Helicobacter suis感染小鼠模型的建立及其意義
2016-08-24 11:26:17楊若明田字彬于亞男李曉宇張帥慶
中國人獸共患病學報 2016年7期
楊若明,楊 林,田字彬,于亞男,毛 濤,李曉宇,張帥慶
?
Helicobacter suis感染小鼠模型的建立及其意義
楊若明,楊林,田字彬,于亞男,毛濤,李曉宇,張帥慶
目的建立Helicobactersuis(H.suis)感染小鼠模型并探究其意義。方法將40只雌性C57BL/6小鼠隨機分為模型組和對照組,模型組給予0.5 mL豬胃黏膜勻漿液(經(jīng)PCR檢測確定存在H.suis)灌胃,對照組給予0.5 mL PBS溶液灌胃。灌胃1個月及3個月后,各處死兩組中半數(shù)小鼠,取其胃黏膜組織,分別行PCR、HE染色檢測H.suis在小鼠胃內(nèi)的定植與胃黏膜淋巴濾泡形成情況。結(jié)果灌胃所用豬胃黏膜勻漿液中存在H.suis;H.suis可以在小鼠胃內(nèi)定植,并且3個月后仍存在;感染H.suis1個月及3個月后的小鼠胃黏膜均有淋巴濾泡的形成,并且后者的淋巴濾泡較前者明顯增大。結(jié)論成功建立了H.suis感染C57BL/6小鼠模型,H.suis可以誘導胃黏膜淋巴濾泡的形成,可能在胃黏膜相關(guān)淋巴組織(MALT)淋巴瘤的發(fā)生過程中發(fā)揮重要作用。
H.suis;豬胃;動物模型;淋巴濾泡
Supported by the National Natural Science Foundation of China (No. 81572320)
海爾曼螺桿菌(Helicobacterheilmannii,Hh)是除幽門螺桿菌(Helicobacterpylori,H.pylori)外最常見的定植于人類胃黏膜的螺旋桿菌,通過16Sr RNA基因序列分析可將其分為兩型[1]:其中感染人類的大部分為1型(約占78.5%),為單獨的菌種,與豬胃黏膜中分離得到的豬螺桿菌(Helicobactersuis,H.suis)有高度的同源性,可視為同一菌種[2];2型代表了定植于貓與犬胃內(nèi)的一組細菌,包括H.felis,H.bizzozeronii,H.salomonis,H.cynogastricus,H.baculiformis和H.heilmanniisp.nov等。H.suis能自然定植于人和諸多動物胃內(nèi)并具有致病性,為人獸共患病病原菌。國外研究已發(fā)現(xiàn),來源于食蟹猴胃內(nèi)的H.suis可穩(wěn)定定植于小鼠胃內(nèi),感染6個月后可使絕大多數(shù)小鼠胃中出現(xiàn)MALT淋巴瘤特征性的淋巴上皮病變(lymphoepithelial lesion, LEL)[3]。因此H.suis可作為H.pylori研究的替代工具,用于闡明螺桿菌與特異宿主之間的相互作用關(guān)系,為研究胃MALT淋巴瘤的發(fā)病機制提供了一個重要途徑。由于H.suis在豬胃中的自然感染率較高(可達60%以上)[2,4],且該菌在體外生長環(huán)境特殊,營養(yǎng)要求高,難以穩(wěn)定培養(yǎng),因此本研究擬從自然感染的豬胃中獲得H.suis并感染小鼠建立動物模型,為螺桿菌致病機制的研究提供理論基礎(chǔ)。
1 材料和方法
1.1實驗動物8周齡雌性SPF級C57BL/6野生型小鼠40只,體質(zhì)量約18~20 g,購自濟南朋悅實驗動物繁育有限公司,飼養(yǎng)于青島大學附屬醫(yī)院動物實驗室(SPF級),溫度20 ~22 ℃,濕度60%~70%,每日照明12 h(8∶00-20∶00),飼料及飲水均經(jīng)高溫高壓滅菌處理。
1.2細菌的獲取10只豬胃購自青島市某屠宰場,用無菌大剪刀沿胃大彎剪開,以自來水仔細沖洗干凈后,用無菌載玻片分別刮取10只豬胃充血發(fā)紅炎性病變區(qū)域的粘液層及表層胃黏膜組織,然后各自收集于無菌離心管中,再加入適量無菌磷酸鹽緩沖液(phosphate buffer saline, PBS),充分研磨勻漿后各自定容為2.0 mL備用。
1.3主要試劑組織DNA提取試劑盒購自北京天根生化科技有限公司,PCR相關(guān)試劑購自大連寶生物工程有限公司。
