不同紫細菌光合基因表達調控機制異同
2016-10-13 06:21:49鄧春浩孫良昱陳坤明
生物技術通報 2016年2期
鄧春浩 孫良昱 陳坤明
(西北農林科技大學生命科學學院,楊凌 712100)
不同紫細菌光合基因表達調控機制異同
鄧春浩 孫良昱 陳坤明
(西北農林科技大學生命科學學院,楊凌 712100)
紫細菌是一類可以進行不放氧光合作用的原核微生物,可以利用光能產生ATP并為其生長代謝提供能量。其光系統由一系列色素及蛋白組成的復合體組成,并由一系列光合基因如puc、puf、bch和crt等編碼。紫細菌光合相關基因的表達主要受到外界氧化還原信號及光照的影響,然而不同紫細菌光合基因表達調控機制具有明顯的多樣性。以球形紅細菌與沼澤紅假單胞菌為重點,介紹了近年來幾種光合基因表達調控系統如PpsR/CrtJ型調控蛋白、雙組分磷酸化調控系統、CRP-FNR型調控蛋白等在紫細菌中的研究進展。通過系統深入分析這些調控通路在不同紫細菌中的特征及功能,發現不同菌株中類似調控通路通常具有相似功能,但又各具特點。旨為對進一步了解紫細菌光合基因表達調控機制并為其應用研究提供參考。
光合基因;球形紅細菌;沼澤紅假單胞菌;紫細菌
紫細菌是一類可以進行不放氧光合作用的光合微生物,種類繁多,不同種類紫細菌的光系統結構非常類似,都主要由各種色素與蛋白組成的復合體構成,存在于細菌質膜內嵌形成的細菌內膜系統(introcytoplasmic membrane,ICM)上,包括核心復合體(core-complex)和外圍天線色素復合體(peripheral antennae)。光能通過外圍天線色素復合體傳遞至核心復合體,并在光反應中心(reaction center,RC)引起電荷分離,最終將光能轉化為電能[1]。紫細菌光系統由一系列光合基因所編碼的蛋白和酶組成或催化合成,其中大部分基因集中在細菌基因組中,被稱為光合基因簇(photosynthesis gene cluster,PGC)的區域。人們對不同種紫細菌的光合基因表達調控機制研究發現,光合基因的表達均受到光照及細胞內氧化還原狀態的強烈影響,并通過一系列調控蛋白,如PpsR、PrrA(RegA)、FixJ、FnrL和 FixK等蛋白來影響puc、puf、puh、bch、crt和hem等光合基因的表達,然而在不同紫細菌類群中,這些光合基因簇表達調控的機制存在顯著的不同。本研究旨在通過分析不同紫細菌光合基因表達調控方式異同,從而深入了解光合基因表達調控機制。
表1介紹了球形紅細菌(Rb. sphaeroides)與沼澤紅假單胞菌(Rp. palustris)光系統主要調控因子及其功能異同。
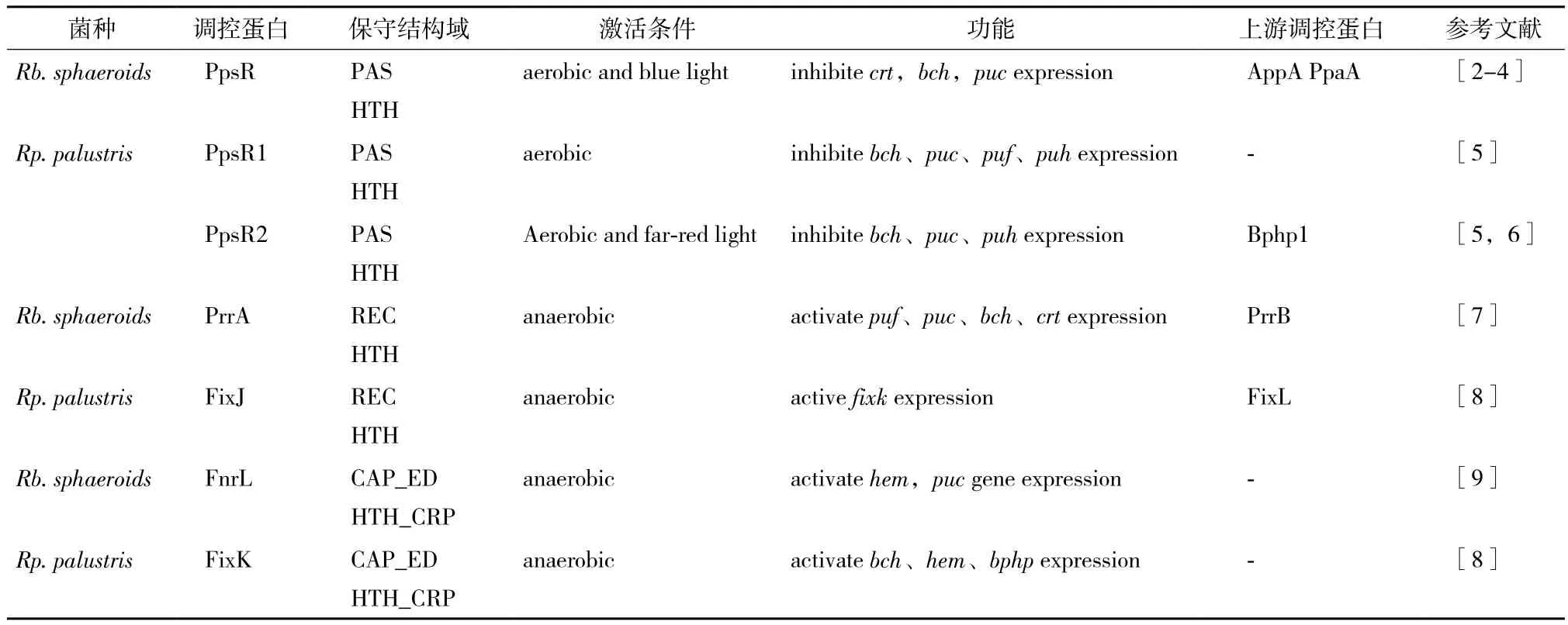
表1 Rhodobacter sphaeroides及Rhodopseudomonas palustris光系統主要調控因子及其功能異同
1 PpsR/CrtJ型調控蛋白
1.1 PpsR/CrtJ型調控蛋白的主要特征
PpsR/CrtJ是一類在紫細菌中普遍存在,并且非常重要的的調控光系統基因表達的蛋白,在不同紫細菌類群中具有相同的PAS(Per-arnt-sim)結構域和HTH(Helix-turn-helix)結構基元[5]。通過對不同紫細菌中與PpsR蛋白結合的目標序列的分析發現,PpsR一般與特定調控序列TGT-N12-ACA的回文區結合,受 PpsR調節的基因主要包括bch、crt、puc、hem等光合基因[10],然而不同紫細菌中PpsR蛋白調控基因表達的方式還有很大差異,有的菌株中只含有一種PpsR蛋白,有的菌株中含有兩種PpsR蛋白,下面將分別介紹不同菌株中PpsR蛋白調控模式。其中圖1比較了Rb. sphaeroides、Rb. capsulatus、Rp. palustris和Bradyrhizobium sp. ORS278中PpsR調控通路及其異同。
