大豆籽粒大小與形狀性狀的QTL定位
2016-09-21 01:13:43強閆龍鄧瑩瑩肖二寧劉兵強楊春燕張孟臣
作物學報 2016年9期
陳 強閆 龍鄧瑩瑩 肖二寧 劉兵強 楊春燕張孟臣
河北省農(nóng)林科學院糧油作物研究所 / 國家大豆改良中心石家莊分中心 / 農(nóng)業(yè)部黃淮海大豆生物學與遺傳育種重點實驗室 / 河北省遺傳育種重點實驗室, 河北石家莊 050035
大豆籽粒大小與形狀性狀的QTL定位
陳 強**閆 龍**鄧瑩瑩 肖二寧 劉兵強 楊春燕*張孟臣*
河北省農(nóng)林科學院糧油作物研究所 / 國家大豆改良中心石家莊分中心 / 農(nóng)業(yè)部黃淮海大豆生物學與遺傳育種重點實驗室 / 河北省遺傳育種重點實驗室, 河北石家莊 050035
大豆籽粒大小和粒形性狀不僅與產(chǎn)量和外觀品質(zhì)緊密相關, 還對機械化播種有著一定的影響。本研究采用大粒栽培品種冀豆12與小粒半野生地方品種黑豆(ZDD03651)雜交衍生的包含188個重組自交系的F6:8和F6:9群體為材料, 對粒長、粒寬、粒厚、長寬比、長厚比和寬厚比的遺傳結(jié)構(gòu)進行分析, 并分別以WinQTLCart 2.5、QTLNetwork 2.1和IciMapping 4.1 3種模型對以上性狀的加性效應QTL, QE互作效應及上位性互作效應進行檢測。6個性狀的廣義遺傳率介于64.01%~79.57%, 遺傳力較高, 且除粒厚外的其他性狀受環(huán)境影響顯著。共定位到加性效應QTL 38個,單個QTL的貢獻率介于2.21%~10.71%之間, 分布在12條染色體的17個標記區(qū)間內(nèi), 且12個染色體區(qū)段至少與2種性狀相關。兩種及以上模型同時檢測到的QTL有24個, 3種模型均能檢測到的QTL共8個, 分別為qSL-17-1、qSL-18-1、qSW-6-1、qST-2-1、qST-6-1、qSLT-2-2、qSWT-2-1和qSWT-20-1。檢測到7對上位性互作QTL, 分別涉及粒長、粒寬、長寬比、長厚比和寬厚比, 互作效應貢獻率介于0.78%~6.20%之間。QE互作效應貢獻率均較低, 介于0.0005%~0.3900%之間。以多種模型同時檢測結(jié)果準確性較高, 可為分子標記輔助育種工作提供可靠理論基礎。
大豆; 粒形; QTL; 上位性互作
在大豆中, 粒長、粒寬、粒厚等籽粒大小性狀是衡量百粒重的重要指標, 與產(chǎn)量密切相關[1]。而大豆粒形性狀直接關系大豆的品質(zhì)和商品性, 既是機械化播種的重要限制因素, 又在一定程度上決定大豆在國際貿(mào)易中的商業(yè)價值[2]。籽粒大小和粒形性狀為多基因控制的數(shù)量性狀[3], 籽粒大小性狀遺傳力一般為 59%~79%, 粒形性狀的遺傳力一般在19%~56%間[4]。通過QTL定位分析, 檢測籽粒大小和粒形相關 QTL位點, 對分子標記輔助選擇的高產(chǎn)、高品質(zhì)大豆育種具有重要的指導意義。
在水稻中籽粒大小和粒形的研究較為深入[5-9],定位了較多 QTL, 并克隆了控制粒長和粒重的基因GS3[5], 控制粒寬和粒重的基因 GW2[6], 控制粒寬、長寬比和粒重的基因qSW5[7]等多個基因。大豆籽粒大小和粒形相關性狀QTL定位的研究相對較少[10-13]。大豆數(shù)據(jù)網(wǎng)站 SoyBase (http://www.soybase.org/)中公布了粒長QTL 18個、粒寬QTL 16個、粒厚QTL14個。近期, 牛遠等[14-15]在初定位基礎上, 采用Lasso法和復合區(qū)間作圖法, 同時將 O連鎖群中 Satt331-Satt592區(qū)段內(nèi)多效性QTL分解后精細定位, 并對目標區(qū)段內(nèi)候選基因進行預測。Xie等[16]對位于C2連鎖群Satt640-Satt422區(qū)間內(nèi)的粒形QTL精細定位分析, 分別檢測到粒長、粒寬、粒厚及百粒重相關聯(lián)的 SNP標記, 并預測了結(jié)果區(qū)段內(nèi)可能的效應基因。