1.4豬胃中H.suis的檢測各自取10只豬胃黏膜勻漿液0.5 mL置于1.5 mL離心管中,用組織DNA提取試劑盒提取總DNA(提取方法參照說明書)。PCR引物序列見表1,由上海生工生物工程股份有限公司合成。采用25 μL反應體系,每管依次加入Premix Tap酶12.5 μL、無菌雙蒸水8.5 μL、dNTP 2.0 μL及上下游引物各1.0 μL。H.suis16S rRNA引物、H.pylori16S rRNA引物及螺桿菌多重引物分別用于特異性檢測H.suis、H.pylori及其他常見螺桿菌 (H.felis,H.bizzozeronii和H.Salomonis)的存在。94 ℃預變性2 min后,以下條件PCR擴增:94 ℃ 2 min,55 ℃ 30 s,72 ℃ 1 min,35個循環(huán),72 ℃再延伸1 min。PCR反應陽性對照以已經(jīng)鑒定為H.suis的DNA為模板,空白對照以無菌雙蒸水替代模板。然后用2%瓊脂糖凝膠電泳檢測PCR產(chǎn)物,確定H.suis及其他主要螺桿菌的存在情況。
表1本研究中所用的PCR引物和擴增產(chǎn)物大小
Tab.1PCR primers used in this study and the PCR product sizes
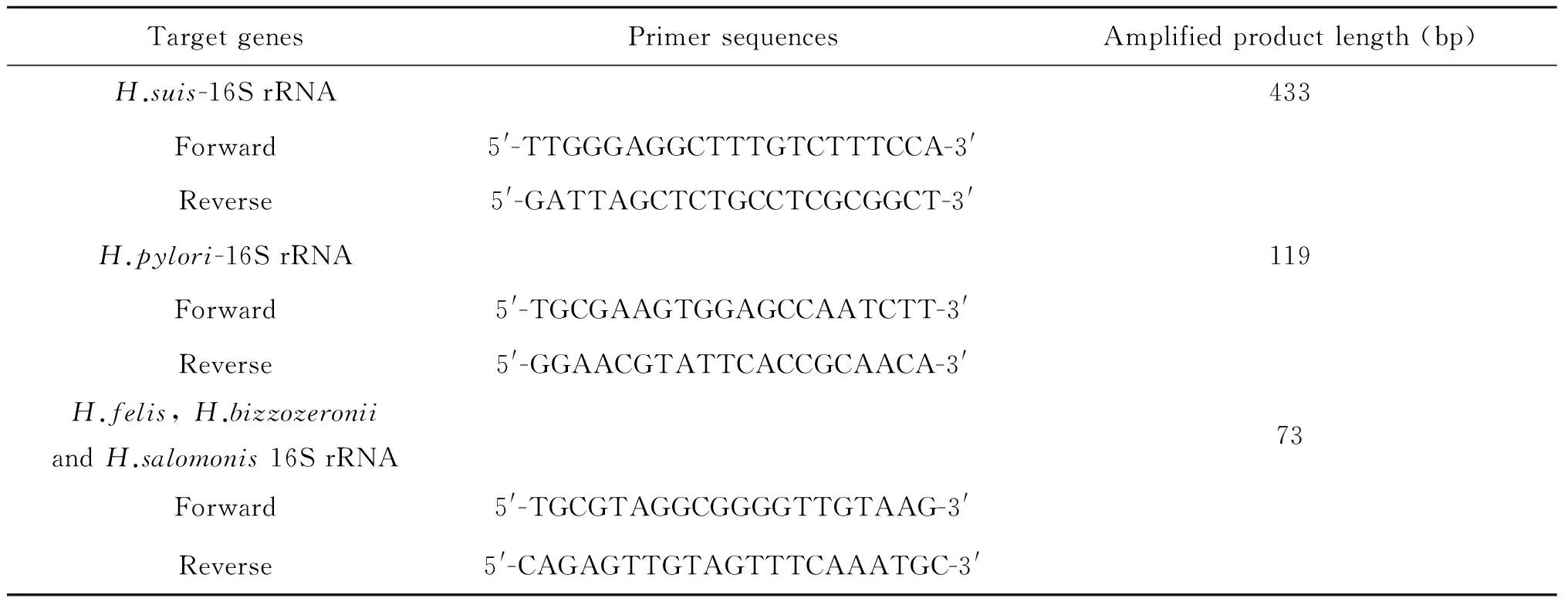
TargetgenesPrimersequencesAmplifiedproductlength(bp)H.suis-16SrRNA433Forward5'-TTGGGAGGCTTTGTCTTTCCA-3'Reverse5'-GATTAGCTCTGCCTCGCGGCT-3'H.pylori-16SrRNA119Forward5'-TGCGAAGTGGAGCCAATCTT-3'Reverse5'-GGAACGTATTCACCGCAACA-3'H.felis,H.bizzozeroniiandH.salomonis16SrRNA73Forward5'-TGCGTAGGCGGGGTTGTAAG-3'Reverse5'-CAGAGTTGTAGTTTCAAATGC-3'
1.5感染模型構(gòu)建C57BL/6小鼠隨機分為模型組和對照組,每組各20只。小鼠先適應性喂養(yǎng)1周,禁食禁水4 h。將6只經(jīng)PCR檢測證明存在H.suis的豬胃黏膜勻漿液混勻后,給予模型組每只小鼠0.5 mL豬胃黏膜勻漿液灌胃,對照組每只小鼠接受0.5 mL無菌PBS溶液灌胃,灌胃后禁食禁水4 h。
1.6小鼠胃組織的處理 各組小鼠灌胃1個月及3個月后,分別處死半數(shù)小鼠,取全胃沿胃大彎剪開,以無菌PBS溶液沖洗胃內(nèi)容物;取一半胃組織刮取黏膜層用于提取DNA行PCR反應。另一半胃組織經(jīng)10%甲醛固定后石蠟包埋,10 μm連續(xù)切片,行蘇木精-伊紅(HE)染色后檢測小鼠胃黏膜淋巴濾泡形成情況。
2 結(jié) 果
2.1豬胃黏膜中H.suis的存在情況分別從10只豬胃黏膜勻漿液中提取DNA,以H.suis特異性16S rRNA引物行PCR,其中6只豬胃可擴增出433 bp的目的條帶,其余4只豬胃及空白對照組的檢測結(jié)果為陰性,未擴增出相應條帶(見圖1)。以H.pylori特異性16S rRNA引物和其他螺桿菌 (H.Felis,H.bizzozeronii和H.Salomonis) 多重引物及無菌雙蒸水行空白對照檢測,其結(jié)果均為陰性,未擴增出相應條帶。以上結(jié)果表明用于建立感染模型的豬胃中只存在H.suis這一常見的致病性螺桿菌。本實驗中H.suis在豬胃中的感染率高達60%,與相關(guān)文獻報道基本一致[2,4]。
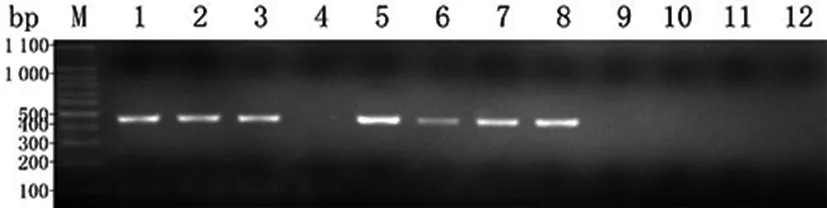
H. suis 16S rRNA gene by PCR amplification; M: DNA marker; 1: Positive control, H. suis; 2,3,4,5,6,7,8,9,10,11: pig gastric mucosa; 12: blank control, Sterile double distilled water. 圖1 10只豬胃黏膜中H. suis的鑒定結(jié)果Fig.1 Identification of H. suis in pig gastric mucosa
