成年心肌梗死小鼠心肌細胞去分化現象觀察
2016-09-05 09:42:43沈文艷陳瑤李軍
山東醫藥 2016年18期
關鍵詞:小鼠
沈文艷,陳瑤,李軍
( 1上海交通大學醫學院附屬仁濟醫院,上海200217;2上海交通大學Med-X 臨床干細胞研究中心)
?
成年心肌梗死小鼠心肌細胞去分化現象觀察
沈文艷1,2,陳瑤2,李軍2
( 1上海交通大學醫學院附屬仁濟醫院,上海200217;2上海交通大學Med-X 臨床干細胞研究中心)
目的觀察成年心肌梗死小鼠心肌細胞是否存在去分化現象。方法20只成年小鼠隨機分為觀察組和對照組各10只,觀察組制備心肌梗死模型,對照組僅行假手術處理。采用RT-PCR法檢測兩組心肌組織 α-橫紋肌肌動蛋白(α-SA)、橫紋肌蛋白(α-actinin)、轉錄因子nkx2.5、肌細胞增強因子2(mef2)、細胞增殖核抗原(Ki67)、干細胞標記物血管內皮生長因子受體(flk-1) mRNA,采用免疫熒光染色法檢測兩組心肌組織磷酸化組蛋白(PH3)、連接蛋白43(cx43)。結果觀察組心肌組織α-SA、α-actinin、nkx2.5、mef2 mRNA表達量均低于對照組,Ki67、flk-1、mRNA表達量及PH3、cx43均高于對照組,P均<0.05, 結論成年心肌梗死小鼠心肌細胞存在去分化現象。
心肌梗死;心肌細胞;細胞去分化;α-橫紋肌肌動蛋白;橫紋肌蛋白;轉錄因子;細胞增殖核抗原;磷酸化組蛋白
急性心肌梗死(AMI)是冠狀動脈急性、持續性缺血缺氧所引起的心肌壞死,是引起充血性心力衰竭的重要原因[1]。心肌梗死發生后,梗死區域的心肌細胞因為缺血損傷而失去了功能,心臟的泵血功能嚴重受損[2, 3]。目前常用治療心肌梗死的方法一般是藥物和介入治療,這些治療方法對已梗死的心肌無法修復。傳統上認為,哺乳動物的心臟是一個終末分化器官[4],而心肌細胞是一種終末分化的細胞,心肌梗死后心肌細胞沒有再生能力而導致心力衰竭的發生[5]。然而,最近越來越多的研究顯示新生小鼠及新生兒心肌均具有再生能力[6~8]。也有研究顯示,成體動物的心肌細胞可能也具有類似的能力[9]。連接蛋白43(cx-43)、 α-橫紋肌肌動蛋白(α-SA)、橫紋肌蛋白(α-actinin)、轉錄因子nkx2.5、肌細胞增強因子2(mef2)是常用的心肌細胞標記物,細胞增殖核抗原(Ki67)和磷酸化組蛋白(PH3)是常用的細胞增殖標記物,血管內皮生長因子受體(flk-1)是常用的干細胞標記物。我們在前期研究中發現,心肌梗死可以誘導小鼠心肌細胞在體外發生類似去分化的現象,但是體內的研究目前尚未見報道。2013年5月~2014年3月,我們觀察了心肌梗死是否可誘導小鼠成體心肌細胞發生去分化現象。現報告如下。
[10] Bayat N, Ebrahimi-Barough S, Ardakan MM, et al. Differentiation of human endometrial stem cells into schwann cells in fibrin hydrogel as 3D culture[J]. Mol Neuro, 2015,6(4):1-7.
[11] Fan L, Yu Z, Li J, et al. Schwann-like cells seeded in acellular nerve grafts improve nerve regeneration[J]. BMC Mus Dis, 2014,15(6):165.
[12] Miura K, Okada Y, Aoi T, et al. Variation in the safety of induced pluripotent stem cell lines[J]. Nat Biotech, 2009,27(8):743-745.
[13] Xu H, Wang Y, He Z, et al. Direct conversion of mouse fibroblasts to GABAergic neurons with combined medium without the introduction of transcription factors or miRNAs[J]. Cell Cycle, 2015,14(15):2451-2460.