Hu等[17]在分離群體中檢測到12個籽粒大小和粒形相關QTL, 在關聯(lián)分析中檢測到41個SNP標記與6個性狀相關聯(lián)。而隨著研究的深入, QTL間的上位性互作及 QE互作逐漸受到研究人員的重視[18-20]。Niu等[21]以 257份大豆栽培品種為材料, 采用關聯(lián)分析方法檢測到59個主效QTL和31個QTL與環(huán)境互作效應。梁慧珍等[22]利用QTLNetwork 2.0軟件檢測到3對粒形性狀上位性互作QTL。閆寧等[23]采用 Bayes分層廣義線性模型對粒形性狀的主效QTL、QTL與環(huán)境互作效應、QQ上位性互作及QTL與細胞質(zhì)互作效應分別進行檢測。蘇成付等[24]和李杰勤等[25]研究結(jié)果顯示不同的定位程序適用的遺傳模型范圍不同, 建議使用多種作圖法共同作圖, 并優(yōu)先標定共同發(fā)現(xiàn)的QTL。
目前, 大豆粒形相關性狀上位性互作及 QE互作研究相對較少, 且前人研究多采用單一模型檢測QTL。本研究利用重組自交系群體為材料, 分別采用WinQTLCart 2.5[26]、QTLNetwork 2.1[27]和IciMapping 4.1[28-29]軟件對籽粒大小和粒形各性狀的加性 QTL,QTL間上位性互作及 QE互作進行檢測, 以得到可靠的QTL信息, 為進一步精細定位及輔助育種提供依據(jù)。
1 材料與方法
1.1材料及田間試驗
以冀豆 12×黑豆雜交構(gòu)建的包含 188個家系的F6:8和 F6:9重組自交系為材料。冀豆 12為河北省農(nóng)林科學院糧油作物研究所育成的百粒重為22 g的大粒品種, 黑豆(ZDD03651)為陜西省半野生地方品種,百粒重為7 g。兩親本粒長、粒寬等籽粒大小相關性狀間存在較大差異。將該群體 F6:8(2011年)和 F6:9(2013年)重組自交系播種于石家莊田間, 采用 3 m行長, 3行區(qū), 隨機區(qū)組試驗設計, 行距50 cm, 株距10 cm。材料自然成熟后從每小區(qū)收取中間行中間10株。
1.2性狀調(diào)查
自每家系籽粒中隨機選取10粒, 采用數(shù)顯游標卡尺測量粒長(seed length, SL)、粒寬(seed width,SW)、粒厚(seed thickness, ST), 長寬比(seed lengthto-width, SLW) = 粒長/粒寬、長厚比(seed length-tothickness, SLT) = 粒長/粒厚、寬厚比(seed width-tothickness, SWT) = 粒寬/粒厚[21], 重復2次, 以平均數(shù)為觀測值。
1.3統(tǒng)計分析
采用 SPSS 17.0軟件中相應程序?qū)Ω餍誀钸M行方差、相關性和偏相關等分析。
1.4QTL定位
在陳強[32]和雷雅坤等[33]分子遺傳連鎖圖譜的基礎上, 采用WinQTLCart 2.5[34]中的復合區(qū)間作圖法(CIM)檢測每個性狀主效QTL位點, 選擇模型6, 向前回歸方法, 選擇被檢區(qū)間每側(cè)10 cM的控制窗口,進行 1000次排列獲得 QTL顯著的 LOD閾值(P<0.05)。利用QTLNetwork 2.1軟件中基于混合線性模型的復合區(qū)間作圖法(MCIM)分析QTL 之間的互作效應[35-36], 將 Permutation次數(shù)設置為 1000次,用于計算F值。以1 cM的步長對全基因組進行QTL掃描, 選擇結(jié)果中 P值顯著的互作確定上位性互作及環(huán)境互作QTL。以IciMapping 4.1軟件利用MET模塊中ICIM-ADD和ICIM-EPI進行多環(huán)境聯(lián)合分析, 以ICIM法進行上位性QTL定位, 常用LOD值為2.0, 由于本研究包含2個環(huán)境, 再結(jié)合定位結(jié)果,故選取LOD值為4.0, 超過閾值就認為存在QTL[37]。同一性狀的LOD峰值相距在5 cM以內(nèi)者視為同一QTL。采取McCouch等[38]的方式命名QTL。
