γ-氨基丁酸產生菌的分離及發酵條件優化
2016-10-26 09:34:13汪祥燕徐海燕辛國芹汪孟娟董佩佩趙影谷巍
中國飼料 2016年4期
關鍵詞:產量
汪祥燕,徐海燕,辛國芹,汪孟娟,董佩佩,趙影,谷巍
(山東寶來利來生物工程股份有限公司,山東泰安271000)
?
添加劑
γ-氨基丁酸產生菌的分離及發酵條件優化
汪祥燕*,徐海燕,辛國芹,汪孟娟,董佩佩,趙影,谷巍
(山東寶來利來生物工程股份有限公司,山東泰安271000)
γ-氨基丁酸(GABA)是哺乳動物體內的一種抑制性神經遞質,具有許多重要的生理功能。利用酸菜、酸奶等食品進行乳酸菌的分離,通過紙層析法篩選得到1株產GABA量較高的菌株Y-3,經16SrDNA鑒定該菌株為短乳桿菌。單因素試驗和正交試驗確定的菌株Y-3最佳發酵條件為:葡萄糖1.5%、酵母膏0.5%、胰蛋白胨1.5%、L-谷氨酸鈉1.0%、丁二酸鈉0.5%,初始pH為6.0,接種量6%,37℃靜置培養48 h,GABA產量達12 g/L以上,比優化前產量提高了近5倍。
γ-氨基丁酸;分離;短乳桿菌;發酵
γ-氨基丁酸(GABA)是一種天然存在的非蛋白氨基酸,屬強神經抑制性氨基酸,具有鎮靜、催眠、抗驚厥、降血壓的作用(Li等,2010)。應用于畜牧生產中能有效地提高動物抗應激能力和抗缺氧能力,改善動物生產性能和動物產品品質(譚良溪等,2012;吳俊鋒和李呂木,2011)。通過減少動物的無意識運動以減少能量消耗,達到促生長的目的(吳凡和譚青松,2015;徐秋良等,2009)。GABA廣泛存在于植物、動物、微生物中,合成制備方法主要有化學合成、植物富集和微生物發酵3種(Dhakal等,2012)。目前GABA的生產主要采用發酵法,而安全性高的乳酸菌可以利用體內的谷氨酸脫羧酶,將L-谷氨酸鈉經α-脫羧后產生GABA,開發前景廣闊。陸小雪等(2009)從泡菜和酸奶中分離出3株產GABA的乳酸菌,分別為乳酸乳球菌乳酸亞種、乳酸乳球菌乳酸亞種和唾液鏈球菌嗜熱亞種,其發酵液中GABA含量分別達到3.680、3.341、2.700 g/L。蔣冬花等(2007)分離出相對高產GABA的植物乳桿菌,在最佳發酵條件下發酵液中GABA含量可達4 g/L。本試驗從泡菜、酸奶等食品中分離高產GABA菌株,對其進行分子生物學鑒定,并優化菌株的發酵條件,為微生物發酵富集GABA的研究及相關產品的開發提供理論依據。
1 材料與方法
1.1試驗材料泡菜、酸菜及各品牌的酸奶(伊利、蒙牛、亞奧特)等。
1.2培養基
1.2.1分離培養基胰蛋白胨10 g/L、酵母膏5 g/L、葡萄糖5 g/L、吐溫-80 1 g/L、K2HPO42 g/L、醋酸鈉5 g/L、檸檬酸二胺2 g/L、MgSO4·7H2O 0.2 g/L、MnSO4·H2O 0.05 g/L、pH 6.5~6.8,37℃培養48 h。
1.2.2種子培養基K2HPO42 g/L,檸檬酸三銨2 g/L,無水乙酸鈉5 g/L,MnSO4·H2O 0.25 g/L,MgSO4·7H2O 0.58 g/L,葡萄糖20 g/L,蛋白胨10 g/L,牛肉膏10 g/L,酵母膏5 g/L。
1.2.3TYG發酵培養基胰蛋白胨5 g/L,酵母膏5 g/L,葡萄糖10 g/L,丁二酸鈉5 g/L,L-谷氨酸鈉10 g/L,pH 6.6~6.8。
1.3試驗方法