1.1.1 單PpsR調控模式 在Rb. sphaeroides與Rb. capsulatus中都只含有一種PpsR蛋白(Rb. capsulatus中為CrtJ),不同菌株中PpsR蛋白的序列與結構高度相似,一般含有3個半胱氨酸殘基(Cys),以四聚物形式存在,通過二硫鍵的形成與斷裂響應細胞內氧化還原狀態,并通過構象變化發揮調控功能[11],其中氧化態的PpsR蛋白有比還原態更強的DNA結合活性[12],二者都發揮阻遏蛋白的特性。在Rb. sphaeroides中存在的AppA蛋白,可以與PpsR蛋白結合,從而使PpsR不能與DNA結合而喪失其阻遏蛋白活性,因此二者在球形紅細菌中共同組成了阻遏-抗阻遏調控系統[13]。AppA的N末端具有FAD結合域及非共價結合的BluF,能夠響應氧化還原信號與光信號,在還原態及無藍光照射條件下可與PpsR結合;其C末端具有SCHIC(sensor containing heme instead of cobalamin)結構域,可以結合heme輔因子,能夠通過該輔因子與PpsR的PAS結構域結合[13-15]。近年來,隨著對AppA及PpsR蛋白晶體結構的解析,AppA與PpsR之間是如何互作的,響應氧化還原信號及光信號的分子機制也逐漸被解析[16-18],為人們進一步了解其分子調控機制并改造菌株提供了有利的支持。
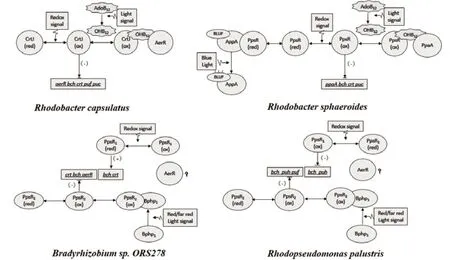
圖1 Rb. sphaeroides、Rb. capsulatus、Rp. palustris和Bradyrhizobium中PpsR調控通路[13-24]
在紫細菌中廣泛存在一類PpaA/AerR類蛋白(Rb. sphaeroides中被命名為PpaA,Rb. capsulatus中被命名為AerR),這類蛋白基因位置非常保守,通常位于光合基因簇中PpsR基因的上游[19]。這類蛋白沒有像AppA一樣的FAD及BluF功能位點,但C末端具有SCHIC結構域,也能通過heme輔因子來調控相關基因表達[19]。通過對ppaA與aerR缺失突變體研究發現,aerR突變體中puf、crtI、crtB和crtJ等光合基因表達上調,因此AerR的存在可能抑制光合基因的表達[20];而與之相反,ppaA突變體puc、crt和bch等光合基因表達下調,因此PpaA
可能促進光合基因表達[21]。然而,最近有研究發現,AerR在光照條件下通過結合B12輔因子,能夠與PpsR蛋白結合,從而使PpsR喪失阻遏蛋白活性,激活相關光合基因表達,因此二者也能夠組成阻遏-抗阻遏調控系統[22]。AerR的抗阻遏活性主要依賴于B12的狀態,在光照條件下,B12可以從腺苷鈷胺素(AdoB12)轉換為羥基鈷胺素(OHB12),而OHB12可以促使AerR與CrtJ的結合[22]。對于aerR缺失突變體中光合基因表達的下調,可能是由于AerR與PpsR基因是聯鎖表達的,aerR缺失突變使ppsR表達也下調,從而使光合基因在缺乏阻遏蛋白情況下表達上調,進而造成AerR抑制光合基因表達的假象[22]。
1.1.2 雙PpsR調控模式 與前兩種紫細菌不同,Rp. palustris和Bradyrhizobium sp. ORS278含有兩種類型的PpsR蛋白,即PpsR1和PpsR2。兩種紫細菌PpsR蛋白的氨基酸序列相似性較高,在Bradyrhizobium sp. ORS278中,PpsR調控蛋白主要調控bch、crt和cycA基因[23],而在Rp. palustris中,PpsR主要調節crt、bch和puc等基因的轉錄[5]。目前,Bradyrhizobium sp. ORS278中PpsR蛋白的功能最先得到研究。Bradyrhizobium sp. ORS278的PpsR1可以通過分子間的二硫鍵來響應氧化還原信號,在還原態以四聚物形式存在,氧化態以八聚物形式存在,其中還原態的四聚PpsR1可以與crt、bch等光合基因啟動子區的目標序列結合,執行轉錄激活作用[23];而其PpsR2蛋白沒有半胱氨酸殘基,不能識別氧化還原信號,但是可以通過細菌光敏色素(bacteriophytochrome,BphP)響應遠紅光信號,被遠紅光激活的BphP可以解除PpsR2對crt、bch及ppsr1等光合相關基因的抑制[23]。由于高度的序列相似性,最初Rp. palustris的兩個PpsR蛋白也被認為與Bradyrhizobium sp. ORS278的PpsRs有相似的功能[23],然而Braatsch等[5,6]通過對Rp. palustris CGA009的ppsR1突變體的研究發現,Rp. palustris的PpsR1在有氧條件下執行轉錄抑制功能;其PpsR2由于含有半胱氨酸殘基,不僅能通過RpBphP響應遠紅光信號而激活基因轉錄;同時,該PpsR2也可通過半胱氨酸殘基響應氧化還原信號。研究發現,Rp. palustris的PpsR2通過RpBphP1響應光信號,RpBphP1包含一個識別光信號的PCD結構域(input photosensory core domain,PCD)和一個傳遞信號的OTD結構域(output transducing domain,OTD),其中OTD結構域包含PAS/PAC結構,RpBphP1通過與RpPpsR2形成復合物來傳遞遠紅光信號[24]。在Rp. palustris和Bradyrhizobium sp. ORS278中也存在PpaA基因,然而其蛋白功能是否與Rb. sphaeroides與Rb. capsulatus中PpaA及AerR功能相似,還未見更多報道。