2 結(jié)果與分析
2.1表型變異及相關分析
兩年的表型數(shù)據(jù)分析結(jié)果(表1)顯示, 冀豆12的粒長、粒寬和粒厚高于黑豆, 而黑豆的長寬比、長厚比和寬厚比高于冀豆12。群體中粒形相關6個性狀均存在較大程度分離, 變異系數(shù)介于 5.30%~8.71%之間, 且各性狀均存在超雙親分離現(xiàn)象。各性狀遺傳力介于 64.01%~79.57%之間。基因型與環(huán)境間的雙因素方差分析結(jié)果顯示, 粒長、粒寬和寬厚比受基因型、環(huán)境和基因型×環(huán)境的影響均極顯著(P<0.01); 粒寬受基因型的影響極顯著(P<0.01); 長寬比受基因型和環(huán)境的影響均極顯著(P<0.01); 長厚比受基因型和環(huán)境的影響極顯著(P<0.01), 而基因與環(huán)境互作的影響顯著(P<0.05)。各性狀的表型值均連續(xù)變異, 且為正態(tài)分布(圖1)。
偏相關分析結(jié)果顯示(表2), 粒長與粒寬、長寬比間均顯著正相關, 與粒厚極顯著負相關, 與長厚比顯著負相關, 與寬厚比在2011年相關性不顯著,2013年極顯著負相關。粒寬與粒厚、寬厚比極顯著正相關, 與長寬比極顯著負相關; 與長厚比在2011年相關性不顯著, 2013年極顯著負相關。粒厚與長寬比極顯著正相關, 與寬厚比極顯著負相關, 與長寬比相關性不顯著。長寬比與長厚比極顯著正相關,與寬厚比極顯著負相關。長寬比與長厚比間為極顯著正相關。粒長與寬厚比2011年簡單相關分析為極顯著正相關, 偏相關分析無顯著相關性, 2013年簡單相關無顯著相關性, 偏相關分析為極顯著負相關。性狀間相關性不穩(wěn)定, 表明該相關受其他性狀的影響較大。
2.2主效QTL定位及QE互作效應分析
WinQTLCart 2.5和QTLNetwork 2.1進行1000次排列獲得QTL顯著的LOD閾值均為2.5。兩年中6性狀共檢測到38個主效應QTL, 分布在12條染色體中(圖2), 單個QTL的貢獻率介于2.21%~10.71%之間。24個QTL能在2種及以上的方法中檢測到,其中8個QTL可被3種方法同時檢測到(表3)。
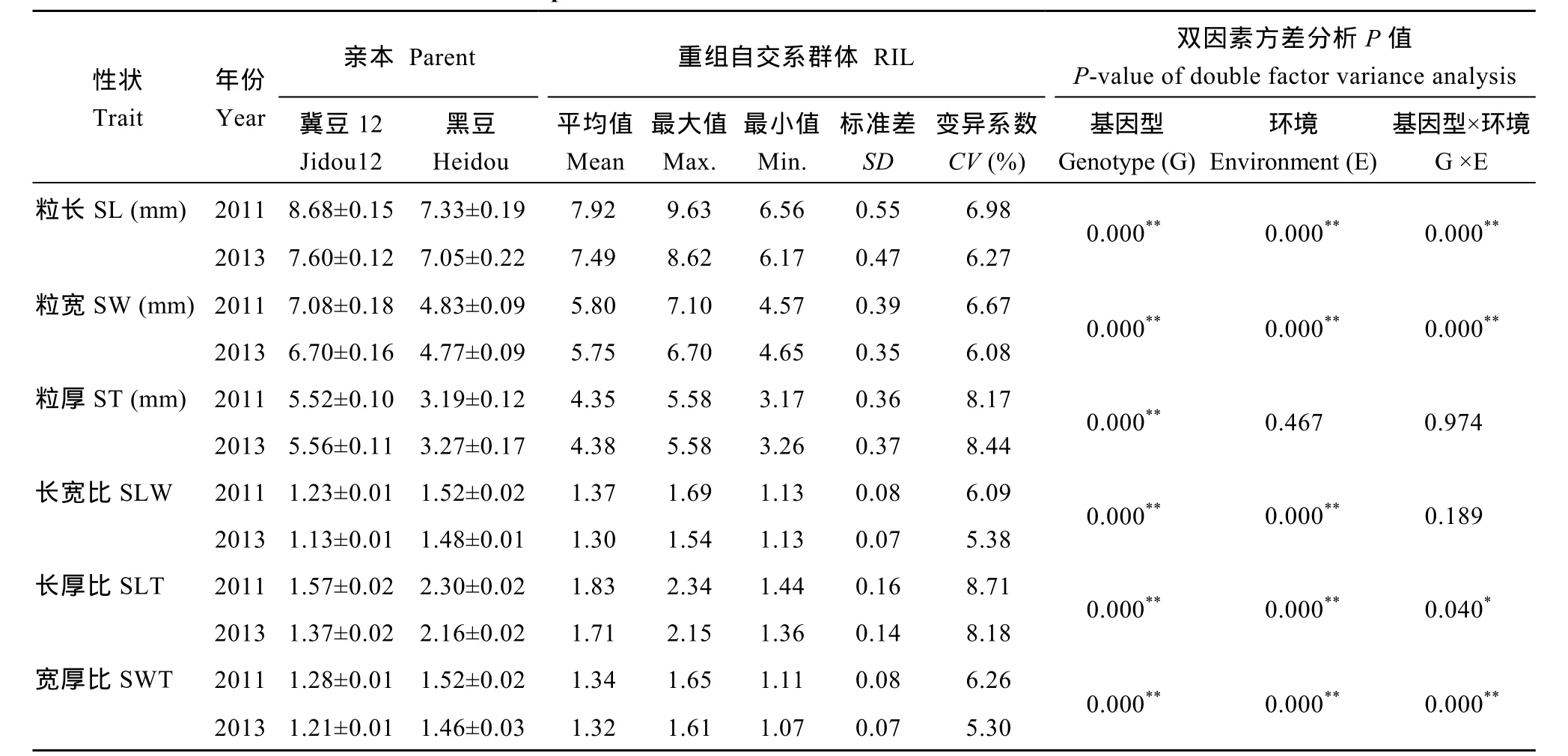
表1 親本及群體籽粒大小與粒形性狀在2011和2013年中的表現(xiàn)Table 1 Characteristics of seed size and shape traits in RILs derived from the cross between Jidou 12 and Heidou in 2011 and 2013
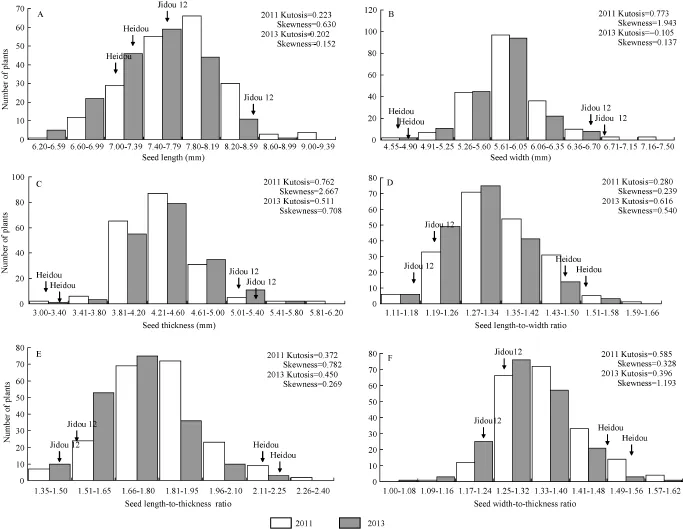
圖1 群體粒形相關性狀頻率分布圖Fig. 1 Frequency distribution of seed size traits of soybean populations in 2011 and 2013A: 群體粒長表型分布; B: 群體粒寬表型分布; C: 群體粒厚表型分布; D: 群體長寬比表型分布; E: 群體長厚比表型分布; F: 群體寬厚比表型分布。A: frequency distribution of seed length; B: frequency distribution of seed width; C: frequency distribution of seed thickness; D: frequency distribution of seed length-to-width ratio; E: frequency distribution of seed length-to-thickness ratio; F: frequency distribution of seed width-to-thickness ratio.