[14] Hoveizi E, Ebrahimi-Barough S, Tavakol S, et al. In Vitro differentiation of human iPS cells into neural iike cells on a biomimetic polyurea[J]. Mol Neuro, 2016,7(12):1-7.
[15] Liu Z, Tang Y, Lü S, et al. The tumourigenicity of iPS cells and their differentiated derivates[J]. J Cell Mol Med, 2013,17(6):782-791.
[16] Hu W, Qiu B, Guan W, et al. Direct conversion of normal and alzheimer′s disease human fibroblasts into neuronal cells by small molecules[J]. Cell stem cell, 2015,17(2):204-212.
[17] Li M, Zhang P, Li H, et al. TGF-β1 is critical for wallerian degeneration after rat sciatic nerve injury[J]. Neur Sci, 2015(284):759-767.
[18] Einheber S, Hannocks MJ, Metz CN,et al.Transforming growth factor-beta 1 regulates axon/Schwann cell interactions[J]. Jour Cell Bio, 1995,129(2):443-458.
[19] Leung AW, Kent Morest D, Li JY. Differential BMP signaling controls formation and differentiation of multipotent preplacodal ectoderm progenitors from human embryonic stem cells[J]. Develop Bio, 2013,379(2):208-220.
[20] Bonaguidi MA, Peng CY, McGuire T, et al. Noggin expands neural stem cells in the adult hippocampus[J]. J Neuro Sci, 2008,28(37):9194-9204.
[21] Monje PV. To myelinate or not to myelinate: fine tuning cAMP signaling in schwann cells to balance cell proliferation and differentiation[J]. Neu Regen Res, 2015,10(12):1936-1937.
[22] Latasa MJ, Cosgaya JM. Regulation of retinoid receptors by retinoic acid and axonal contact in schwann cells[J]. PLoS One,2011,6(2):17023.
[23] Zhang L, Ma Z, Smith GM, et al. GDNF-enhanced axonal regeneration and myelination following spinal cord injury is mediated by primary effects on neurons[J]. Glia, 2009,57(11):1178-1191.
1 材料與方法
1.1材料6~8周齡C57BL/6 SPF級雄性小鼠20只(體質量22~25 g),購自上海斯萊克模型動物有限公司;包埋劑OCT購自Sakura公司,1%SDS購自Sigma公司,Hanks平衡鹽溶液購自法國Biowest公司,脫氧核糖核酸酶Ⅰ(DNase Ⅰ)購自Roche公司;鼠源抗小鼠細胞Ki67抗體購自美國BD公司,兔源抗小鼠PH3抗體購自美國Cell Signaling 公司,鼠源抗小鼠α-SA抗體購自英國Abcam公司,兔抗小鼠cx43抗體購自美國CST公司;細胞核熒光染料 4,6-二脒基-2-苯基吲哚(DAPI)購自美國Sigma公司,驢源抗兔Alexa Fluor488熒光染料,驢源抗兔Alexa Fluor555熒光染料均購自美國Invitrogen公司。引物由上海生工生物合成有限公司合成。
1.2小鼠心肌梗死模型制備將20只小鼠隨機分為觀察組和對照組各10只。觀察組用異氟烷預麻箱麻醉后打開胸腔,于左心耳下緣約2 mm處結扎冠狀動脈左前降支,制備心肌梗死模型。以小鼠冠狀動脈左前降支供血區域心肌顏色變白[10]視為造模成功,最終造模成功10只。對照組僅麻醉后打開胸腔,不結扎血管行假手術處理。
