液相色譜—質譜聯用分析流感病毒感染引起宿主蛋白類泛素化修飾
2016-10-21 11:12:42彭其勝李光譜孫萬春楊靜波全桂花劉寧
分析化學 2016年6期
關鍵詞:分析
彭其勝 李光譜 孫萬春 楊靜波 全桂花 劉 寧
摘要干擾素刺激基因15編碼蛋白質(Interferon stimulated gene 15 kDa protein, ISG15)是最早被鑒定的類泛素分子蛋白質,在病毒感染和免疫調節等方面具有重要作用。本研究利用免疫沉淀技術將被類泛素ISG15修飾的蛋白富集純化,采用液相色譜質譜聯用技術對流感病毒感染A549宿主細胞過程中產生的類泛素ISG15修飾蛋白進行了分析。實驗結果表明,在流感病毒感染的實驗組A549細胞中,鑒定到了22種來源于宿主細胞的ISG15修飾的蛋白,包括類泛素蛋白ISG15、細胞周期蛋白T1、熱休克蛋白71、鈣調素結合蛋白、真核翻譯起始因子等,以及1種來源于流感病毒的非結構蛋白NS1。在鑒定的22種宿主蛋白中,有6種蛋白在未感染病毒的對照組A549細胞中也得到鑒定,包括膜聯蛋白A1、果糖二磷酸醛縮酶A、線粒體三磷酸腺苷合成酶亞基g、烯醇化酶、肌動蛋白、微管蛋白。生物信息學分析表明,流感病毒感染引起的ISG15修飾的宿主蛋白分別歸屬于9個不同的蛋白分類,包括細胞骨架蛋白、分子伴侶蛋白、酶調節劑、核酸結合蛋白、激酶類、轉移酶類、轉錄因子、氧化還原酶類以及結構蛋白。本研究為大規模分析鑒定ISG15修飾蛋白提供了一種特異、有效的研究方法。
關鍵詞 流感病毒; 類泛素蛋白; 干擾素刺激基因15編碼蛋白質; 液相色譜質譜聯用
1引言
流感病毒等微生物在宿主細胞中有效復制和拮抗宿主免疫反應涉及到細胞內的多種重要信號轉導通路,如WNT[1], NFКB[2], MAPK[3], PI3K/Akt[4]等,是一個非常復雜的生物學過程。在此過程中,蛋白質翻譯后修飾如泛素和類泛素化修飾起到了十分重要的作用。泛素和類泛素化修飾是一類最富誘導性和可逆性的蛋白修飾[5]。其中,干擾素刺激基因15蛋白 (Interferon stimulated gene 15 kDa protein,ISG15) 類泛素化修飾在調節病原微生物的感染和復制、宿主細胞先天免疫反應等過程中發揮著極其重要的作用[6,7]。ISG15是由干擾素誘導產生的一種分子量約為15 kDa的蛋白質,由兩個泛素樣的小亞基通過一個短鉸鏈區連接組成[8]。ISG15對蛋白質的修飾作用是可逆的共價修飾,可以調節底物蛋白的穩定性、功能活性狀態以及細胞內定位等[9]。研究表明,ISG15缺失的小鼠對包括流感病毒在內的許多病毒感染的易受性明顯高于野生型小鼠[10]。進一步的研究發現,ISG15修飾能夠在很大程度上抑制流感病毒在人源細胞中的基因表達和復制,而在小鼠胚胎成纖維細胞中卻沒有觀察到明顯的類似的抑制作用[11]。這說明,類泛素修飾ISG15在流感病毒感染人源細胞過程中發揮著極其重要的作用。因此,分析鑒定流感病毒感染引起人源宿主細胞中類泛素ISG15修飾蛋白的變化,對于深入理解人源宿主通過ISG15抵抗病毒感染的分子機制具有重要意義。