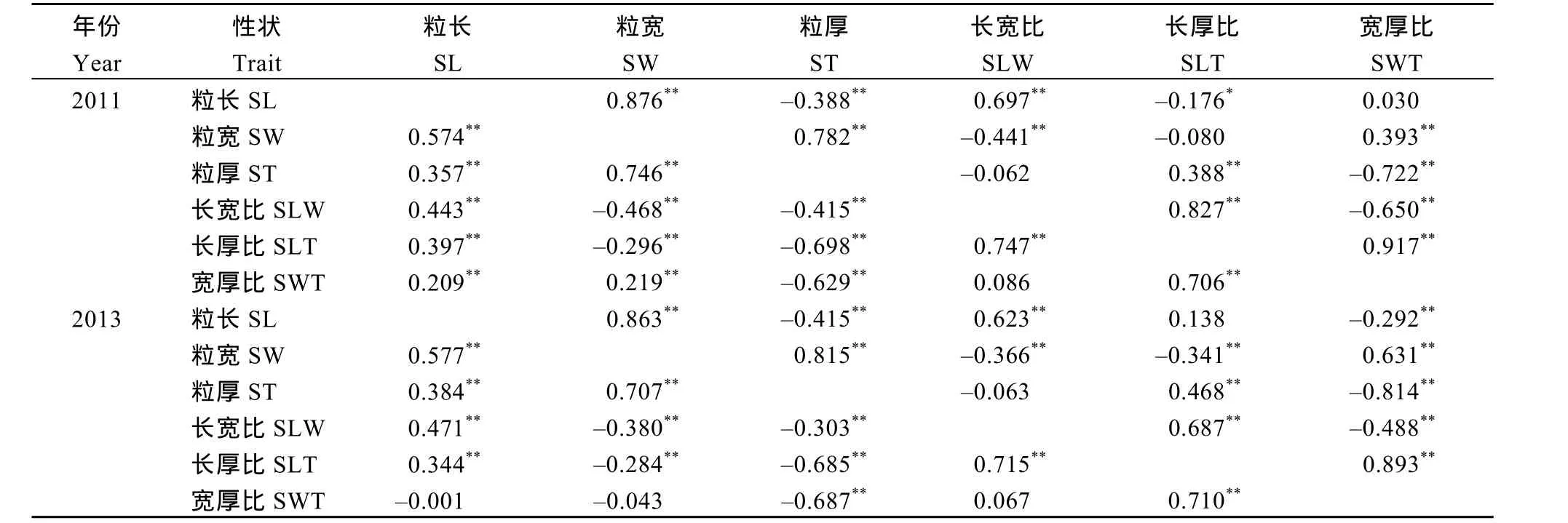
表2 兩年中粒形性狀間的簡單相關和偏相關分析Table 2 Simple and partial correlation coefficients for seed traits in soybean in two years
檢測到4個控制粒長的加性QTL, 貢獻率范圍為4.04%~8.76%。其中qSL-17-1和qSL-18-1可被3種方法同時檢測到, 貢獻率最大分別為 4.67%和8.72%, 加性效應最大分別為-0.10 mm和0.15 mm,增效基因分別來自于冀豆12和黑豆, 與環(huán)境互作的貢獻率最大分別為0.10%和0.29%。檢測到3個控制粒寬的加性QTL, 其中qSW-6-1在3種方法中均能被檢測到, 貢獻率最大為 9.59%, 加性效應最大值為-0.12 mm, 與環(huán)境互作的貢獻率最大值為0.02%;qSW-17-1和qSW-2-1分別被兩種方法檢測到, 貢獻率最大分別為 6.37%和 7.44%, 加性效應值為-0.12 mm 和-0.15 mm, 環(huán)境互作貢獻率分別為0.03%和0.04%。檢測到7個控制粒厚的加性QTL,其中qST-6-1和qST-2-1在3種方法中可同時被檢測到, 貢獻率最大分別為6.80%和5.72%, 加性效應值最大分別為-0.10 mm和-0.09 mm, 環(huán)境互作貢獻率最大值分別為0.20%和0.09%; qST-8-1、qST-12-1、qST-13-1和qST-17-1同時被2種方法檢測到, 貢獻率最大分別為5.61%、8.01%、6.73%和6.25%, 加性效應最大分別為-0.07、-0.09、-0.10和-0.10 mm, 與環(huán)境互作貢獻率范圍為0.01%~0.27%。
檢測到 3個控制長寬比的加性 QTL, 其中qSLW-2-1在兩種方法中同時被檢測到, 貢獻率最大為8.67%, 加性效應最大分別為0.07, 與環(huán)境互作貢獻率為0.39%。共檢測到9個控制長厚比的加性QTL,其中qSLT-2-2在3種方法中同時被檢測到, 貢獻率最大為8.69%, 加性效應最大為0.07, 與環(huán)境互作貢獻率為0.15%。qSLT-8-1和qSLT-13-1分別在兩種方法中檢測到, 貢獻率最大分別為5.07%和8.23%。共檢測到 8個控制寬厚比的加性 QTL, qSWT-2-1和qSWT-20-1可同時在 3種方法中被檢測到, 貢獻率最大分別為7.87%和7.17%, 加性效應均為0.02, 與環(huán)境互作貢獻率分別為0.29%和0.23%; qSWT-6-1、qSWT- 8-1、qSWT-13-1和qSWT-17-1可分別被兩種方法檢測到, 貢獻率分別為7.05%、10.71%、8.51% 和6.46%。
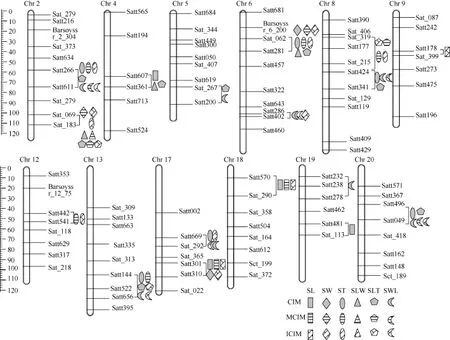
圖2 檢測到的主效QTL在連鎖群中的分布Fig. 2 Distribution of main-effect QTLs for seed shape traits on linkage groups

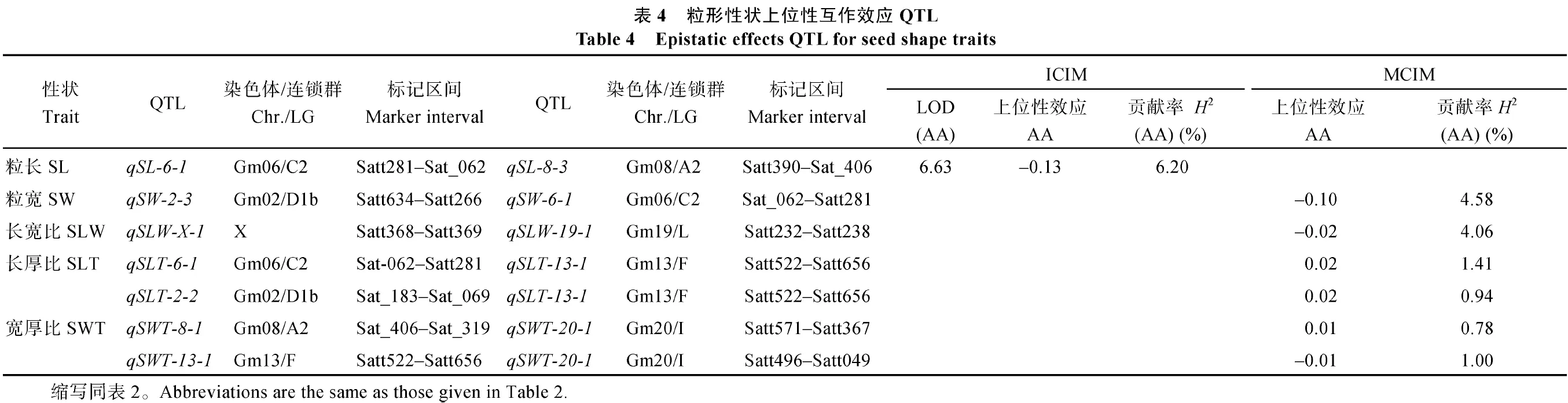
2.3粒形性狀上位性互作效應結(jié)果
共檢測到 7對上位性互作 QTL, 分別涉及粒長、粒寬、長寬比、長厚比和寬厚比(表 4)。粒長上位性互作 QTL發(fā)生在非主效 QTL間, 貢獻率為6.20%; 粒寬和長寬比上位性互作均發(fā)生在主效QTL與非主效 QTL間, 貢獻率分別為 4.58%和4.06%, 效應值分別為-0.10 mm和-0.02, 表明該互作為親本型大于重組型; 長厚比中檢測到 2對發(fā)生在主效QTL間的上位性互作, 為qSLT-13-1分別與qSLT-6-1和 qSLT-2-2發(fā)生的互作, 貢獻率分別為1.41%和 0.94%, 效應值均為 0.02。寬厚比上位性QTL為qSWT-20-1分別與qSWT-8-1和qSWT-13-3互作, 效應值均為0.01, 但效應相反。
3 討論
3.1與前人研究結(jié)果的比較
在QTL定位研究中, 前人建議使用多種作圖法,并優(yōu)先標定共同發(fā)現(xiàn)的QTL[24-25]。本研究采用的3種作圖方法, 共檢測到籽粒大小和粒形相關性狀的加性效應 QTL 38個, 其中 3種方法同時檢測到的QTL有8個, 分別是粒長QTL qSL-17-1和 qSL-18-1,粒寬QTL qSW-6-1, 粒厚QTL qST-2-1和qST-6-1,長厚比 QTL qSLT-2-2, 寬厚比 QTL qSWT-2-1和qSWT-20-1。參照Song等[39]的大豆公共圖譜標記信息與前人研究結(jié)果比較發(fā)現(xiàn), 以上 8個位點中有 5個與前人報道基本一致。本研究檢測到的位于第18染色體的粒長 QTL, 標記區(qū)間為 Satt570-Sat_290, 與Moongkanna等[10]檢測到粒長QTL區(qū)間Satt303-Sat_ 315部分重疊; 位于第17染色體的粒長QTL標記區(qū)間為 Satt301-Satt310, 與 Yu等[18]檢測到粒長 QTL區(qū)間Satt266-Sat_354部分重疊。本研究檢測到的位于第 6染色體的粒寬、粒厚 QTL標記區(qū)間為Sat_062-Satt28, 與 Yu等[18]檢測到粒寬 QTL區(qū)間Satt640-Satt422重疊, 且 Xie等[16]在該區(qū)段內(nèi)對粒長、粒寬和粒厚精細定位分析, 并分別檢測到關聯(lián)標記, Salas等[11]在該區(qū)段內(nèi)同樣檢測到粒寬和粒厚QTL。本研究檢測到的位于第 20染色體的寬厚比QTL標記區(qū)間為Satt496-Satt049, 與Yu等[18]檢測到粒寬QTL區(qū)間Satt419-Satt354部分重疊。該結(jié)果表明多種方法同時檢測到的位點有著較高的準確性。而對于粒厚QTL qST-2-1、長厚比QTL qSLW-2-2、寬厚比QTL qSWT-2-1則未見報道。
本研究用2種方法檢測到的QTL有14個, 僅用1種方法檢測到的QTL有14個。其中在第4染色體中的粒長QTL qSL-4-1標記區(qū)間為Satt607-Satt361,Niu等[21]檢測到的粒長相關聯(lián)的標記AW277661位于該區(qū)段內(nèi); 在第8染色體中粒厚QTLqST-8-2標記區(qū)間Satt424-Satt341內(nèi)同樣含有Niu等[21]檢測到的粒厚相關聯(lián)的標記 AW132402; 在第 5染色體長厚比和寬厚比的QTLqSLT-5-1和qSWT-5-1標記區(qū)間Sat_267-Satt200, 與 Salas等[11]檢測到的粒形 QTL標記區(qū)間A975_1~K636_2部分重疊。
本研究中檢測到的位于在第 2染色體 Satt266-satt611區(qū)間、第2染色體Sat_069-Sat_183區(qū)間、第12染色體 Satt422-Satt541區(qū)間、第 13染色體Satt144-Satt656區(qū)間和第17染色體Satt669-Sat_292區(qū)間, 共檢測到13個QTL, 均未見前人報道。
3.2染色體區(qū)段多效性分析