1.3心肌組織α-SA、α-actinin、nkx2.5、mef2、Ki67、flk-1 mRNA表達檢測采用RT-PCR法。造模7 d取兩組小鼠,斷頭處死后取心肌組織;按照試劑盒說明書提取出總RNA,逆轉錄合成cDNA后進行擴增,以β-actin為內參。引物序列:β-actin上游引物5′-GGCTGTATTCCCCTCCATCG-3′,下游引物5′-CCAGTTGGTAACAATGCCATGT-3′,引物長度154 bp;Ki67上游引物5′-CCTGCCCGACCCTACAAAAT-3′,下游引物5′-TCCGCCGTCTTAAGGTAGGA-3′,引物長度313 bp;flk-1上游引物5′-AGAGTCAGACAACAACAATTGGCGAGAC-3',下游引物5′-GCATCATAAGGCAAGCGTTCACAGC-3′,引物長度451 bp;Nkx2.5上游引物5′-CAATGCCTATGGCTACAACGC-3′,下游引物5′-GAGTCATCGCCCTTCTCCTAAA-3′,引物長度297 bp;α-SA上游引物5′-CAGCCCTCTTTCATTGGT-3′,下游引物5′-CTGCCTCATCATACTCTT-3′,引物長度310 bp;mef上游引物5′-AAGTACACCGAGTACAACG-3′,下游引物5′-TAACTGGCATCTCAAAGC-3′,引物長度235 bp;α-actinin上游引物5′-GACCCAGATCATGTTTGAGACCTTC-3′,下游引物5′-CATTATGAGTTACACCATCGCCAGA-3′,引物長度130 bp。 反應體系 30 μL, 擴增條件:72 ℃、45 s延伸, 95 ℃ 、30 s,55 ℃、30 s,30個循環,72 ℃、10 min,4 ℃冷卻,進行瓊脂糖凝膠電泳。用Image J軟件分析目的條帶灰度,以其與β-actin的灰度比值計算目的基因相對表達量。實驗重復3次。
1.4心肌組織PH3、cx43表達檢測采用免疫熒光染色法。取造模7 d時兩組小鼠心肌組織, OCT包埋切片,驢血清封閉;用1%BSA稀釋抗體(1∶100)一抗孵育,4 ℃ 過夜。次日37 ℃復溫45 min,二抗孵育后洗滌。DAPI復染,封片,采用熒光顯微鏡觀察心肌組織PH3、cx43染色結果。用Image J軟件分別測免疫熒光結果中陽性熒光面積比上該視野的面積,計算相對表達量。實驗重復3次。
2 結果
2.1兩組心肌組織α-SA、α-actinin、nkx2.5、mef2、Ki67、flk-1 mRNA表達比較見表1。
2.2兩組心肌組織PH3、cx43表達比較見表2。
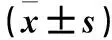
表1 造模7 d兩組心肌組織α-SA、α-actinin、nkx2.5、mef2、Ki67、flk-1 mRNA相對表達量比較
注:與對照組比較,#P<0.05。
3 討論
長期以來,人們認為心肌細胞是終末分化細胞,自身沒有再生能力。有研究顯示,心肌細胞可以被誘導再分裂,使心肌細胞去分化,重新進入細胞周期[11]。新生的心肌細胞也可以通過去分化的方式參與心臟自穩態的維護和病變組織的修復[12]。研究發現,成年小鼠心肌細胞可在急性損傷后發生去分化現象,以保證在存活基礎上在特定條件下進一步再生為心肌細胞。
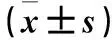
表2 造模7 d兩組心肌組織PH3、cx43相對表達量比較
注:與對照組比較,#P<0.05。
本研究發現,造模7 d觀察組心肌組織cx-43、α-SA及α-actinin mRNA相對表達量均低于對照組,提示心肌肌節解聚現象明顯,心肌細胞橫紋肌結構破壞。 Nkx2.5和mef2是兩個與心臟發育密切相關的轉錄因子。Nkx2.5參與了心臟形成,包括右側環化、心腔分化、心臟間隔功能成熟以及工作心肌和傳導系統的維持等過程[13]。心肌梗死后nkx2.5和mef2的表達減少,提示心肌梗死后成熟心肌細胞結構喪失,退回到未成熟心肌細胞或前體細胞狀態。我們前期研究顯示,心肌梗死后小鼠的心肌細胞有Ki67表達的增加,并與乳鼠心肌組織表達接近,進一步說明心肌梗死后部分心肌細胞的增殖能力可能退回到接近乳鼠未成熟心肌細胞或前體細胞的階段。在正常成體組織中,絕大部分細胞處于非增殖狀態,細胞學上稱為G0期。要構建良好的組織結構并執行特定的功能,必須依靠這些增殖周期外的G0期細胞。Ki67免疫組化染色可將大部分G0期以外的增殖細胞標記,因而也被稱為細胞的增殖指數。Ki67陽性率越高,說明處于增殖周期的細胞比例越高,組織分化越好。PH3只出現于有絲分裂期細胞染色體,靜止期細胞不表達,PH3表達的增高可以更準確地提示心肌梗死誘導成體心肌細胞增殖[14]。本研究發現,觀察組心肌組織存在Ki67和PH3高表達,與以往研究結果一致。flk-1是啟動胚胎造血發育的必需基因。Shalaby等[15]報道,flk-1基因能調節早期中胚層來源的間質細胞遷移至造血微環境;Miyagi等[16]發現小鼠胚胎干細胞體外發育至表達flk-1階段已具備重建造血功能,flk-1基因敲除的胚胎干細胞不能發育為造血細胞。本研究中,觀察組心肌組織中flk-1相對表達量明顯高于對照組,進一步提示心肌梗死后心肌細胞退回至未成熟心肌細胞或前體細胞階段。
綜上所述,成年心肌梗死小鼠心肌細胞存在去分化現象,但其具體作用機制尚有待于進一步研究。
[1] Guo J. Influence of dietary patterns on the risk of acute myocardial infarction in China population: the INTERHEART China study[J]. Chin Med J, 2013,126(3):464-470.