2.2小鼠胃黏膜中H.suis定植情況造模組小鼠接受豬胃黏膜勻漿液灌胃1個月與3個月后,分別取胃黏膜勻漿提取DNA,以H.suis特異性16S rRNA引物行PCR,均能擴增出約433 bp的目的條帶。以H.pylori特異性16S rRNA引物和其他螺桿菌 (H.felis,H.bizzozeronii和H.Salomonis) 多重引物及無菌雙蒸水行空白對照檢測,其結(jié)果均為陰性,未擴增出相應條帶(圖2、3)。表明來源于豬黏膜的H.suis可在小鼠胃內(nèi)定植,3個月后仍穩(wěn)定存在,未被清除。
2.3小鼠胃黏膜病理組織學變化小鼠感染H.suis1個月及3個月后,其胃黏膜中均檢測到淋巴濾泡形成,且3個月時淋巴濾泡的大小較1個月時明顯增大。而對照組未檢測到胃淋巴濾泡的形成 (圖4)。
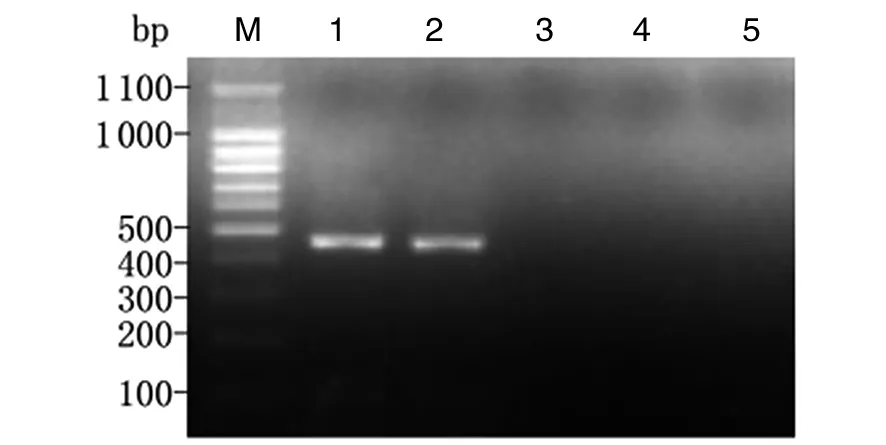
1, 2: H. suis 16S rRNA gene by PCR amplification; 3: H. pylori 16S rRNA gene by PCR amplification; 4: H. felis, H. bizzozeronii and H. salomonis special 16S rRNA gene by PCR amplification. M: DNA marker; 1: Positive control, H. suis; 2,3,4: mice gastric mucosa; 5: blank control, sterile double distilled water.圖2 感染H. suis 1個月后小鼠胃黏膜中H. suis的鑒定結(jié)果Fig.2 Identification of H. suis in the gastric mucosa of mice at 1 month postinfection
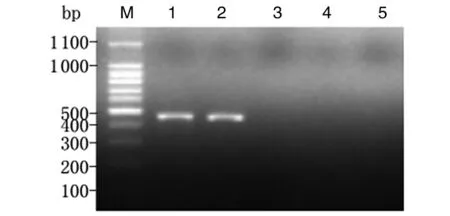
1, 2: H. suis 16S rRNA gene by PCR amplification; 3: H. pylori 16S rRNA gene by PCR amplification; 4: H. felis, H. bizzozeronii and H. salomonis special 16S rRNA gene by PCR amplification. M: DNA marker; 1: Positive control, H. suis; 2, 3, 4: mice gastric mucosa; 5: blank control, sterile double distilled water.圖3 感染H.suis 3個月后小鼠胃黏膜中H. suis的鑒定結(jié)果Fig.3 Identification of H. suis in the gastric mucosa of mice at 3 months after H. suis infection
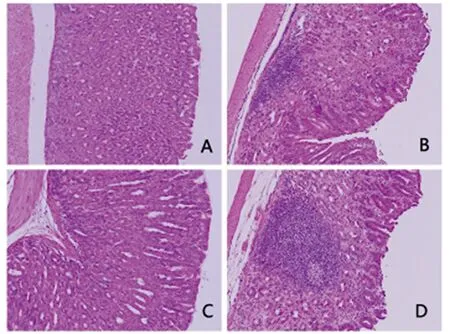
A: control group at 1 month after PBS solution treatment;B: model group at 1 month after H. suis infection;C: control group at 3 months after PBS solution treatment;D: model group at 3 months after H. suis infection.圖4 各組小鼠胃黏膜組織病理學表現(xiàn)(HE 染色,×200)Fig.4 Pathological changes in the gastric mucosa of mice in each group (HE staining, ×200)
3 討 論