本研究檢測到的38個QTL集中分布在17個標記區(qū)間內(nèi), 其中13個標記區(qū)段存在同時與多個性狀相關的現(xiàn)象。前人研究表明, 作物相關性狀QTL的集中分布是一種普遍現(xiàn)象, 有不少Q(mào)TL可能具有多效性。Mansur等[40]認為與性狀間的高度遺傳力和遺傳相關一致, 可能是一個上位基因作用的結(jié)果, 也可能是控制這些性狀的基因緊密排列所致。在水稻中, 杜景紅等[41]應用剩余雜合體衍生群體, 在初定位基礎上, 把水稻第 6染色體短臂上控制水稻谷殼硅含量的qHUS-6分解為qHUS6-1和qHUS6-2, 并進一步將 qHUS6-2分解為一個效應較大的qHUS6-2a和效應較小的 qHUS6-2b。在大豆中, 牛遠等[14-15]通過構(gòu)建剩余雜合系和重組自交系, 將 O染色體Satt331-Satt592 區(qū)間剖分為與標記S21-S22 和O23~O19/O19~O21關聯(lián)的2個多效性QTL。Xie等[16]對第6染色體Satt640-Satt422間的粒形性狀效應分析, 分別檢測到粒長、粒寬和粒厚相關聯(lián)的SNP標記。梁慧珍等[22]對粒形性狀進行 QTL定位分析,同樣發(fā)現(xiàn)同一區(qū)段同時與多個性狀相關的現(xiàn)象, 通過對不同性狀間增效基因的來源對區(qū)段的多效性進行了初步推測。
本研究第2染色體Sat_069-Sat_183區(qū)段和第6染色體Sat_062-Satt281區(qū)段均同時與粒寬、粒厚、長寬比和長厚比相關, 第 4染色體 Satt607-Satt361區(qū)段同時與粒長、長寬比和長厚比相關, 第13染色體Satt144-Satt656區(qū)段同時與粒厚、長厚比和寬厚比相關。以上4個區(qū)段內(nèi)的QTL, 對于籽粒大小性狀(粒長、粒寬或粒厚)增效基因來自冀豆12, 而對于粒形性狀(長寬比、長厚比或?qū)捄癖龋﹦t相反, 推測其可能為多個 QTL的聚合。位于第 17染色體Satt301-Satt310區(qū)段同時與粒長和粒寬相關, 且增效基因均來自冀豆12, 推測其可能為多效性QTL。具體是一個主效基因還是由多個 QTL 緊密連鎖的聚合體還需進一步研究。
4 結(jié)論
共檢測到籽粒大小和粒形相關性狀中加性效應QTL 38個, 以3種方法同時檢測到的位點8個, 分別為qSL-17-1、qSL-18-1、qSW-6-1、qST-2-1、qST-6-1、qSLT-2-2、qSWT-2-1和qSWT-20-1, 可靠性較高, 其中qST-2-1、qSLT-2-2和qSWT-2-1為首次報道。環(huán)境互作效應貢獻率均較低, 而上位性互作位點 7對,效應相對較高, 其中對粒形性狀影響顯著, 不能忽略。
References
[1] Liang H Z, Li W D, Wang H, Fang X J. Genetic effects on seed traits in soybean. Acta Genet Sin, 2005, 32: 1199-1204
[2] Wilson D O Jr. Storage of orthodox seeds. In: Basra A S ed. Seed Quality: Basic Mechanisms Agricultural Implications. New York:Food Products Press, 1995. pp 173-208
[3] Nelson R L, Wang P. Variation and evaluation of seed shape in soybean. Crop Sci, 1989, 29: 147-150
[4] Cober E R, Voldeng H D, Fregeau-Reid J A. Heritability of seed shape and seed size in soybean. Crop Sci, 1997, 37: 1767-1769
[5] Fan C C, Xing Y Z, Mao H L, Lu T T, Han B, Xu C G, Li X H,Zhang Q F. GS3, a major QTL for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein. Theor Appl Genet, 2006, 112:1164-1171
[6] Song X J, Huang W, Shi M, Zhu M Z, Lin H X. A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase. Nat Genet, 2007, 39: 623-630
[7] Wan X Y, Weng J F, Zhai H Q, Wang J K, Lei C L, Liu X L, Guo T, Jiang L, Su N, Wan J M. QTL analysis for rice grain width and fine mapping of an identified QTL allele gw-5 in a recombination hotspot region on chromosome 5. Genetics, 2008, 179: 2239-2252
[8] Zhang X J, Wang J F, Huang J, Lan H X, Wang C L, Yin C F, Wu Y Y, Tang H J, Qian Q, Li J Y, Zhang H S. Rare allele of OsPPKL1 associated with grain length causes extra-large grain and a significant yield increase in rice. Proc Natl Acad Sci USA,2012, 109: 21534-21539
[9] Sun L J, Li X J, Fu Y C, Zhu Z F, Tan L B, Liu F X, Sun X Y, Sun X W, Sun C Q. GS6, a member of the GRAS gene family, negatively regulates grain size in rice. J Integr Plant Biol, 2013, 55:938-949
[10] Moongkanna J, Nakasathien S, Novitzky W P, Kwanyuen P, Sinchaisri P, Srinives P. SSR markers linking to seed traits and total oil content in soybean. Thai J Agric Sci, 2011, 44: 233-241
[11] Salas P, Oyarzo-Llaipen J C, Wang D, Chase K, Mansur L. Genetic mapping of seed shape in three populations of recombinant inbred lines of soybean (Glycine max L. Merr.). Theor Appl Genet, 2006, 113: 1459-1466
[12] 劉曉芬. 大豆栽培品種群體粒形性狀及百粒重的關聯(lián)分析.南京農(nóng)業(yè)大學博士學位論文, 江蘇南京, 2012 Liu X F. Association Analysis for Seed Shape Traits and 100-seed Weight in Soybean (Glycine max L. Merr.). PhD Dissertation of Nanjing Agricultural University, Nanjing, China, 2012 (in Chinese with English abstract)