1.3.1乳酸菌的分離分別取泡菜汁、酸菜汁及伊利、蒙牛、亞奧特酸奶各1 mL,接種于50 mL分離培養基中富集培養3 d,取富集培養液各1 mL,分別稀釋至10-1、10-2、10-3、10-4、10-5濃度,將所有樣品稀釋液各取0.3 mL,倒入已滅菌的平皿中,然后倒入冷卻至50℃左右的分離培養基混勻,凝固后置30℃培養箱中培養3 d,挑取帶有溶鈣圈的菌落,接種于MRS斜面培養基。
1.3.2乳酸菌的篩選將初篩得到的菌株轉接于液體MRS種子培養基中,37℃靜置培養12~15 h,然后以2%的接種量接入到含1%L-谷氨酸鈉的TYG發酵培養基中,37℃靜置培養24 h。將發酵液離心后進行檢測分析。
1.3.3分析方法
1.3.3.1γ-氨基丁酸含量的測定GABA含量的測定采用紙層析-比色法。(1)展開:采用新華3號濾紙,每個樣品點樣4 μL,在室溫下用不飽和法展開,展開劑用(苯酚∶水=4∶1)上行法展開;(2)定量:配制0.5%的GABA標準品,點樣4 μL;菌株發酵液經離心后,取上清液點樣4 μL,放入層析缸中展開;(3)顯色:展開結束后自然晾干,用0.5%的茚三酮無水乙醇溶液噴淋,在65℃顯色30 min;(4)比色測定:剪下GABA顯色條帶,用5 mL硫酸銅乙醇溶液(0.1%硫酸銅溶液∶75%乙醇=2∶38)浸泡30 min,然后測定520 nm波長下的吸光值(A)。根據標準品的A值和樣品的A值即可計算出樣品的GABA含量。
1.3.3.2菌株生長量的測定發酵培養過程中,在一定時間取培養液,以空白培養基作對照,于600 nm波長下測定吸光值(OD600)。
1.3.3.3pH的測定采用PHS-3C型精密pH計測定。
1.3.4發酵條件優化采用單因素試驗和正交試驗優化發酵條件。
1.3.5菌株的鑒定
1.3.5.116SrDNA擴增與序列分析目的菌株接種于新鮮的種子培養基中培養12 h,采用天根公司的試劑盒提取菌體DNA,并對其進行16S rDNA序列擴增。所用引物為通用引物:1492r 5'-GGTTACCTTGTTACGACTT-3'27f 5'-AGAGTTG ATCCTGGCTCAG-3';PCR反應體系(50 μL)為:Mixture 25 μL(含Taq DNA聚合酶及dNTP等,天根生化科技有限公司),上下游引物各1 μL,模板DNA 2 μL,超純水21 μL。PCR擴增程序為94℃預變性5 min,94℃變性1 min,52℃退火1 min,72℃延伸2 min,30個循環,72℃延伸10 min。PCR產物送北京博尚生物技術有限公司進行序列測定。
1.3.5.2系統發育分析登錄GenBank(http:// www.ncbi.nlm.nih.gov),利用Blast對菌株16SrDNA測序結果進行檢索,并下載相關屬種的16SrDNA序列,采用DNAMAN、DNAclub、MEGA5.2等軟件進行同源性分析,并構建系統進化樹。
2 結果與分析
2.1菌株篩選樣品經處理后,利用乳酸菌選擇培養基從各分離源中共分離純化出113株乳酸菌,并編號保存。采用紙層析-比色法初篩,得到8株能夠利用L-谷氨酸鈉轉化生成γ-氨基丁酸的菌株。
由表1可知,不同菌株產γ-氨基丁酸的能力有一定差別,僅2株菌的GABA產量高于1 g/L,其中Y-3在未經培養優化的條件下,可產生GABA 2.21 g/L。將Y-3菌株傳接10代,分別測定Y-3菌株1~10代發酵后GABA產量,發現其GABA產量穩定,為(2.36±0.22)g/L,后續對菌株Y-3進行發酵條件優化。