H.pylori感染與慢性胃炎、消化性潰瘍、胃癌以及胃MALT淋巴瘤的發(fā)病密切相關(guān)[5]。為了探究其發(fā)病機制,國內(nèi)外學者對H.pylori感染動物模型進行了大量的研究,但未經(jīng)改良的H.pylori菌株難以長期定植于動物胃內(nèi),極大阻礙了H.pylori發(fā)病機制的進一步研究。Kim等人研究發(fā)現(xiàn)感染H.pylori80周的小鼠只是誘導出增生性胃炎、慢性萎縮性胃炎等癌前病變,并未出現(xiàn)胃癌[6],這說明單純H.pylori感染致癌的周期可能很長。而作為胃內(nèi)的第2種螺桿菌,H.suis可穩(wěn)定定植于小鼠胃內(nèi),較H.pylori更適用于胃MALT淋巴瘤發(fā)病機制的研究。H.suis感染后誘發(fā)的胃黏膜免疫反應以Th1為主[7],而Th1型免疫反應在C57BL/6小鼠中占優(yōu)勢,且此類小鼠其遺傳背景明確、個體差異小、價格便宜、操作簡單,各種處理因素容易控制,故本實驗選用C57BL/6小鼠為造模對象。
既往國內(nèi)外大部分海爾曼螺桿菌的研究都集中于個案報道和流行病學調(diào)查。有學者報道經(jīng)胃鏡檢查可從患者胃粘膜活檢標本中獲得海爾曼螺桿菌,進而感染小鼠建立模型。但由于海爾曼螺桿菌(Hh)在人群中感染率較低(我國約為5%~11%)[8],因此通過內(nèi)鏡獲取該菌較為困難。另外,Hh有著廣泛的動物宿主,目前在豬、犬、貓、鼠、雪貂及包括人類的靈長類動物胃內(nèi)均可檢測出Hh[9]。感染者多與家畜與寵物,如豬、犬、貓等有密切接觸史,這些動物可作為Hh傳播的貯存庫,人類與其直接接觸是導致感染Hh的危險因素[10-11]。有研究報道屠宰后的豬胃中60%以上可檢測到H.suis[2,4],其在生豬肉中可持續(xù)存活48 h以上,而在水中可存活4 d以上,因此接觸被污染的生豬肉及水源可能也是感染H.suis的途徑[12],豬胃有望成為獲取該菌的重要來源。相關(guān)報道表明采用PCR法檢測H.suis16S rRNA證實該菌存在的敏感性和特異性均優(yōu)于免疫組化、尿素酶試驗及姬姆薩染色等方法[13],本實驗采用PCR法在大部分豬胃及感染后 1個月與3個月的小鼠胃粘膜中均檢測到H.suis的存在,表明H.suis在豬胃中自然感染率較高,且未合并其他常見致病性螺桿菌的感染。H.suis可長期穩(wěn)定定植于小鼠胃內(nèi),保證了建立小鼠感染模型的可行性。
螺桿菌感染胃黏膜后可激活上皮細胞和固有免疫細胞,誘導炎癥因子的產(chǎn)生,作用于間質(zhì)細胞使其分泌趨化因子,募集淋巴細胞形成獲得性的MALT。H.suis可以直接引起慢性炎癥,也可以通過驅(qū)動胃上皮細胞壞死,導致細胞內(nèi)容物的釋放,進而間接促進炎癥[14]。慢性炎癥的長期刺激可導致淋巴細胞的克隆生長,形成以B細胞為主的MALT淋巴瘤[15]。在人類,H.suis主要定植于胃竇部, 位于黏液層、上皮細胞表面和胃小凹深部;而在本研究中我們發(fā)現(xiàn),感染H.suis1個月后,小鼠胃竇粘膜層及粘膜下層有大量淋巴細胞聚集,形成了淋巴濾泡,且其大小隨著感染時間的延長而增加,說明該模型可模擬人類感染H.suis后的病理變化。目前胃MALT淋巴瘤的發(fā)病率有逐年增高的趨勢,深入研究Helicobacter感染后MALT形成的機制并進行相應的干預,對預防胃MALT淋巴瘤的發(fā)生具有重要意義。
綜上所述,H.suis感染C57BL/6小鼠模型的建立可成功誘導胃黏膜淋巴濾泡的形成,為我們進一步研究胃MALT淋巴瘤的發(fā)病機制奠定了基礎(chǔ)。雖然既往有H.suis分離成功的個例報道[16],但多數(shù)學者在相同條件下重復試驗均告失敗,無法進行鑒定。故目前認為H.suis在體外難以穩(wěn)定培養(yǎng);本研究所用的細菌也來自豬胃,未經(jīng)純化。因此,為更好的研究H.suis在消化系統(tǒng)疾病中的具體致病機理,成功體外分離培養(yǎng)該菌并獲取純化菌株是今后努力的方向。
[1] O’Rourke JL, Solnick JV, Neilan BA, et al. Description of ‘CandidatusHelicobacterheilmannii’ based on DNA sequence analysis of 16S rRNA and urease genes[J].Int J Syst Evol Microbiol, 2004, 54(Pt 6): 2203-2211. DOI: 10.1099/ijs.0.63117-0