[13] Xu Y, Li H N, Li G J, Wang X, Cheng L G, Zhang Y M. Mapping quantitative trait loci for seed size traits in soybean (Glycine max L. Merr.). Theor Appl Genet, 2011, 122: 581-594
[14] 牛遠, 徐宇, 李廣軍, 王云清, 劉曉芬, 李河南, 魏世平, 章元明. 大豆籽粒大小和粒形的馴化研究. 大豆科學, 2012, 31:522-528 Niu Y, Xu Y, Li G J, Wang Y Q, Liu X F, Li H N, Wei S P, Zhang Y M. Domestication of seed size and shape traits in soybean. Soybean Sci, 2012, 31: 522-528 (in Chinese with English abstract)
[15] 牛遠, 謝芳騰, 布素紅, 謝尚潛, 韓世鳳, 耿青春, 劉兵, 章元明. 大豆粒形性狀 QTL的精細定位. 作物學報, 2013, 39:609-616 Niu Y, Xie F T, Bu S H, Xie S Q, Han S F, Geng Q C, Liu B,Zhang Y M. Fine mapping of quantitative trait loci for seed shape traits in soybean. Acta Agron Sin, 2013, 39: 609-616 (in Chinese with English abstract)
[16] Xie F T, Niu Y, Zhang J, Bu S H, Zhang H Z, Geng Q C, Feng J Y,Zhang Y M. Fine mapping of quantitative trait loci for seed size traits in soybean. Mol Breed, 2014, 34: 2165-2178
[17] Hu Z B, Zhang H R, Kan G Z, Ma D Y, Zhang D, Shi G X, Hong D L, Zhang G Z, Yu D Y. Determination of the genetic architecture of seed size and shape via linkage and association analysis in soybean (Glycine max L. Merr.). Genetica, 2013, 141: 247-254
[18] Yu S B, Li J X, Xu C G. Importance of epistasis as the genetic basis of heterosis in an elite rice hybrid. Proc Natl Acad Sci USA,1997, 94: 922-931
[19] 孫亞男, 仕相林, 蔣洪蔚, 孫殿軍, 辛大偉, 劉春燕, 胡國華,陳慶山. 大豆百粒重 QTL的上位效應和基因型×環(huán)境互作效應分析. 中國油料作物學報, 2013, 34: 598-603 Sun Y N, Shi X L, Jiang H W, Sun D J, Xin D W, Liu C Y, Hu G H, Chen Q S. Epistatic effects and QE interaction effects of QTLs for 100-seed weight in soybean. Chin J Oil Crop Sci, 2013, 34:598-603 (in Chinese with English abstract)
[20] Xing Y Z, Tan Y F, Hua J P, Sun X L, Xu C G, Zhang Q F. Characterization of the main effects, epistatic effects and their environmental interactions of QTLs on the genetic basis of yield traits in rice. Theor Appl Genet, 2002, 105: 248-257
[21] Niu Y, Xu Y, Liu X F, Yang S X, Wei S P, Xie F T, Zhang Y M. Association mapping for seed size and shape traits in soybean cultivars. Mol Breed, 2013, 31: 785-794
[22] 梁慧珍, 余永亮, 楊紅旗, 張海洋, 董薇, 杜華, 暐崔文, 劉學義, 方宣鈞. 大豆粒形性狀主效QTL環(huán)境互作和上位性檢測.中國農(nóng)業(yè)科學, 2013, 46: 5081-5088 Liang H Z, Yu Y L, Yang H Q, Zhang H Y, Dong W, Du H, Cui W W, Liu X Y, Fang X J. Main, environmentally interacted and epistatic QTL for seed shape traits in soybean. Sci Agric Sin,2013, 46: 5081-5088 (in Chinese with English abstract)
[23] 閆寧, 謝尚潛, 耿青春, 徐宇, 李廣軍, 劉兵, 汪霞, 李其剛,章元明. 利用 Bayes分層廣義線性模型剖析大豆籽粒性狀的遺傳基礎. 作物學報, 2013, 39: 258-268 Yan N, Xie S Q, Geng Q C, Xu Y, Li G J, Liu B, Wang X, Li Q G,Zhang Y M. Genetic basics of seed traits in soybean with bayes hierarchical gengralized linear model method. Acta Agron Sin,2013, 39: 258-268 (in Chinese with English abstract)
[24] 蘇成付, 趙團結(jié), 蓋鈞鎰. 不同統(tǒng)計遺傳模型 QTL定位方法應用效果的模擬比較. 作物學報, 2010, 36: 1100-1107 Su C F, Zhao T J, Gai J Y. Simulation comparisons of effectiveness among QTL mapping procedures of different statistical genetic models. Acta Agron Sin, 2010, 36: 1100-1107 (in Chinese with English abstract)
