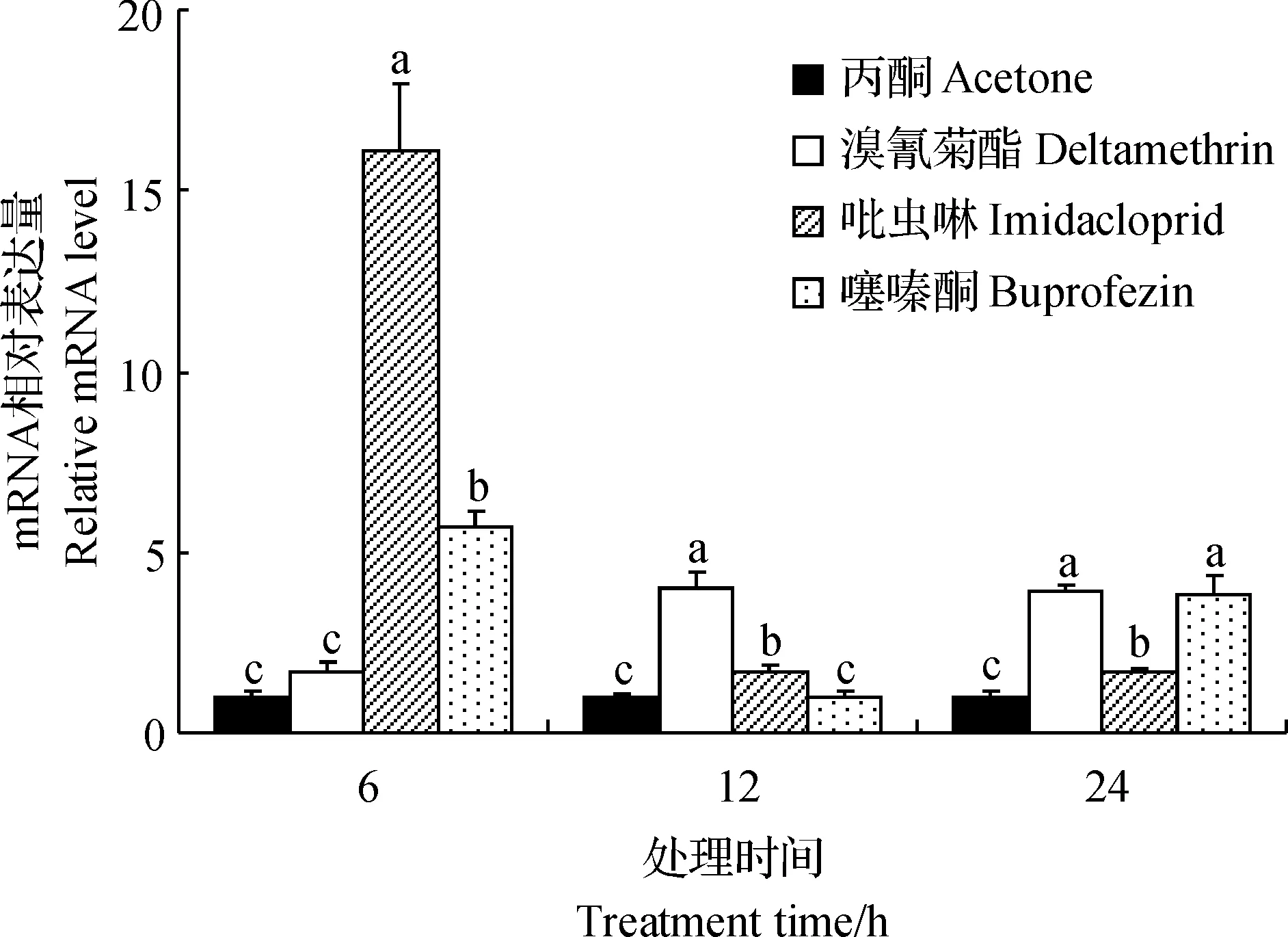
短柵上的不同小寫字母表示在P<0.05水平差異有統計學意義。Different lowercase letters above bars indicate statistically significant differences at the 0.05 probability level. 圖4 3種殺蟲劑滴定后SfCPR表達水平變化Fig.4 Relative mRNA levels of SfCPR after exposure of three insecticides
3 討論
本研究克隆了白背飛虱SfCPR的cDNA片段,分析了SfCPR的分子特征,使用qPCR檢測了SfCPR在白背飛虱不同組織、不同發育階段以及在受到不同殺蟲劑脅迫后的表達模式。CPR為細胞色素P450單加氧酶(CYPs)提供電子,在各種內源性和外源性化合物代謝過程中起著重要作用。本試驗結果為后續SfCPR功能研究以及利用該基因開發白背飛虱的新型防治技術奠定了基礎。
大多數昆蟲、脊椎動物和真菌基因組中只表達1個CPR基因。而在一些維管植物如水稻(Oryzasativa)和棉花(Gossypiumhirsutum)中表達2個以上的CPR基因[26-27]。本研究通過BLAST搜尋轉錄組數據,發現在白背飛虱轉錄組中只有1個CPR基因的轉錄本。這與在其他昆蟲上的研究結果[11-12]一致。
SfCPR蛋白質的2級結構具有CPR的典型特征,例如:保守的FMN、FAD和NADPH結合區域,這些區域在電子傳遞過程中有重要作用;N端的跨膜區,這一區域可將CPR蛋白錨定在細胞內質網上[6]。有研究表明,雖然N端跨膜區與CPR的活性無關,但這一區域若缺失,CPR就不能傳遞電子給CYP,催化反應也不能進行[2,28]。
多序列比對結果表明,SfCPR與其他昆蟲CPR的氨基酸序列一致性均較高,尤其是3種飛虱的CPR之間一致性超過95%。但同時也發現,在3種飛虱CPR的氨基酸序列中,FAD和NADPH結合區域之間的氨基酸殘基差異較大。哺乳動物CPR的晶體結構顯示,FMN和FAD結構域之間的連接區域與輔酶的定向有關,并且有助于穩定CPR的3維結構[9]。但FAD和NADPH結構域之間的連接區域是否具有同樣的功能,還有待進一步研究。