[2] Bearzi C. Human cardiac stem cells[J]. Proc Natl Acad Sci USA, 2007,104(35):14068-14073.
[3] Altarche W,Xifro F. Cardiac c-kit+AT2+cell population is increased in response to ischemic injury and supports cardiomyocyte performance[J]. Stem Cells, 2009,27(10): 2488-2497.
[4] Hosoda T. C-kit-positive cardiac stem cells and myocardial regeneration[J]. Am J Card Dis, 2012, 2(1): 58-67.
[5] Zaruba MM. Cardiomyogenic potential of C-kit(+)-expressing cells derived from neonatal and adult mouse hearts[J]. Circulation, 2010,121(18): 1992-2000.
[6] Porrello ER, Mahmoun Al, Simpson E, et al. Transient regenerative potential of the neonatal mouse heart[J]. Science, 2011,331(6020):1078-1080.
[7] Rücker MC, Pecker F, Godreau D, et al. Dedifferentiation of atrial myocytes during atrial fibrillation:role of fibroblast proliferation in vitro[J]. Cardiovasc Res, 2002,55(1):38-52.
[8] Polizzotti BD, Ganapathy B, Walsh S, et al. Neuregulin stimulation of cardiomyocyte regeneration in mice and human myocardium reveals a therapeutic window[J]. Sci Transl Med, 2015,7(281):231-245.
[9] Zhang Y, Li TS, Lee ST, et al. Dedifferentiation and proliferation of mammalian cardiomyocytes[J]. PLoS One, 2010,5(9):12559.
[10] Breitbach M, Bostani T, Roell W, et al. Potential risks of bone marrow cell transplantation into infarcted hearts [J]. Blood,2007,110(4):1362-1369.
[11] Engel FB, Schebest M, Duong MT, et al. P38 MAP kinase inhibition enables proliferation of adult mammalian cardiomyocytes[J]. Genes Dev, 2005,19(10):1175-1187.
[12] Beltrami AP,Urdanek K,Kajstura J,et al.Evidence that human cardiac myocytes divide after myocarction[J]. N Engl J Med,2001(344):1750-1757.
[13] Matsuura K. Cardiomyocytes fuse with surrounding noncardiomyocytes and reenter the cell cycle[J]. J Cell Biol, 2004,167(2):351-363.
[14] Engel FB,Schebest M,Duong MT,et al. P38 MAP kinase inhibition enables proliferation of adult mammalian cardiomyocytes[J]. Genes Dev, 2005,19(10):1175-1187.
[15] Shalaby F, Ho J, Stanford WL, et al. A requirement for Flk-1 in primitive and definitive hematopoiesis and vasculogenesis[J]. Cell, 1997,89(6);981-990.
[16] Miyagi T, Takeno M, Nagafuchi H, et al. FLK1+cells derived from mouse emhryonic stem cells recruiting VEGFR1+stem cells from bone-marrow microenvironment[J]. Nature Med, 2002,18(8):841-849.
(收稿日期:2016-01-15)
國家自然科學基金資助項目(81300088)。
李軍(E-mail: jun.li@sjtu.edu.cn)
10.3969/j.issn.1002-266X.2016.18.011
R544.22
A
1002-266X(2016)18-0034-03
2016-01-20)
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