現代質譜技術在蛋白質結構分析和大規模蛋白質組分析等領域得到了越來越廣泛的應用。相對于其它分析技術,質譜技術具有高分辨率和高靈敏度等優勢,逐漸成為多肽、蛋白質分子結構分析研究中最為重要的技術手段之一[12~15]。而免疫沉淀技術是基于高特異性的抗體抗原反應,利用抗體將復雜混合物中的抗原分子特異性富集的一種樣品制備技術。目前,將免疫分離技術與質譜分析技術相結合,已經成為對復雜生物樣品進行定性、半定量、甚至定量分析的強有力手段。這種分析技術將傳統免疫分析技術的高度特異性和質譜技術的高度靈敏性、高分辨率等特性相結合,可以對復雜生物樣品如具有各種翻譯后修飾的蛋白質進行分析鑒定[16,17]。
目前,大規模分析泛素或類泛素修飾的研究主要集中于對泛素(Ubiquitin)和類泛素(SUMO、ISG15)修飾蛋白在各種生物學體系中的表征[18~21]。對于流感病毒感染人源宿主細胞過程中ISG15修飾蛋白的大規模分析研究未見報導。在本研究中,采用免疫沉淀技術與液相色譜質譜聯用技術,對流感病毒感染宿主細胞過程中的ISG15修飾蛋白進行分析鑒定。首先將人流感病毒株H3N2接種到來源于人肺腺癌上皮的細胞系A549中,感染一定時間后,利用免疫沉淀技術將被類泛素ISG15修飾的蛋白富集純化,利用液相色譜質譜聯用技術對富集蛋白進行分析,成功鑒定到了22種來源于宿主細胞的蛋白(包括類泛素ISG15蛋白)和1種流感病毒非結構蛋白。結果表明,流感病毒感染能夠引起多種宿主蛋白的類泛素ISG15修飾,而免疫沉淀結合液相色譜質譜聯用技術可以非常有效地對類泛素ISG15修飾蛋白進行分析鑒定。
2實驗部分
2.1儀器與試劑
LTQ Orbitrap電場軌道阱回旋共振組合質譜儀(美國Thermo公司);BIO Wide Pore C18 反相毛細管色譜柱(150 mm×0.18 mm, 5 μm, 美國Sigma公司);MiniPROTEAN Tetra Cell垂直電泳及Mini TransBlot Cell電轉系統(美國BioRad公司);Allegra TM X22R離心機(美國Beckman Coulter公司);Microchemi Chemiluminescence system 4.2凝膠成像儀(以色列DNR BioImaging Systems公司);Varioskan Flash酶標儀(美國Thermo公司);四維旋轉混合儀(海門其林貝爾公司)。
Bradford蛋白定量試劑盒、尿素、碳酸氫銨、二硫蘇糖醇、碘乙酰胺(美國BioRad公司);測序級TPCK修飾的胰蛋白酶(美國Promega公司);免疫沉淀試劑盒Classic IP Kit(美國Pierce公司);甲酸、乙腈、β巰基乙醇(美國Sigma公司);抗ISG15單克隆抗體、HRP標記的兔抗鼠二抗(美國Santa Cruz公司);DMEM培養基美國(Hyclone公司);胎牛血清(美國Gibco公司);牛血清白蛋白(BSA)、聚偏氟乙烯(PVDF)膜(美國Millipore公司);增強化學發光底物試劑(ECL Prime,瑞典GE Healthcare公司);其它試劑均為國產分析純。人肺腺癌上皮的細胞系A549及人流感病毒株H3N2由本實驗室保存;實驗用水為經MilliQ純水儀(美國Millipore公司)制備的超純水。