[2] De Groote D, van Doorn LJ, Ducatelle R, et al. ‘CandidatusHelicobactersuis’, a gastric helicobacter from pigs, and its phylogenetic relatedness to other gastrospirilla[J].Int J Syst Bacteriol, 1999, 49(Pt 4): 1769-1777.
[3] Nakamura M, Murayama SY, Serizawa H, et al. “CandidatusHelicobacterheilmannii” from a cynomolgus monkey induces gastric mucosa-associated lymphoid tissue lymphomas in C57BL/6 mice[J]. Infect Immun, 2007, 75(3): 1214-1222. DOI: 10.1128/ial.01459-06
[4] Baele M, Decostere A, Vandamme P, et al. Isolation and characterization ofHelicobactersuissp. nov. from pig stomachs[J]. Int J Syst Evol Microbiol, 2008, 58(6): 1350-1358. DOI: 10.1099/ijs.0.65133-0
[5] Ahn HJ, Lee DS.Helicobacterpyloriin gastric carcinogenesis[J]. World J Gastrointest Oncol, 2015, 7(12): 455-465. DOI: 10.4251/wjgo.v7.i12.455
[6] Kim DH, Kim SW, Song YJ, et al. Long-term evaluation of mice model infected withHelicobacterpylori: focus on gastric pathology including gastric cancer[J]. Aliment Pharmacol Ther, 2003, 18(Suppl 1): 14-23. DOI: 10.1046/j.1365-2036.18.s1.4.x
[7] Mimura T, Yoshida M, Nishiumi S, et al. IFN-γ plays an essential role in the pathogenesis of gastric lymphoid follicles formation caused byHelicobactersuisinfection[J]. Fems Immunol Med Mic, 2011, 63(1): 25-34. DOI: 10.1111/j.1574-695x.2011.00823.x
[8] Liu J, He L, Haesebrouck F, et al. Prevalence of coinfection with gastric non-HelicobacterpyloriHelicobacter(NHPH) species inHelicobacterpylori-infected patients suffering from gastric disease in Beijing, China[J]. Helicobacter, 2015, 20(4): 284-290. DOI: 10.1111/hel.12201
[9] Haesebrouck F, Pasmans F, Flahou B, et al. Gastric helicobacters in domestic animals and nonhuman primates and their significance for human health[J].Clin Microbiol Rev, 2009, 22(2): 202-223. DOI:10.1128/cmr.00041-08