SfCPR基因在白背飛虱成蟲的不同組織中均能檢測到表達,其中以腹部表達量最高,足部最低。這與褐飛虱和灰飛虱中相應CPR基因的表達模式相似[20-21]。同樣,SfCPR在白背飛虱不同發育期也均有表達,其中以成蟲表達量最高,4齡若蟲最低。這一模式與褐飛虱和灰飛虱略有不同,可能是由于物種之間的差異所致[20-21]。此前針對棉鈴蟲和小菜蛾CPR的研究發現,該基因在殺蟲劑抗性種群中的表達水平顯著高于敏感種群,而且當試蟲受到殺蟲劑脅迫后,CPR基因的表達量均顯著上升。本研究使用3種不同作用類型的殺蟲劑處理白背飛虱若蟲后,SfCPR基因的表達水平同樣有不同程度的上調。其中,SfCPR對吡蟲啉和噻嗪酮的響應較快,處理后6 h表達水平即顯著上升;相較而言,SfCPR對溴氰菊酯的響應較慢,處理后12 h表達水平才顯著上調。這是首次發現飛虱科昆蟲的CPR基因對殺蟲劑脅迫產生響應,也暗示了SfCPR基因很可能參與了P450介導的對殺蟲劑的解毒過程。此前的報道顯示,注射dsRNA沉默褐飛虱和灰飛虱的CPR基因后,試蟲對殺蟲劑的敏感性顯著增加[20-21]。因此,本研究結果也為利用SfCPR開發針對白背飛虱的新型防治技術奠定了前期基礎。
[1]FEYEREISEN R. Insect P450 enzymes.AnnualReviewofEntomology, 1999,44:507-533.
[2]ANDERSEN J F, UTERMOHLEN J G, FEYEREISEN R. Expression of housefly CYP6A1 and NADPH-cytochrome P450 reductase inEscherichiacoliand reconstitution of an insecticide-metabolizing P450 system.Biochemistry, 1994,33:2171-2177.