2.2病毒接種與感染A549細胞
用含血清的DMEM培養基將A549細胞培養至80%~90%成片狀態,棄去培養基,用PBS清洗3遍。 加入含0.2%胰酶的無血清DMEM培養基繼續培養2 h后,接種經適當稀釋的流感病毒株H3N2。感染24 h后,棄去培養上清液,收集細胞。使用IP裂解液(pH 7.4, 25 mmol/L Tris, 150 mmol/L NaCl, 1 mmol/L EDTA, 1% NP40, 5% Glycerol)裂解細胞,于4℃、12000 r/min離心20 min,取上清液,分裝后置
Symbolm@@ 80℃保存。使用Bradford蛋白定量試劑盒測定蛋白濃度。
2.3免疫沉淀技術富集ISG15修飾蛋白
采用Pierce公司Classic IP Kit免疫沉淀試劑盒進行蛋白的富集。首先,在細胞裂解液上清液中加入未偶聯抗體的瓊脂糖微球(表面包被有蛋白A/蛋白G)進行預處理,4℃旋轉振搖過夜,離心,收集上清液。再按照免疫沉淀試劑盒說明書步驟,將抗ISG15單克隆抗體偶聯到瓊脂糖磁珠表面上。經充分清洗后,加入到經過預處理的細胞裂解液中。混勻后,4℃旋轉振搖過夜,離心收集瓊脂糖微球。經洗滌后,用洗脫緩沖液進行洗脫。對照組使用未經流感病毒感染的A549細胞,采用相同的方法富集內源性的ISG15修飾蛋白。將含有經免疫沉淀富集的ISG15修飾蛋白樣品分裝后保存于Symbolm@@ 80℃。取少量蛋白樣品加入2×SDS上樣緩沖液,95℃保溫5 min。用12% SDSPAGE進行分離,電泳結束后將蛋白電轉移到PVDF膜上。將PVDF膜清洗、用BSA封閉后,與抗ISG15單克隆抗體在室溫孵育2 h。將膜充分清洗后,再與HRP標記的兔抗鼠二抗在室溫孵育2 h。將PVDF膜充分清洗后,加上新混合的ECL(增強化學發光)底物,使用凝膠成像儀記錄蛋白印跡信號。
2.4蛋白質樣品的胰蛋白酶水解
在2.3小節中免疫沉淀富集的蛋白質樣品中加入5倍體積的預冷丙酮,
Symbolm@@ 20℃放置過夜后離心,棄去上清液。將沉淀用含有50 mmol/L NH4HCO3的2 mol/L尿素溶液溶解,加入終濃度50 mmol/L 二硫蘇糖醇溶液, 室溫放置60 min。再加入終濃度100 mmol/L 碘乙酰胺,避光反應60 min,將巰基烷基化。加入新配制的測序級TPCK胰蛋白酶溶液,37℃反應過夜后,加入適量10%甲酸終止反應。離心取上清液,凍干后于
Symbolm@@ 20℃保存。
2.5液相色譜質譜聯用分析鑒定蛋白質