[10] van Loon S, Bart A, den Hertog EJ, et al.Helicobacterheilmanniigastritis caused by cat to child transmission[J]. J Pediatr Gastroenterol Nutr, 2003, 36(3): 407-409. DOI:10.1097/00005176-200303000-00021
[11] Svec A, Kordas P, Pavlis Z, et al. High prevalence ofHelicobacterheilmannii-associated gastritis in a small, predominantly rural area: further evidence in support of a zoonosis?[J]. Scand J Gastroenterol, 2000, 35(9): 925-928. DOI:10.1080/003655200750022968
[12] De Cooman L, Flahou B, Houf K, et al. Survival ofHelicobactersuisbacteria in retail pig meat[J]. Int J Food Microbiol, 2013, 166(1): 164-167. DOI:10.1016/j.ijfoodmicro.2013.05.020
[13] Groote DD, Ducatelle R, Van Doorn LJ, et al. Detection of “CandidatusHelicobactersuis” in gastric samples of pigs by PCR: comparison with other invasive diagnostic techniques[J]. J Clin Microbiol, 2000, 38(3): 1131-1135.
[14] Flahou B, Haesebrouck F, Pasmans F, et al.Helicobactersuiscauses severe gastric pathology in mouse and Mongolian gerbil models of human gastric disease[J]. PLoS ONE, 2010, 5(11): e14083-e14083. DOI: 10.1371/journal.pone.0014083
[15] Randall TD, Mebius RE. The development and function of mucosal lymphoid tissues: a balancing act with micro-organisms[J]. Mucosal Immunol, 2014, 7(3): 455-466. DOI: 10.1038/mi.2014.11
[16] Joosten M, Blaecher C, Flahou B, et al. Diversity in bacterium-host interactions within the speciesHelicobacterheilmanniisensu stricto[J]. Vet Res, 2013, 44(1): 65-75. DOI: 10.1186/1297-9716-44-65
Establishment of mice model withHelicobactersuisinfection and its significance
YANG Ruo-ming, YANG Lin, TIAN Zi-bin, YU Ya-nan,MAO Tao, LI Xiao-yu, ZHANG Shuai-qing
(DepartmentofGastroenterology,theAffiliatedHospitalofQingdaoUniversity,Qingdao266003,China)