1.2 不同紫細菌中PpsR調控通路的異同
由于PpsR型調控通路是紫細菌中調控光合基因表達的最重要的調控通路之一,并且為近年來研究熱點,在許多紫細菌中均有較為深入的研究,對該通路中不同類型的調控蛋白按照其結構與功能分類,可以更清楚看到該調控通路的特點。PpsR/CrtJ型調控蛋白所屬調控通路中,Rb. sphaeroides含有兩個調節PpsR功能的調控蛋白AppA及PpaA,而Rb. capsulatus的PpsR/CrtJ上游調控蛋白僅有與PpaA同源的AerR;Bradyrhizobium sp. ORS278與Rp. palustris都含有兩種PpsR蛋白,其中Bradyrhizobium sp. ORS278的PpsR1執行轉錄激活功能,而Rp. palustris的PpsR1卻執行轉錄抑制功能。同時,這些調控蛋白在不同紫細菌中也存在一些共性特點:所有菌株中的PpsR蛋白都結合相似的TGT-N12-ACA回文區序列;Bradyrhizobium sp. ORS278與Rp. palustris的PpsR2同源,二者都通過Bphp受到光信號的調控;Rb. sphaeroides與Rb. capsulatus的AppA、PpaA及AerR都具有SCHIC結構域,并通過heme類輔因子與PpsR結合組成各種阻遏-抗阻遏系統。
2 雙組分磷酸化調控系統
2.1 雙組分磷酸化調控系統的主要特征
在Rb. sphaeroides與Rb. capsulatus中各自存在一套調控光合相關基因表達的雙組分磷酸化調控系統,分別為PrrB/A與RegB/A調控系統,二者調控功能相似。外界信號通過一個膜定位的具有組氨酸激酶活性的跨膜蛋白PrrB(RegB),將胞內另一個轉錄調控蛋白PrrA(RegA)磷酸化激活后影響一系列基因的轉錄,膜定位蛋白通常包含N末端的跨膜結構域及C末端組氨酸激酶結構域,胞內蛋白通常包含一個N末端的REC結構域(signal receiver domain)及C末端的HTH結構域。在Rp. palustris中同樣存在類似的雙組分磷酸化調控系統,人們主要研究了兩種與類似的與Rb. sphaeroides及Rb. capsulatus類似的調控系統,即RegS/R與FixL/J。
2.2 不同紫細菌中雙組分磷酸化調控系統功能異同
在Rb. sphaeroides與Rb. capsulatus中,該系統通過響應氧化還原信號,除調控固氮相關基因的表達外,還能夠調控puf、puc、bch和crt等光合基因的轉錄[25]。現有研究表明,PrrA/B(RegA/B)雙組分系統主要通過以下幾種可能的機制來識別氧化還原信號:Rb. capsulatus的RegB可以通過265位的Cys殘基來響應氧化還原信號,氧化態下通過形成分子間二硫鍵,促使分子變為無激酶活性的四聚體繼而阻斷下游通路[26];Swem等[27]發現Rb. capsulatus的RegB的跨膜區含有輔酶Q結合位點,醌池(ubiquinone pool)的氧化還原狀態能夠調節RegB的激酶活性,氧化態的輔酶Q可以影響RegB的激酶活性繼而阻斷下游通路;而Kim等[28]則認為Rb. sphaeroides中PrrB的激酶活性主要受到cbb3氧化酶的調節,輔酶Q結合位點對于PrrB的激酶活性并沒有影響,cbb3氧化酶作為氧化還原信號受體,調節PrrB的磷酸酶及磷酸激酶活性平衡,還原態下使PrrB表現出激酶活性繼而激活下游通路[29]。近年來研究表明PrrC對于cbb3氧化酶銅中心的裝配密切相關,PrrC蛋白與PrrB/A系統密切相關的蛋白[30,31],其基因位置相鄰,然而對于氧化還原信號影響PrrB(RegB)激酶活性的具體機制仍未闡明。還原條件下,具有激酶活性的PrrB(RegB)可以磷酸化激活PrrA(RegA),而PrrA(RegA)通過識別特異性序列5'-GCTGGCGCGANNTATANNCGAC-3'而定位其所調控的基因。PrrA(RegA)對不同基因的調控方式也有所不同:可以直接與轉錄抑制因子PpsR(CrtJ)競爭bchC的啟動子區,并通過與RNA聚合酶互作而增加基因轉錄起始效率[32];可通過與其他蛋白(FnrL、CbbR和NtrYX等)協同作用增強相關基因轉錄[33-36];可通過與gpx(RSP2389)起始密碼子下游作用位點結合并抑制該基因轉錄[37];而亞精胺及DNA拓撲結構對PrrA與目的位點結合有重要影響,亞精胺可以提高光合基因轉錄效率,DNA的負超螺旋也有助于PrrA促進基因轉錄[7]。