使用液相色譜質譜聯用技術分析胰酶水解多肽樣品。將多肽樣品溶解在洗脫緩沖液A(99% 水+1%乙腈+0.1%甲酸)中, 經BIO Wide Pore C18 反相毛細管色譜柱(150 mm×0.18 mm, 5 μm)分離。梯度洗脫程序為: 0~180 min,5%~40% 緩沖液B(99% 乙腈+1% 水+0.1% 甲酸)。流速為0.25 μL/min。洗脫出的多肽直接進入LTQ Orbitrap質譜離子源進行分析。毛細管溫度200℃,噴霧電壓為1.5 kV,質核比檢測范圍為400~2000 amu。應用數據依賴方式進行二級質譜掃描,每個全掃描后進行5個二級掃描,碰撞能量為35%歸一化能量,母離子窗口寬度為2 Da,持續動態排除時間為1 min。將采集到的質譜數據傳輸到數據處理工作站,并使用Thermo Proteome Discoverer 1.1質譜數據分析軟件進行分析。蛋白質數據庫檢索時采用SEQUEST模式,在Uniprot蛋白質數據庫進行檢索。
2.6生物信息學分析
參照文獻[22,23]的方法,使用PANTHER (Protein annotation through evolutionary relationship)分析軟件對得到的ISG15修飾蛋白質進行分析,將所鑒定的蛋白按照不同的生物學功能進行分類。
3結果與討論
3.1ISG15修飾蛋白的免疫沉淀富集
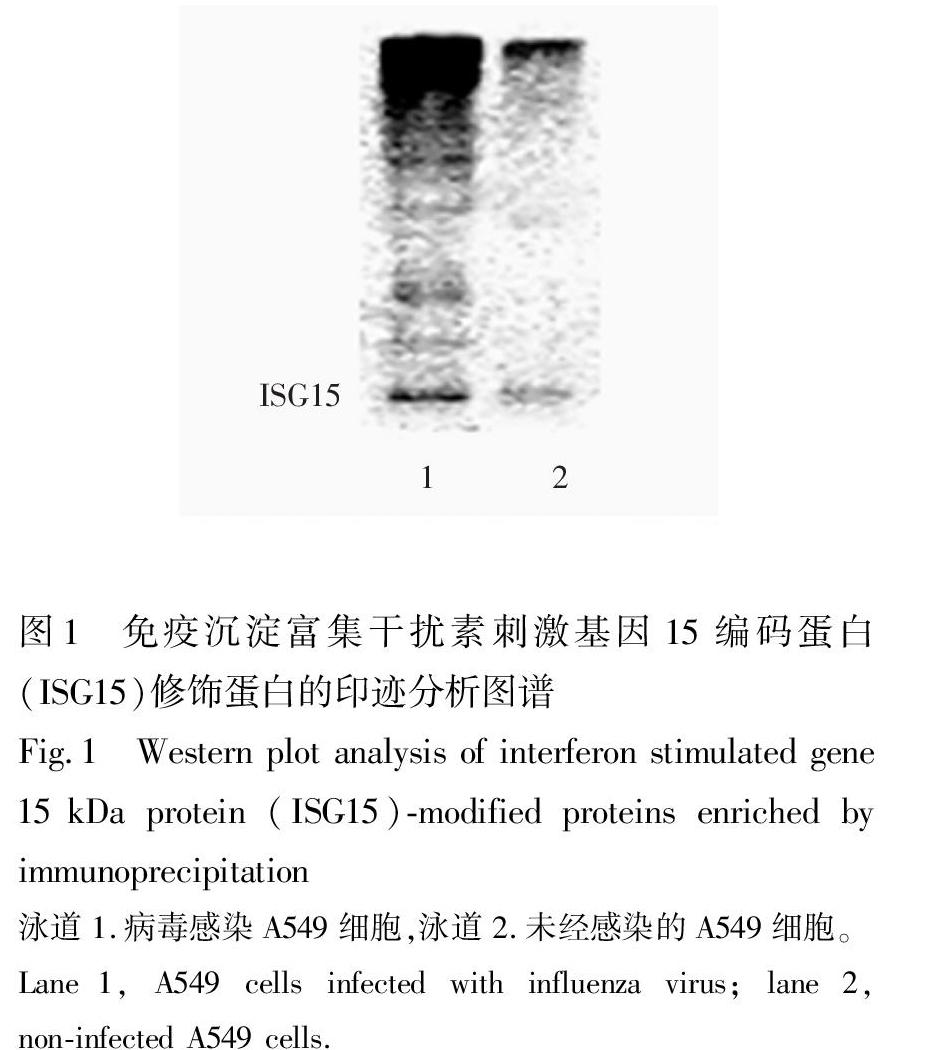
將H3N2病毒株接種于A549細胞,在不含血清的DMEM培養基中培養24 h,棄去培養液,以PBS緩沖液洗滌3次后,收集并裂解細胞。對照組為未接種病毒的A549細胞,培養條件與實驗組相同。共進行6次平行實驗。實驗組和對照組細胞裂解液分別采用抗ISG15抗體免疫沉淀富集、SDSPAGE電泳分離及轉膜,進行蛋白印跡分析(圖1)。為了減少非特異性吸附,在免疫沉淀富集之前,向細胞裂解液中加入未偶聯抗體的瓊脂糖微球進行預吸附,然后離心棄去瓊脂糖微球。再在上清液中加入偶聯ISG15抗體的瓊脂糖微球進行免疫沉淀實驗。實驗結果表明,未經病毒感染的A549細胞中也能檢測到極微量的ISG15以及ISG15修飾蛋白;而病毒感染細胞后,顯著誘導了ISG15及其修飾系統的表達,使得宿主細胞蛋白產生明顯的ISG15修飾,說明ISG15修飾在宿主細胞抵抗病毒的固有免疫反應中發揮了重要作用。
3.2ISG15修飾蛋白的液相色譜質譜聯用分析鑒定
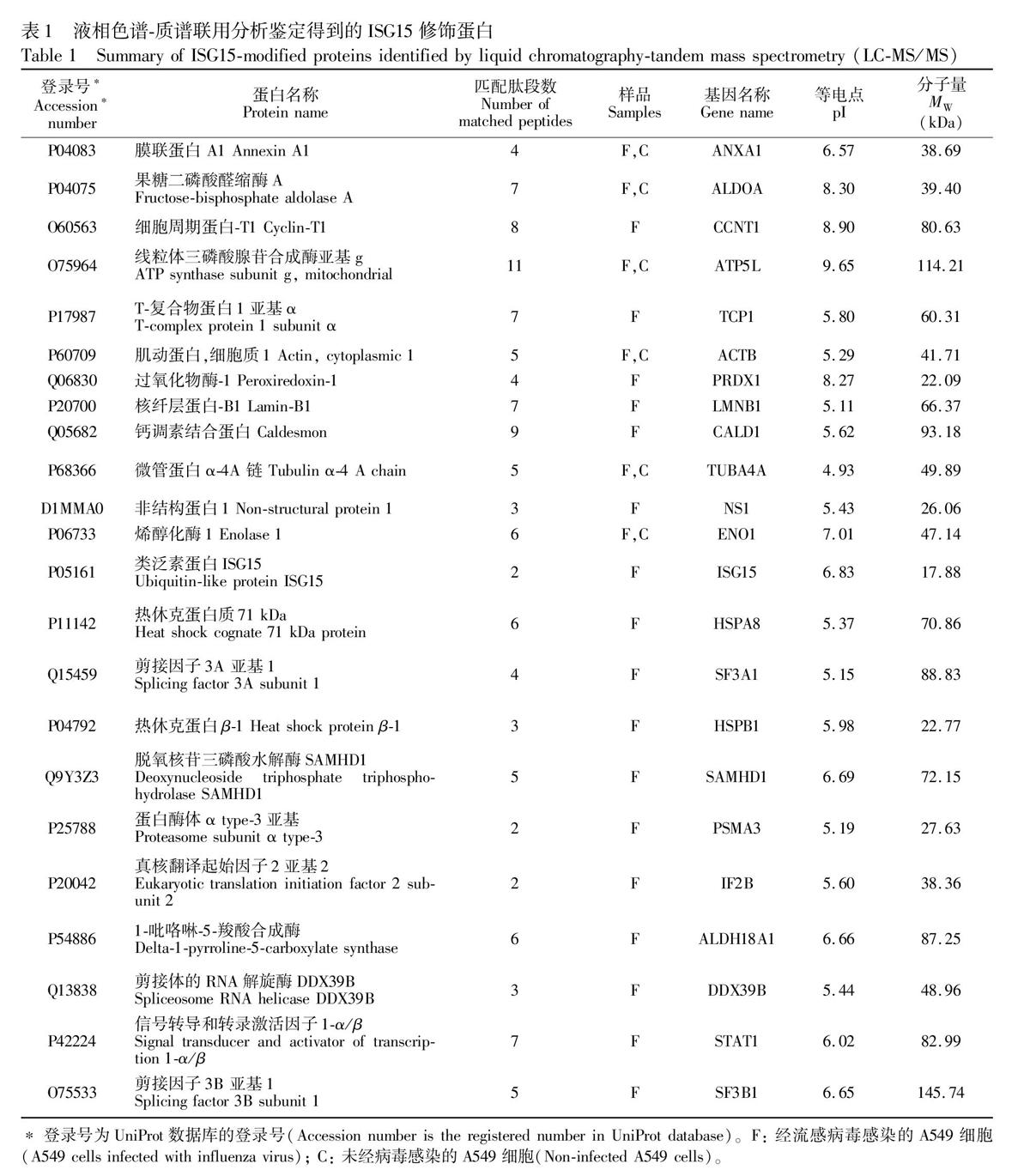
將經免疫沉淀富集的ISG15修飾蛋白進行二硫鍵還原、碘乙酰胺烷基化后,利用胰蛋白酶對蛋白質混合物進行特異性水解,得到酶解多肽的混合物。經液相色譜質譜聯用分析得到酶解多肽的質譜數據,再通過蛋白質數據庫檢索,在流感病毒感染的A549細胞中鑒定得到包括ISG15蛋白在內的23種蛋白(表1)。其中6種蛋白在未經感染的細胞中也得到表達,說明在細胞正常的代謝過程中,一部分細胞內源性蛋白也會發生ISG15修飾。文獻[24]表明,細胞經過病毒感染等外界刺激后,會顯著誘導表達ISG15。