[25] 李杰勤, 張啟軍, 葉少平, 趙兵, 梁永書, 彭勇, 吳發(fā)強, 王世全, 李平. 四種不同 QTL作圖方法的比較研究. 作物學報,2005, 31: 1473-1477 Li J Q, Zhang Q J, Ye S P, Zhao B, Liang Y S, Peng Y, Wu F Q,Wang S Q, Li P. Comparative research on four mapping methods of QTLs. Acta Agron Sin, 2005, 31: 1473-1477 (in Chinese with English abstract)
[26] Wang S C, Basten C J, Zeng Z B. Windows QTL Cartographer 2.5. North Carolina State University, Raleigh, NC, USA,2001-2006
[27] Yang J, Zhu J, Williams R W. Mapping the genetic architecture of complex traits in experimental populations. Bioinformatics, 2007,23: 1527-1536
[28] Li H H, Ye G Y, Wang J K. A modified algorithm for the improvement of composite interval mapping. Genetics, 2007, 175:361-374
[29] Meng L, Li H H, Zhang L Y, Wang J K. QTL IciMapping: Integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. Crop J, 2015,3: 269-283
[30] Nyquist W E, Baker R J. Estimation of heritability and prediction of selection response in plant populations. Crit Rev Plant Sci,1991, 10: 235-322
[31] Lu W G, Wen Z X, Li H C, Yuan D H, Li J Y, Zhang H. Identification of the quantitative trait loci (QTL) underlying water soluble prote incontent in soybean. Theor Appl Genet, 2013, 126: 425-433
[32] 陳強. 大豆籽粒相關性狀 QTL定位分析. 河北科技師范學院碩士學位論文, 河北秦皇島, 2014 Chen Q. QTL Mapping for seed related traits in soybean (Glycine max L. Merr.). MS Dissertation of Hebei Normal University of Science & Technology, Qinhuangdao, China, 2014 (in Chinese with English abstract)
[33] 雷雅坤, 閆龍, 楊春燕, 宋曉坤, 張孟臣, 黃占景. 大豆公共遺傳圖譜C1連鎖群SSR標記空白區(qū)段的填補. 華北農(nóng)學報,2012, 27(6): 5-10
Lei Y K, Yan L, Yang C Y, Song X K, Zhang M C, Huang Z J. Complete the blank section with SSR markers on linkage group C1 of public genetic map in soybean. Acta Agric Boreali-Sin,2012, 27(6): 5-10 (in Chinese with English abstract)
[34] Wang S C, Basten C J, Zeng Z B. Windows QTL Cartographer V2.5. Department of Statistics. North Carolina: North Carolina State University, 2007
[35] Yang J, Zhu J. Predicting superior genotypes in multiple environments based on QTL effects. Theor Appl Genet, 2005, 110:1268-1274
[36] Yang J, Hu C C, Hu H, Yu R D, Xia Z, Ye X Z, Zhu J. QTL Network: mapping and visualizing genetic architecture of complex traits in experimental populations. Bioinformatics, 2008, 24:721-723
[37] 張紅梅, 李海朝, 文自翔, 顧和平, 袁星星, 陳華濤, 崔曉艷,陳新, 盧為國. 大豆籽粒維生素E含量的QTL分析. 作物學報,2015, 41: 187-196 Zhang H M, Li H C, Wen Z X, Gu H P, Yuan X X, Chen H T, Cui X Y, Chen X, Lu W G. Identification of QTL associated with vitamin e content in soybean seeds. Acta Agron Sin, 2015, 41:187-196 (in Chinese with English abstract)
[38] McCouch S, Chen X, Panaud O, Temnykh S, Xu Y, Cho Y G,Huang N, Ishii T, Blair M. Microsatellite marker development,mapping and applications in rice genetics and breeding. Plant Mol Biol, 1997, 35: 89-99
[39] Song Q J, Marek L F, Shoemaker R C, Lark K G, Concibido V C,Delannay X, Specht J E, Cregan P B. A new integrated genetic linkage map of the soybean. Theor Appl Genet, 2004, 109: 122-128
[40] Mansur L M, Orf J, Chase K, Jarvik T, Cregan P B, Lark K G. Genetic mapping of agronomictraits using recombi-nant inbred lines of soybean. Crop Sci, 1996, 36: 1327-1336
[41] 杜景紅, 樊葉楊, 吳季榮, 莊杰云. 水稻第 6染色體短臂產(chǎn)量性狀QTL簇的分解. 中國農(nóng)業(yè)科學, 2008, 41: 939-945 Du J H, Fan Y Y, Wu J R, Zhuang J Y. Dissection of QTLs for yield traits on the short arm of rice chromosome 6. Sci Agric Sin 2008, 41: 939-945 (in Chinese with English abstract)