[3]WEN Z, PAN L, BERENBAUM M R,etal. Metabolism of linear and angular furanocoumarins byPapiliopolyxenesCYP6B1 co-expressed with NADPH cytochrome P450 reductase.InsectBiochemistryandMolecularBiology, 2003,33:937-947.
[4]HORIKE N, TAKEMORI H, NONAKA Y,etal. Molecular cloning of NADPH-cytochrome P450 oxidoreductase from silkworm eggs.EuropeanJournalofBiochemistry, 2000,267:6914-6920.
[5]QIU Y, TITTIGER C, WICKER-THOMAS C,etal. An insect-specific P450 oxidative decarbonylase for cuticular hydrocarbon biosynthesis.ProceedingsoftheNationalAcademyofSciencesoftheUSA, 2012,109:14858-14863.
[6]WANG M, ROBERTS D L, PASCHKE R,etal. Three-dimensional structure of NADPH-cytochrome P450 reductase: Prototype for FMN- and FAD-containing enzymes.ProceedingsoftheNationalAcademyofSciencesoftheUSA, 1997,94:8411-8416.
[7]LAMB D C, KIM Y, YERMALITSKAYA L V,etal. A second FMN binding site in yeast NADPH-cytochrome P450 reductase suggests a mechanism of electron transfer by diflavin reductases.Structure, 2006,14:51-61.
[8]ELLIS J, GUTIERREZ A, BARSUKOV I L,etal. Domain motion in cytochrome P450 reductase: Conformational equilibria revealed by NMR and small-angle X-ray scattering.JournalofBiologicalChemistry, 2009,284:36628-36637.
[9]XIA C, HAMDANE D, SHEN A L,etal. Conformational changes of NADPH-cytochrome P450 oxidoreductase are essential for catalysis and cofactor binding.JournalofBiologicalChemistry, 2011,286:16246-16260.
[10]HOVEMANN B T, SEHLMEYER F, MALZ J.DrosophilamelanogasterNADPH-cytochrome P450 oxidoreductase: Pronounced expression in antennae may be related to odorant clearance.Gene, 1997,189:213-219.
[11]LYCETT G J, MCLAUGHLIN L A, RANSON H,etal.AnophelesgambiaeP450 reductase is highly expressed in oenocytes andinvivoknockdown increases permethrin susceptibility.InsectMolecularBiology, 2006,15:321-327.
[12]ZHU F, SAMS S, MOURAL T,etal. RNA interference of NADPH-cytochrome P450 reductase results in reduced insecticide resistance in the bed bug,Cimexlectularius.PLoSONE, 2012,7:e31037.
[13]LIU S, LIANG Q M, HUANG Y J,etal. Cloning, functional characterization, and expression profiles of NADPH-cytochrome P450 reductase gene from the Asiatic rice striped stem borer,Chilosuppressalis(Lepidoptera: Pyralidae).ComparativeBiochemistryandPhysiologyPartB:Biochemistry&MolecularBiology, 2013,166:225-231.
[14]ZHAO C, TANG T, FENG X,etal. Cloning and characterisation of NADPH-dependent cytochrome P450 reductase gene in the cotton bollworm,Helicoverpaarmigera.PestManagementScience, 2014,70:130-139.
[15]CHEN X E, ZHANG Y L. Identification and characterization of NADPH-dependent cytochrome P450 reductase gene and cytochrome b5gene fromPlutellaxylostella: Possible involvement in resistance to beta-cypermethrin.Gene, 2015,558(2):208-214.
[16]程家安,朱金良,祝增榮,等.稻田飛虱災變與環境調控.環境昆蟲學報,2008,30(2):176-182.
CHENG J A, ZHU J L, ZHU Z R,etal. Rice planthopper outbreak and environment regulation.JournalofEnvironmentalEntomology, 2008,30(2):176-182. (in Chinese with English abstract)
[18]祝增榮,程家安.中國水稻害蟲治理對策的演變及其展望.植物保護,2013,39(5):25-32.