而ISG15對感染病毒的宿主細胞中的哪些蛋白進行修飾,尚未有系統的研究報道。流感病毒的非結構蛋白NS1是人源宿主細胞中類泛素ISG15修飾的一個重要靶蛋白[25,26],宿主細胞通過ISG15修飾抑制NS1的功能,從而抵抗病毒在機體內的有效復制,是病毒和宿主相互作用途徑之一。在本研究中,也同樣鑒定到來自流感病毒的非結構蛋白NS1。在流感病毒感染宿主細胞過程中,NS1蛋白是一個關鍵致病因子,它通過多種機制,包括雙鏈RNA的結合和隔離來拮抗宿主的抗病毒反應。
目前,采用免疫沉淀和免疫共沉淀技術將目標蛋白進行純化,再經液相色譜質譜聯用技術進行分析鑒定的方法,已廣泛應用于生命分析科學的多個領域 [27,28]。然而,采用這種技術對ISG15修飾蛋白的研究還鮮有報道。目前,關于ISG15修飾蛋白的研究,多先構建帶有某種標簽(如Flag, HA等)的ISG15表達載體,再轉染到合適的細胞中進行表達。將細胞裂解后,利用對標簽具有特異性的固相支持物對ISG15修飾的蛋白進行富集[29,30]。這種方法對修飾蛋白富集的特異性較高(源自于標簽的特異性)。然而由于方法本身的限制,多用于體外研究。而本研究所采用的直接對ISG15免疫富集的方法,可以更有效地分析識別內源性ISG15的修飾狀態,以及減少外源表達體系對生物學過程的干擾。因此,這種方法更適合體內研究體系以及基于臨床樣本的研究。
3.3ISG15修飾蛋白的生物信息學分析
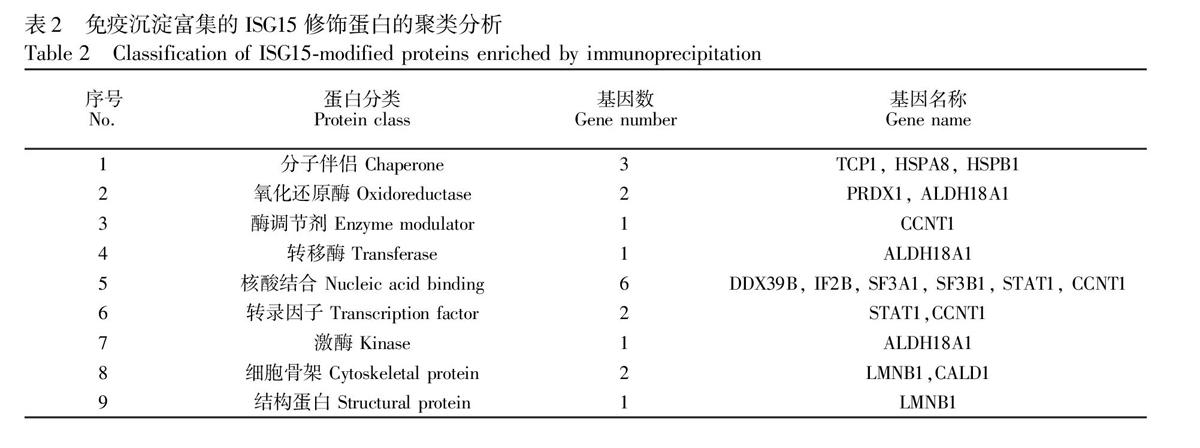
本研究得到的大部分ISG15修飾蛋白是經病毒感染后誘導產生ISG15修飾的,這些蛋白的修飾往往跟流感病毒與宿主細胞相互作用的分子機制密切相關。采用PANTHER軟件對這些蛋白(只在流感病毒感染的細胞中檢測到的ISG15修飾的宿主蛋白,共15種)進行了聚類分析,將具有相似生物學功能屬性的蛋白進行歸屬(表2)。結果表明,流感病毒感染引起的ISG15修飾蛋白具有較廣泛的功能分布,分別歸屬于9個不同的蛋白分類(Protein class),包括細胞骨架、分子伴侶、酶調節劑、核酸結合、激酶、轉移酶、轉錄因子、氧化還原酶以及結構蛋白。ISG15修飾作為一種重要的翻譯后修飾,在多種信號轉導途徑中具有十分重要的作用。研究表明,ISG15可以通過多種方式在流感病毒和宿主相互作用中發揮生物學功能。一方面,ISG15可以通過對宿主蛋白的修飾,增強宿主對病毒的抵抗作用。另一方面,ISG15可以對病毒蛋白進行修飾,導致其不能形成特定的高級結構,從而降低其對宿主抗病毒作用的抵抗能力。
[KH*4D][HT5”SS][HJ*4]表2免疫沉淀富集的ISG15修飾蛋白的聚類分析
Table 2Classification of ISG15modified proteins enriched by immunoprecipitation
[HT6SS][BG(][BHDFG4,WK8,WK18,WK8*2,WK22W]序號No.蛋白分類Protein class基因數Gene number基因名稱Gene name