Rp. palustris中的RegS/R雙組分調控系統是Rp. palustris中與PrrA/B親緣關系最近的雙組分調控系統,其中RegR與PrrA的相似性高達72%。但是通過突變體研究發現,RegS/R不參與光合自養、光合異養及固氮條件下的相關基因表達,只影響了吸氫酶的活性[38]。然而Rp. palustris中的另一套雙組分調控系統FixL/J,與PrrB/A親緣關系較遠,但可以參與光合基因調控。通過結構域分析可以發現FixL比PrrB多兩個PAS結構域,這導致二者接受氧化還原信號的機制可能不一樣。FixJ在厭氧條件下可以激活另外一個與光合基因調控相關的正調控蛋白FixK基因的轉錄,fixL/J突變體不能很好地在厭氧光照下生長[8]。然而在Rp. palustris中,氧化還原信號是如何傳遞給FixL的,有關FixJ對其他基因的調控作用,是否還有其他雙組分調控系統調控光合基因表達都還有待進一步研究。
3 CRP-FNR型調控蛋白
3.1 CRP-FNR型調控蛋白的主要特征
紫細菌中還存在一類參與光合基因表達調控的CRP-FNR 型調控蛋白,該蛋白可以結合cNMP,以其含有的4Fe-4S簇響應氧化還原信號,并可以通過與DNA的結合來調控相關基因的表達[39],現有研究表明不同菌株中的CRP-FNR型調控蛋白主要調控啟動子區具有保守的FNR調控序列(TTGAT-N4-ATCAA)的基因[8,9]。
3.2 不同CRP-FNR型調控蛋白功能異同
在Rb. sphaeroides中調控光合基因的CRP-FNR型調控蛋白為FnrL。在低氧分壓下,還原態的FnrL能結合并激活具有保守FNR調控序列的光合相關基因,如puc、hemA、hemF、hemZ和bchE等[9,40]。FnrL是Rb. sphaeroides的一個關鍵調控因子,其fnrL突變體不能在光照條件下生長,不能在黑暗厭氧條件下利用DMSO作為電子傳遞受體[9],也不能形成正常的內膜系統[41]。Rb. sphaeroides中FnrL蛋白經常與其他蛋白共同調控基因表達,如puc基因啟動子區的FNR調控序列與其IHF調控區域有重合;hemA上游雙啟動子也受到PrrA及FnrL的共同調節[40]。Rb. capsulatus的FnrL蛋白與Rb. sphaeroides的親緣關系最高,但是二者的功能有很大的差異。Rb. capsulatus的fnrL突變使菌株不能在黑暗厭氧條件下利用DMSO,但是對菌株厭氧與有氧條件下的光合生長沒有影響,也不影響細胞內膜系統的形成[41];對Rb. capsulatus的hem啟動子的深入分析發現,hemE在半好氧條件下能夠被FnrL明顯激活,而hemB的表達則受到FnrL蛋白的抑制,但FnrL對其他hem基因表達的影響則非常微弱[42]。
在Rp. palustris中存在多種CRP-FNR家族蛋白(圖2),其中AadR與FixK兩種蛋白與Rb. sphaeroides與Rb. capsulatus的FnrL相似性最高。研究發現AadR主要參與苯酸鹽代謝,但不參與光合基因表達調控[43]。而FixK的功能則與Rb. sphaeroides的FnrL類似,識別相似的調控序列,都調控電子傳遞及光系統相關基因的表達[8]。FixK可以通過直接或者間接的方式與基因上游特異調控序列結合而調節基因轉錄,在厭氧或微氧條件下,FixL可以激活fixK表達,FixK接著可以激活一系列基因如bch、hem、cooNOQP及pioABC等的表達[8,44]。

圖2 Rp. palustris中CRP-FNR型調控蛋白系統發育分析
4 SPB調控因子