ZHU Z R, CHENG J A. The evolution and perspective of rice insect pest management strategy in China.PlantProtection, 2013,39(5):25-32. (in Chinese with English abstract)
[19]婁永根,程家安.稻飛虱災變機理及可持續治理的基礎研究.應用昆蟲學報,2011,48(2):231-238.LOU Y G, CHENG J A. Basic research on the outbreak mechanism and sustainable management of rice planthoppers.ChineseJournalofAppliedEntomology, 2011,48(2):231-238. (in Chinese with English abstract)
[20]LIU S, LIANG Q M, ZHOU W W,etal. RNA interference of NADPH-cytochrome P450 reductase of the rice brown planthopper,Nilaparvatalugens, increases susceptibility to insecticides.PestManagementScience, 2015,71(1):32-39.
[21]ZHANG Y L, WANG Y M, WANG L H,etal. Knockdown of NADPH-cytochrome P450 reductase results in reduced resistance to buprofezin in the small brown planthopper,Laodelphaxstriatellus(Fallén).PesticideBiochemistryandPhysiology, 2015,127:21-27.
[22]XU Y, ZHOU W W, ZHOU Y J,etal. Transcriptome and comparative gene expression analysis ofSogatellafurcifera(Horváth) in response to southern rice black-streaked dwarf virus.PLoSONE, 2012,7:e36238.
[23]TAMURA K, PETERSON D, PETERSON N,etal. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods.MolecularBiologyandEvolution, 2011,28:2731-2739.
[24]LIVAK K J, SCHMITTGEN T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCTmethod.Methods, 2001,25:402-408.
[25]TANG Q Y, ZHANG C X. Data processing system (DPS) software with experimental design, statistical analysis and data mining developed for use in entomological research.InsectScience, 2013,20:254-260.
[26]YANG C Q, LU S, MAO Y B,etal. Characterization of two NADPH: Cytochrome P450 reductases from cotton (Gossypiumhirsutum).Phytochemistry, 2010,71:27-35.
[27]PARK S, KIM Y S, RUPASINGHE S G,etal. Rice P450 reductases differentially affect P450-mediated metabolism in bacterial expression systems.BioprocessandBiosystemsEngineering, 2013,36:325-331.
[28]LAMB D C, WARRILOW A G S, VENKATESWARLU K,etal. Activities and kinetic mechanisms of native and soluble NADPH-cytochrome P450 reductase.BiochemicalandBiophysicalResearchCommunications, 2001,286:48-54.
Molecular characterization and expression profiles of cytochrome P450 reductase gene in Sogatella furcifera (Hemiptera: Delphacidae).JournalofZhejiangUniversity(Agric. &LifeSci.), 2016,42(4):391-400
YU Hang, LIU Su, ZHU Qingzi, ZHOU Wenwu, LIANG Qingmei, SHI Xiaoxiao, ZHU Zijie, ZHU Zengrong*
(StateKeyLaboratoryofRiceBiology/KeyLaboratoryofAgriculturalEntomology,MinistryofAgriculture/InstituteofInsectSciences,ZhejiangUniversity,Hangzhou310058,China)
cytochrome P450 reductase;Sogatellafurcifera; insecticide; expression profile
農業部公益性行業(農業)科研專項(201403030);國家自然科學基金(31371935);國家重點基礎研究發展計劃(973計劃)(2010CB126200);亞洲發展銀行-國際水稻研究所(ADB-IRRI)稻飛虱項目(RETA13,RETA14,RETA15)。
Corresponding author):祝增榮(http://orcid.org/0000-0002-3247-1486),Tel:+86-571-88982355,E-mail:zrzhu@zju.edu.cn
聯系方式:余航(http://orcid.org/0000-0002-6651-0797),E-mail:hangyubig@qq.com
2015-12-17;接受日期(Accepted):2016-03-02;網絡出版日期(Published online):2016-07-18
S 435.112.3; Q 963
A
URL:http://www.cnki.net/kcms/detail/33.1247.S.20160718.2038.016.html