1分子伴侶 Chaperone3TCP1, HSPA8, HSPB1
2氧化還原酶 Oxidoreductase
2PRDX1, ALDH18A13酶調節劑 Enzyme modulator1CCNT14
轉移酶 Transferase1ALDH18A15核酸結合 Nucleic acid binding 6DDX39B, IF2B, SF3A1, SF3B1, STAT1, CCNT1
6轉錄因子 Transcription factor 2STAT1,CCNT17激酶 Kinase 1ALDH18A1
8細胞骨架 Cytoskeletal protein 2LMNB1,CALD19結構蛋白 Structural protein 1LMNB1[BHDFG1*2,WKZQ0W][BG)W][HT5][HJ]
4結 論
本研究利用免疫沉淀技術將類泛素ISG15修飾的蛋白富集純化后,利用液相色譜質譜聯用技術對流感病毒感染A549宿主細胞過程中產生的類泛素ISG15修飾蛋白進行了分析鑒定。成功鑒定出了包括ISG15在內的22種來源于宿主細胞的蛋白和1種來源于流感病毒的非結構蛋白。研究結果提示類泛素ISG15在流感病毒和宿主的相互作用中發揮重要作用。本研究為大規模分析鑒定ISG15修飾蛋白提供了一種特異、有效的研究方法。
References
1Shapira S D, GatViks I, Shum B O, Dricot A, de Grace M M, Wu L, Gupta P B, Hao T, Silver S J, Root D E, Hill D E, Regev A, Hacohen N. Cell, 2009, 139(7): 1255-1267
2Schmitz M L, Kracht M, Saul V V. Biochim. Biophys. Acta, 2014, 1843(11): 2754-2764
3Buggele W A, Johnson K E, Horvath C M. J. Biol. Chem., 2012, 287(37): 31027-31040
4Zhao X, Dai J, Xiao X, Wu L, Zeng J, Sheng J, Su J, Chen X, Wang G, Li K. PLoS One, 2014, 9(8): e104506
5Isaacson M K, Ploegh H L. Cell Host Microbe, 2009, 5(6): 559-570
6Harty R N, Pitha P M, Okumura A. J. Innate Immun., 2009, 1(5): 397-404
7Zhao C, Collins M N, Hsiang T Y, Krug R M. Trends Microbiol., 2013, 21(4): 181-186