SPB是Rb. sphaeroides中的一個調節蛋白,包含一個C端的K-box和N端負責DNA結合的H-NS結構,能識別puf和puc操縱子的操縱區,在強光照射下抑制puf和puc的轉錄,因此又叫作高光抑制因子[45]。SPB缺失突變使菌株在光照條件下Bchl表達上調,同時也可提高菌株的產氫效率[46]。SPB蛋白不能響應氧化還原信號,也不包含能識別光信號的基團。2007年,Shimada等[47]通過免疫共沉淀技術,分離得到了一個與SPB相互作用的蛋白SIP,SIP同樣屬于H-NS家族的蛋白,但有關強光信號傳遞給SPB的具體機制,仍然沒有闡明。在Rb. capsulatus中存在一個可以響應光信號并具有H-NS結構的蛋白——HvrA[48],但是與SPB不同的是HvrA蛋白負責在低光照下激活puf和puh的轉錄,hvrA缺失菌株表現出在中低光照下光合效率下降[48]。與SPB一樣,HvrA蛋白響應光照的機理仍未闡明。
在Rp. palustris中細菌光敏色素RpBphP2與RpBphP3可以響應光照強度信號,繼而調控光合基因表達[49],但是在Rp. palustris中未發現H-NS家族蛋白,對于Rb. sphaeroides和Rb. capsulatus中是否存在類似的調控蛋白還有待進一步研究。
5 非編碼小RNA(small-noncoding RNA)調控
非編碼小RNA廣泛參與微生物細胞代謝、應激反應等生理過程,近些年來受到人們廣泛重視。2009年,Borghoff等[50]用高濃度氧氣處理Rb. sphaeroides菌株后,通過高通量測序及Northern blot技術,獲得了20個響應氧化還原狀態的非編碼小RNA。隨后作者所在課題組選取其中一個僅在Rb. sphaeroides中特異存在的非編碼小RNA-PcrZ,通過一系列研究發現pcrZ在厭氧條件下被PrrA激活轉錄形成小RNA-PcrZ,而PcrZ可以通過互補配對的形式與puc、bch等基因結合而抑制光合基因表達[51]。在這個調控模式里,PcrZ還可與轉錄激活因子PrrA競爭性結合目的基因,從而實現對基因表達的精細調節。
在Rp. palustris中,2012年,Hirakawa等[52]通過RNAseq技術得到了一系列反義RNA,并鑒定出其中一個反義RNA具有調節群體信號感應受體表達的功能。然而,目前關于小RNA調控Rp. palustris光系統方面的研究,尚未見更多報道。
6 結語
紫細菌光合基因表達調控是一個非常復雜的網絡,除本文中詳細對比的幾種調控通路及其所含的調控蛋白外,還有很多調控蛋白參與光合基因表達調控。例如,Rb. sphaeroides中的外膜蛋白TspO參與AppA-PpsR調控系統中氧化還原信號向AppA的傳遞[53];Rb. sphaeroides與Rb. capsulatus中存在的硫氧還原蛋白可通過調節DNA促旋酶的活性,改變DNA的超螺旋狀態,繼而影響光合基因表達[54];紫細菌中還普遍存在一類細菌光敏色素(bacteriophytochrome,Bphp),可以通過響應光信號而調節光合基因表達[55];而另外一類向光素依賴蛋白(phototropin-related proteins)也可通過響應光信號調控光合基因表達[55]。但是這些調控蛋白及其所屬調控通路中,光信號及氧化還原信號是如何傳遞的,下游調控蛋白是如何識別并調控基因的,不同紫細菌中是否具有相似的調控模式,都還有待進一步研究。
值得注意的是,即使氨基酸序列最為相近的蛋白,其突變體在不同紫細菌中可能具有不同的表型;相似性較低的蛋白,其突變體在不同紫細菌中也表現出相似的功能。例如,Rp. palustris中與PrrA/B(RegA/B)最為相似的RegS/R調控系統,并不參與光系統形成,而后來發現的FixL/J調控通路卻參與光合系統相關基因調控;Rb. sphaeroides與Rb. capsulatus的FnrL相似性很高,但其突變體卻表現出不同的表型,相似性較低的Rp. palustris的FixK與Rb. sphaeroides的FnrL之間,在功能上卻最為相近,都具有相似的調控序列及調控機制。不同紫細菌光合基因表達調控模式都有其共性及特性,而通過對不同類群紫細菌光合基因表達調控機制相似性研究,有助于人們進一步深入了解紫細菌光合基因表達調控機制,并在其他類型光合細菌中發現類似的調控通路。
[1]Roszak AW, Howard TD, Southall J, et al. Crystal structure of the RC-LH1 core complex from Rhodopseudomonas palustris[J]. Science, 2003, 302(5652):1969-1972.