The aim of this study was to establish mice model withHelicobactersuis(H.suis) infection and explore its significance. A total of 40 female C57BL/6 mice were randomly divided into model group and control group. The mice in model group were given 0.5 mL pig gastric mucosal homogenates containingH.suisby gavage, while the mice in control group were given the same amount of PBS. After 1 month and 3 months, half of each group were randomly sacrificed and their gastric mucosa were taken out for PCR and HE staining respectively to detect theH.suiscolonization and the formation of lymphoid follicles in the stomach. Results showed thatH.suisexisted in pig gastric mucosal homogenates used for gavage and could colonize in the stomach of mice, which also existed at 3 months postinfection. The formation of gastric lymphoid follicles was detected in the stomachs of mice at 1 month and 3 months afterH.suisformation, and the size of follicles at 3 months postinfection increased significantly compared with that at 1 month postinfection. In conclusion, a mouse model ofH.suisinfection was established successfully.H.suisinfection can induce the formation of lymphatic follicles in C57BL/6 mice. Therefore it may play an important role in the development of gastric mucosa-associated lymphoid tissue (MALT) Lymphoma.
H.suis; pig stomach; mice model; lymphoid follicles
Yang Lin, Email: bobotony@126.com
楊林,Email:bobotony@126.com
青島大學附屬醫(yī)院消化內(nèi)科,青島266003
R378
A
1002-2694(2016)07-0604-04
2016-02-24;
2016-06-19
DOI:10.3969/j.issn.1002-2694.2016.07.003
國家自然科學基金面上項目(No. 81572320)資助
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2020年10期)2020-11-26 08:24:50
學苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
數(shù)學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
海峽科技與產(chǎn)業(yè)(2016年3期)2016-05-17 04:32:12