8Sgorbissa A1, Brancolini C. Cytokine Growth Factor Rev., 2012, 23(6): 307-314
9Herrmann J, Lerman L O, Lerman A. Circ. Res., 2007, 100(9): 1276-1291
10Lenschow D J, Lai C, FriasStaheli N, Giannakopoulos N V, Lutz A, Wolff T, Osiak A, Levine B, Schmidt R E, GarcíaSastre A, Leib D A, Pekosz A, Knobeloch K P, Horak I, Virgin H W 4th. Proc. Natl. Acad. Sci. USA, 2007, 104(4): 1371-1376
11Hsiang T Y, Zhao C, Krug R M. J. Virol., 2009, 83(12): 5971-5977
12ZHANG Wei. Chinese J. Anal.Chem., 2014, 42(12): 1859-1868
張 偉. 分析化學, 2014, 42(12): 1859-1868
13Liu N, Song W, Wang P, Lee K, Chan W, Chen H, Cai Z. Proteomics, 2008, 8(9): 1851-1858
14Sun N, Sun W, Li S, Yang J, Yang L, Quan G, Gao X, Wang Z, Cheng X, Li Z, Peng Q, Liu N. Int. J. Mol. Sci., 2015, 16(11): 25982-25998
15LIN Lin, LUO ShuSheng, WANG LingJue, YANG Jie, SHENG HaiNan, TIAN RuiJun, Chinese J. Anal. Chem., 2015, 43(10): 1479-1489
林 琳, 羅樹生, 王靈玨, 楊 杰, 沈海南, 田瑞軍. 分析化學, 2015, 43(10): 1479-1489
16LI ZiJian, HE Xin, PAN ChenYu, LIU Ning. Chinese J. Anal. Chem., 2013, 41(11): 1653-1658
李子健, 何 鑫, 潘晨宇, 劉 寧. 分析化學, 2013, 41(11): 1653-1658
17Liu N, Wang G, Lee KC, Guan Y, Chen H, Cai Z. FASEB J., 2009, 23(10): 3377-3382
18Lewicki M C, Srikumar T, Johnson E, Raught B. J. Proteomics, 2015, 118: 39-48