[2]Penfold RJ, Pemberton JM. Sequencing, chromosomal inactivation,and functional expression in Escherichia coli of ppsR, a gene which represses carotenoid and bacteriochlorophyll synthesis in Rhodobacter sphaeroides[J]. J Bacteriol, 1994, 176(10):2869-2876.
[3] Gomelsky M, Kaplan S. Genetic evidence that PpsR from Rhodobacter sphaeroides 2.4.1 functions as a repressor of puc and bchF expression[J]. J Bacteriol, 1995, 177(6):1634-1637.
[4]Gomelsky M, Kaplan S. AppA, a redox regulator of photosystem formation in Rhodobacter sphaeroides 2.4.1, is a flavoprotein. Identification of a novel fad binding domain[J]. J Biol Chem,1998, 273(52):35319-35325.
[5]Braatsch S, Bernstein JR, Lessner F, et al. Rhodopseudomonas palustris CGA009 has two functional ppsR genes, each of which encodes a repressor of photosynthesis gene expression[J]. Biochemistry, 2006, 45(48):14441-14451.
[6]Braatsch S, Johnson JA, Noll K, et al. The O2-responsive repressor PpsR2 but not PpsR1 transduces a light signal sensed by the BphP1 phytochrome in Rhodopseudomonas palustris CGA009[J]. FEMS Microbiol Lett, 2007, 272(1):60-64.
[7]Eraso JM, Kaplan S. Regulation of gene expression by PrrA in Rhodobacter sphaeroides 2.4.1:role of polyamines and DNA topology[J]. J Bacteriol, 2009, 191(13):4341-4352.
[8]Rey FE, Harwood CS. FixK, a global regulator of microaerobic growth, controls photosynthesis in Rhodopseudomonas palustris[J]. Mol Microbiol, 2010, 75(4):1007-1020.
[9]Zeilstra-Ryalls JH, Kaplan S. Aerobic and anaerobic regulation in Rhodobacter sphaeroides 2.4.1:the role of the fnrL gene[J]. J Bacteriol, 1995, 177(22):6422-6431.
[10] Elsen S, Jaubert M, Pignol D, et al. PpsR:a multifaceted regulator of photosynthesis gene expression in purple bacteria[J]. Mol Microbiol, 2005, 57(1):17-26.
[11] Choudhary M, Kaplan S. DNA sequence analysis of the photosynthesis region of Rhodobacter sphaeroides 2.4.1[J]. Nucleic Acids Res, 2000, 28(4):862-867.
[12]Masuda S, Bauer CE. AppA is a blue light photoreceptor that antirepresses photosynthesis gene expression in Rhodobacter sphaeroides[J]. Cell, 2002, 110(5):613-623.
[13]趙志平. 紫細菌光合機構及光合作用基因的表達調控[J].微生物學通報, 2008, 35(5):830-835.
[14]Han Y, Meyer MH, Keusgen M, et al. A haem cofactor is required for redox and light signalling by the AppA protein of Rhodobacter sphaeroides[J]. Mol Microbiol, 2007, 64(4):1090-1104.
[15]Yin L, Dragnea V, Bauer CE. PpsR, a regulator of heme and bacteriochlorophyll biosynthesis, is a heme-sensing protein[J]. J Biol Chem, 2012, 287(17):13850-13858.
[16]Pandey R, Flockerzi D, Hauser Marcus JB, et al. Modeling the Light- and Redox-Dependent Interaction of PpsR/AppA in Rhodobacter sphaeroides[J]. Biophys J, 2011, 100(10):2347-2355.
[17]Winkler A, Heintz U, Lindner R, et al. A ternary AppA-PpsR-DNA complex mediates light regulation of photosynthesis-related gene expression[J]. Nat Struct Mol Biol, 2013, 20(7):859-867.
[18]Heintz U, Meinhart A, Winkler A. Multi-PAS domain-mediatedprotein oligomerization of PpsR from Rhodobacter sphaeroides[J]. Acta Crystallogr D Biol Crystallogr, 2014, 70(Pt 3):863-876.
[19]Moskvin OV, Gilles-Gonzalez MA, Gomelsky M. The PpaA/AerR regulators of photosynthesis gene expression from anoxygenic phototrophic proteobacteria contain heme-binding SCHIC domains[J]. J Bacteriol, 2010, 192(19):5253-5256.
[20]Dong C, Elsen S, Swem LR, et al. AerR, a second aerobic repressor of photosynthesis gene expression in Rhodobacter capsulatus[J]. J Bacteriol, 2002, 184(10):2805-2814.
[21]Gomelsky L, Sram J, Moskvin OV, et al. Identification and in vivo characterization of PpaA, a regulator of photosystem formation in Rhodobacter sphaeroides[J]. Microbiology, 2003, 149(Pt 2):377-388.
[22]Cheng Z, Li K, Hammad LA, et al. Vitamin B12 regulates photosystem gene expression via the CrtJ antirepressor AerR in Rhodobacter capsulatus[J]. Mol Microbiol, 2014, 91(4):649-664.