Mapping Quantitative Trait Loci for Seed Size and Shape Traits in Soybean
CHEN Qiang**, YAN Long**, DENG Ying-Ying, XIAO Er-Ning, LIU Bing-Qiang, YANG Chun-Yan*, and ZHANG Meng-Chen*
Institute of Cereal and Oil Crops, Hebei Academy of Agriculture and Forestry Sciences / Shijiazhuang Branch of National Soybean Improvement Center / Huang-Huai-Hai Key Laboratory of Biology and Genetic Improvement of Soybean, Ministry of Agriculture, Shijiazhuang 050035, China
Seed size and shape not only relate to seed yield and quality, but also affect mechanical seeding in soybean (Glycine max L.). In this study, F6:8and F6:9populations derived from Jidou 12 × Heidou were used to analyze the genetic character and detect quantitative trait loci (QTLs) for seed length, seed width, seed thickness, seed length-to-width ratio, seed length-to-thickness ratio, and seed width-to-thickness ratio. Softwares WinQTLCart 2.5, QTLNetwork 2.1 and IciMapping 4.1 were used to identify the additive, epistatic and environmentally interacted QTLs for seed size and shape related traits. As results,the heritability of the six traits varied from 64.01% to 79.57%. A total of 38 additive QTLs were identified to be located on 12 chromosomes, with the heritability varying from 2.21% to 10.71%. Eight of them (qSL-17-1, qSL-18-1, qSW-6-1, qST-2-1,qST-6-1, qSLT-2-2, qSWT-2-1, and qSWT-20-1) were identified using three methods, simultaneously. In the meantime, seven pairs of additive × additive epistasis were detected and the heritability of epistasis pairs ranged from 0.78% to 6.20%. Additionally, the effects of QTL by environment interaction ranged from 0.0005% to 0.3900%. The QTLs identified using different mapping softwares in this study could provide a reliable theoretical basis for marker-assisted selection breeding.
Soybean; Seed shape; Quantitative trait locus; Epistasis
10.3724/SP.J.1006.2016.01309
本研究由國家自然科學基金項目(31471522), 河北省自然科學基金項目(C2015301012), 國家高技術研究發(fā)展計劃(863計劃)項目(2012AA101106), 國家現(xiàn)代農(nóng)業(yè)產(chǎn)業(yè)技術體系建設專項(CARS-004-PS06)和國家轉(zhuǎn)基因生物新品種培育重大專項(2014ZX0800402B)資助。
This study was supported by the National Natural Science Foundation of China (31471522), the Natural Science Foundation of Heibei Province (C2015301012), the National High-tech Research and Development Program of China (2012AA101106), the China Agriculture Research System (CARS-004-PS06), and the National Major Project for Developing New GM Crops (2014ZX0800402B).
(Corresponding authors): 張孟臣, E-mail: mengchenzhang@hotmail.com; 楊春燕, E-mail: chyyang66@163.com
**同等貢獻(Contributed equally to this work)
聯(lián)系方式: 陳強, E-mail: chenqiangwsm@163.com; 閆龍, E-mail: dragonyan1979@163.com
Received(): 2016-01-26; Accepted(接受日期): 2016-05-09; Published online(網(wǎng)絡出版日期): 2016-06-06.
URL: http://www.cnki.net/kcms/detail/11.1809.S.20160606.0855.006.html
猜你喜歡
今日農(nóng)業(yè)(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年5期)2022-08-30 01:58:26
今日農(nóng)業(yè)(2021年20期)2021-11-26 01:23:56
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
今日農(nóng)業(yè)(2021年14期)2021-10-14 08:35:34
中學生數(shù)理化·七年級數(shù)學人教版(2020年12期)2021-01-18 06:57:46
中學生數(shù)理化·七年級數(shù)學人教版(2020年12期)2021-01-18 06:57:46
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