19Thomas S N, Zhang H, Cotter R J. Clin. Proteomics, 2015, 12(1): 14
20Ordureau A, Münch C, Harper J W. Mol. Cell, 2015, 58(4): 660-676
21Giannakopoulos N V, Luo J K, Papov V, Zou W, Lenschow D J, Jacobs B S, Borden E C, Li J, Virgin H W, Zhang D E. Biochem. Biophys. Res. Commun., 2005, 336(2): 496-506
22Mi H, Muruganujan A, Casagrande J T, Thomas P D. Nat. Protoc., 2013, 8(8): 1551-1566
23SUN NingNing, WANG ZiJian, GAO Xiang, CHENG Xin, SUN WanChun, YANG JingBo, LIU Ning. Chinese J. Anal. Chem., 2015, 43(6): 808-813
孫寧寧, 王子健, 高 祥, 程 欣, 孫萬春, 楊靜波, 劉 寧. 分析化學, 2015, 43(6): 808-813
24Liu L, Zhou J, Wang Y, Mason R J, Funk C J, Du Y. J. Proteome Res., 2012, 11(8): 4091-4101
25Zhao C, Hsiang T Y, Kuo R L, Krug R M. Proc. Natl. Acad. Sci. USA, 2010, 107(5): 2253-2258
26Tang Y, Zhong G, Zhu L, Liu X, Shan Y, Feng H, Bu Z, Chen H, Wang C. J. Immunol., 2010, 184(10): 5777-5790
27Ning Z, Hawley B, Chiang C K, Seebun D, Figeys D. Methods Mol. Biol., 2014, 1164: 1-13
28Li K W, Chen N, Klemmer P, Koopmans F, Karupothula R, Smit A B. Proteomics, 2012, 12(1516): 2428-2432
29Zhao C, Denison C, Huibregtse J M, Gygi S, Krug R M. Proc. Natl. Acad. Sci. USA, 2005, 102(29): 10200-10205
30Takeuchi T, Yokosawa H. Methods Mol. Biol., 2008, 446: 139-149
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