[23]Jaubert M, Zappa S, Fardoux J, et al. Light and redox control of photosynthesis gene expression in Bradyrhizobium:dual roles of two PpsR[J]. J Biol Chem, 2004, 279(43):44407-44416.
[24]Bellini D, Papiz MZ. Structure of a bacteriophytochrome and light-stimulated protomer swapping with a gene repressor[J]. Structure, 2012, 20(8):1436-1446.
[25]Willett J, Smart JL, Bauer CE. RegA control of bacteriochlorophyll and carotenoid synthesis in Rhodobacter capsulatus[J]. J Bacteriol, 2007, 189(21):7765-7773.
[26]Swem LR, Kraft BJ, Swem DL, et al. Signal transduction by the global regulator RegB is mediated by a redox-active cysteine[J]. EMBO J, 2003, 22(18):4699-4708.
[27]Swem LR, Gong X, Yu CA, et al. Identification of a ubiquinonebinding site that affects autophosphorylation of the sensor kinase RegB[J]. J Biol Chem, 2006, 281(10):6768-6775.
[28]Kim YJ, Ko IJ, Lee JM, et al. Dominant role of the cbb3 oxidase in regulation of photosynthesis gene expression through the PrrBA system in Rhodobacter sphaeroides 2.4.1[J]. J Bacteriol, 2007,189(15):5617-5625.
[29]Oh JI, Ko IJ, Kaplan S. Reconstitution of the Rhodobacter sphaeroides cbb3-PrrBA signal transduction pathway in vitro[J]. Biochemistry, 2004, 43(24):7915-7923.
[30]Badrick AC, Hamilton AJ, Bernhardt PV, et al. PrrC, a Sco homologue from Rhodobacter sphaeroides, possesses thiol-disulfide oxidoreductase activity[J]. FEBS Lett, 2007, 581(24):4663-4667.
[31] Thompson AK, Gray J, Liu A, et al. The roles of Rhodobacter sphaeroides copper chaperones PCu(A)C and Sco(PrrC)in the assembly of the copper centers of the aa(3)-type and the cbb(3)-type cytochrome c oxidases[J]. Biochim Biophys Acta,2012, 1817(6):955-964.
[32]Bowman WC, Du S, Bauer CE, et al. In vitro activation and repression of photosynthesis gene transcription in Rhodobacter capsulatus[J]. Mol Microbiol, 1999, 33(2):429-437.
[33]Ranson-Olson B, Jones DF, Donohue TJ, et al. In vitro and in vivo analysis of the role of PrrA in Rhodobacter sphaeroides 2.4.1 hemA gene expression[J]. J Bacteriol, 2006, 188(9):3208-3218.
[34]Dangel AW, Tabita FR. Protein-protein interactions between CbbR and RegA(PrrA), transcriptional regulators of the cbb operons of Rhodobacter sphaeroides[J]. Mol Microbiol, 2009, 71(3):717-729.
[35]Dangel AW, Luther A, Tabita FR. Amino acid residues of RegA important for interactions with the CbbR-DNA complex of Rhodobacter sphaeroides[J]. J Bacteriol, 2014, 196(17):3179-3190.
[36]Carrica Mdel C, Fernandez I, Sieira R, et al. The two-component systems PrrBA and NtrYX co-ordinately regulate the adaptation of Brucella abortus to an oxygen-limited environment[J]. Mol Microbiol, 2013, 88(2):222-233.
[37]Eraso JM, Roh JH, Zeng X, et al. Role of the global transcriptional regulator PrrA in Rhodobacter sphaeroides 2.4.1:combined transcriptome and proteome analysis[J]. J Bacteriol, 2008, 190(14):4831-4848.
[38]Rey FE, Oda Y, Harwood CS. Regulation of uptake hydrogenase and effects of hydrogen utilization on gene expression in Rhodopseudomonas palustris[J]. J Bacteriol, 2006, 188(17):6143-6152.
[39]Ouchane S, Picaud M, Therizols P, et al. Global regulation of photosynthesis and respiration by FnrL:the first two targets in the tetrapyrrole pathway[J]. J Biol Chem, 2007, 282(10):7690-7699.
[40]Ranson-Olson B, Zeilstra-Ryalls JH. Regulation of the Rhodobacter sphaeroides 2.4.1 hemA gene by PrrA and FnrL[J]. J Bacteriol,2008, 190(20):6769-6778.
[41]Fedotova Y, Zeilstra-Ryalls J. Analysis of the role of PrrA,PpsR, and FnrL in intracytoplasmic membrane differentiation of Rhodobacter sphaeroides 2.4.1 using transmission electron microscopy[J]. Photosynth Res, 2014, 119(3):283-290.
[42]Smart JL, Willett JW, Bauer CE. Regulation of hem gene expression in Rhodobacter capsulatus by redox and photosystem regulators RegA, CrtJ, FnrL, and AerR[J]. J Mol Biol, 2004, 342(4):1171-1186.
[43]Dispensa M, Thomas CT, Kim MK, et al. Anaerobic growth of Rhodopseudomonas palustris on 4-hydroxybenzoate is dependent on AadR, a member of the cyclic AMP receptor protein family of transcriptional regulators[J]. J Bacteriol, 1992, 174(18):5803-5813.
[44]Bose A, Newman DK. Regulation of the phototrophic iron oxidation(pio)genes in Rhodopseudomonas palustris TIE-1 is mediated by the global regulator, FixK[J]. Mol Microbiol, 2011, 79(1):63-75.
[45]Nishimura K, Shimada H, Hatanaka S, et al. Growth, pigmentation,and expression of the puf and puc operons in a light-respondingrepressor(SPB)-disrupted Rhodobacter sphaeroides[J]. Plant Cell Physiol, 1998, 39(4):411-417.
[46]Wang X, Yang H, Zhang Y, et al. Remarkable enhancement on hydrogen production performance of Rhodobacter sphaeroides by disrupting spbA and hupSL genes[J]. International Journal of Hydrogen Energy, 2014, 39(27):14633-14641.
[47]Shimada H, Ishida K, Machiya Y, et al. Isolation of SIP, a protein that interacts with SPB, a possible transcriptional regulatory factor in Rhodobacter sphaeroides[J]. Plant Cell Physiol, 2007, 48(10):1504-1508.
[48]Buggy JJ, Sganga MW, Bauer CE. Characterization of a lightresponding trans-activator responsible for differentially controlling reaction center and light-harvesting-I gene expression in Rhodobacter capsulatus[J]. J Bacteriol, 1994, 176(22):6936-6943.
[49] Fixen KR, Baker AW, Stojkovic EA, et al. Apo-bacteriophytochromes modulate bacterial photosynthesis in response to low light[J]. Proc Natl Acad Sci USA, 2014, 111(2):E237-244.
[50] Berghoff BA, Glaeser J, Sharma CM, et al. Photooxidative stressinduced and abundant small RNAs in Rhodobacter sphaeroides[J]. Mol Microbiol, 2009, 74(6):1497-1512.
[51] Mank NN, Berghoff BA, Hermanns YN, et al. Regulation of bacterial photosynthesis genes by the small noncoding RNA PcrZ[J]. Proc Natl Acad Sci USA, 2012, 109(40):16306-16311.
[52] Hirakawa H, Harwood CS, Pechter KB, et al. Antisense RNA that affects Rhodopseudomonas palustris quorum-sensing signal receptor expression[J]. Proc Natl Acad Sci USA, 2012, 109(30):12141-12146.
[53]Zeng X, Kaplan S. TspO as a modulator of the repressor/antirepressor(PpsR/AppA)regulatory system in Rhodobacter sphaeroides 2.4.1[J]. J Bacteriol, 2001, 183(21):6355-6364.
[54]Li K, Pasternak C, Hartig E, et al. Thioredoxin can influence gene expression by affecting gyrase activity[J]. Nucleic Acids Res,2004, 32(15):4563-4575.
[55]Evans K, Georgiou T, Hillon T, et al. The purple phototrophic bacteria[M]. Springer Netherlands, 2009:799-809.
(責任編輯 狄艷紅)
Differences in Regulation of Photosynthetic Gene Expression Between Different Purple Bacteria
DENG Chun-hao SUN Liang-yu CHEN Kun-ming
(College of Life Sciencess,Northwest A&F University,Yangling 712100)
The purple bacteria are prokaryotes that can proceed photosynthesis without oxygen release. They use light energy to produce ATP for their own growth and metabolism. The photosystem of a purple bacterium consists of a series of complex of pigments and proteins encoded by photosynthetic genes like puc, puf, puh, bch, crt and so on. The expressions of their photosynthetic genes mainly are influenced by light and external redox signals;however, the expression and regulation mode of photosynthetic genes are greatly different between different kinds of purple bacteria. This review introduces the research advances on several expression and regulation systems of photosynthetic genes, such as PpsR/CrtJ like regulator, two-component phosphorylation regulation system and CRP-FNR like regulator in purple bacteria while focusing on Rhodobacter sphaeroides and Rhodopseudomonas palustris. The results from the functional and characteristic analysis of the regulation mechanism in different purple bacteria suggest that, each regulation system existing in different bacterial strain shows similar functions but with somewhat unique characteristics. This study gives us further insights for understanding the mechanisms of expression and regulation of photosynthetic genes in the purple bacteria, and provides important
for application of these purple bacteria in the future.
photosynthetic genes;Rhodobacter sphaeroides;Rhodopseudomonas palustris;purple bacteria
10.13560/j.cnki.biotech.bull.1985.2016.02.005
2015-04-22
教育部新世紀優秀人才支持計劃項目(NCET-11-0440),教育部留學回國人員科研啟動費
鄧春浩,男,碩士,研究方向:細胞生物學;E-mail:dchdr@163.com
陳坤明,男,博士,研究方向:植物細胞逆境分子生理與調控,光合作用與生物能源;E-mail:kunmingchen@nwsuaf.edu.cn
猜你喜歡
工業設計(2022年8期)2022-09-09 07:43:20
鴨綠江(2021年35期)2021-04-19 12:24:18
軍民兩用技術與產品(2021年10期)2021-03-16 06:05:30
北京測繪(2020年12期)2020-12-29 01:33:58
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
裝備制造技術(2019年12期)2019-12-25 03:06:46
中國洗滌用品工業(2019年4期)2019-05-11 09:27:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
家庭影院技術(2017年9期)2017-09-26 03:41:45
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