表1 菌株篩選結果
2.2菌株Y-3發酵條件的優化
2.2.1接種量對發酵生產γ-氨基丁酸的影響由圖1看出,接種量對Y-3菌株發酵生產GABA有較大的影響。接種量為2%~4%時,GABA產量在2.20 g/L左右;接種量增大至6%后,GABA產量明顯提高,達4.46 g/L,提高2倍;接種量大于6%后,GABA產量降低,反而不利于菌株生產GA-BA。所以,菌株Y-3最優接種量確定為6%。

圖1 接種量對Y-3菌株GABA產量的影響
2.2.2pH對發酵生產γ-氨基丁酸的影響將發酵培養基起始pH分別調為4.0、5.0、6.0和自然pH(6.70左右),研究發酵培養基起始pH對Y-3菌株生長、pH及GABA產量的影響,結果見圖2。隨著發酵培養基起始pH增大,Y-3菌株的菌體量和發酵液pH逐漸增大。發酵培養基起始pH為4.0時,發酵結束時菌體密度僅有1.084;而在起始pH為6.0時,發酵結束時菌體密度為2.109,兩者相比,菌體密度增大了1倍。隨著發酵培養基起始pH升高,GABA產量為先升高后降低,在pH=6.0時達最大,為3.10 g/L。在pH=6.0時菌體密度也較大,菌體量大代表有更多菌體參與GABA生產,因而GABA產量才會提高。雖然自然pH時(6.70左右)菌體生長量大于pH為6.0時,但是此時的發酵液pH較低,降低了谷氨酸脫羧酶的活性,GABA產量反而沒有pH=6.0時高。綜合分析,發酵培養基起始pH確定為6.0。

圖2 發酵培養基起始pH對Y-3菌株生長、pH及GABA產量的影響
2.2.3發酵時間對發酵生產γ-氨基丁酸的影響發酵時間對于菌株Y-3和PS-9生產GABA有很大的影響,結果見圖3。隨著發酵時間延長,菌株Y-3的菌體密度逐漸增大,24 h達到最大,為2.109,之后處于基本穩定狀態。發酵液pH在 24 h時最低,僅為2.94,然后升高,之后處于基本穩定。分析原因可能是:24 h前菌株增殖速度快,處于菌體積累階段,菌株會迅速消耗葡萄糖,將其分解為小分子酸,使得發酵液pH降低,之后菌株Y-3會利用H+,在谷氨酸脫羧酶作用下將L-谷氨酸鈉轉化為GABA,又提高了發酵液的pH。且在一定范圍內,GABA產量是隨著發酵時間延長逐漸增大的,在48 h時達到最大,為3.57 g/L。此后,GABA產量有下降但不明顯。從培養周期和成本等工業擴大生產因素考慮,培養時間不宜過長,確定菌株Y-3最佳發酵時間為48 h。

圖3 發酵時間對Y-3菌株生長、pH及GABA產量的影響
2.3菌株Y-3發酵培養基的優化
2.3.1碳源對發酵生產γ-氨基丁酸的影響以TYG發酵培養基為基礎,分別選取葡萄糖、蔗糖、麥芽糖、可溶性淀粉及丁二酸鈉為唯一碳源,添加量為10 g/L,發酵培養48 h后檢測發酵液的GABA含量,結果見圖4。結果顯示,對于菌株Y-3,葡萄糖作為發酵培養基的碳源最為適合,麥芽糖、可溶性淀粉和丁二酸鈉次之,蔗糖最差。葡萄糖作為可快速利用的碳源為菌株Y-3的生長和GABA生產提供能量來源,故以其作碳源,無論是菌株生長,還是GABA產量都優于其他碳源。

圖4 碳源對Y-3菌株生長和GABA產量的影響
2.3.2氮源對發酵生產γ-氨基丁酸的影響以TYG發酵培養基為基礎,分別選取胰蛋白胨、牛肉膏、酵母膏、蛋白胨、玉米漿、硫酸銨及硝酸鈉為唯一氮源,添加量為10 g/L,發酵培養48 h檢測發酵液的GABA含量。由圖5可知,對于菌株Y-3,有機氮效果比無機氮效果好。硫酸銨、硝酸鈉兩種無機氮源發酵效果最差,紙層析圖上基本看不出條帶。酵母膏作為單一氮源發酵效果最好,GABA最高為5.95 g/L,其次是牛肉膏、蛋白胨和胰蛋白胨,玉米漿的效果較差。酵母膏中含有多種成分,相對于其他氮源,酵母膏的營養成分可能更有利于乳酸菌的生長,且其可能含有刺激谷氨酸脫羧酶活性的因子,因此GABA產量較高。

圖5 氮源對Y-3菌株生長和GABA產量的影響
進一步,以TYG發酵培養基為基礎,在以酵母膏作氮源的基礎上分別復合蛋白胨、牛肉膏和胰蛋白胨為復合氮源,發酵培養48 h,以紙層析-比色法測定上清中GABA含量。由圖6可知,對于菌株Y-3,以酵母膏和胰蛋白胨復合時發酵效果最好,GABA的產量達6.26 g/L,比酵母膏作唯一氮源時產量提高5%。所以,菌株發酵產GABA試驗選擇酵母膏復合胰蛋白胨作氮源。

圖6 復合氮源對Y-3菌株生長和GABA產量的影響
2.3.3正交試驗設計優化發酵培養基以葡萄糖為碳源,酵母膏和胰蛋白胨為復合氮源,添加L-谷氨酸鈉,進行L9(34)正交試驗設計。正交試驗設計及結果分析見表2。
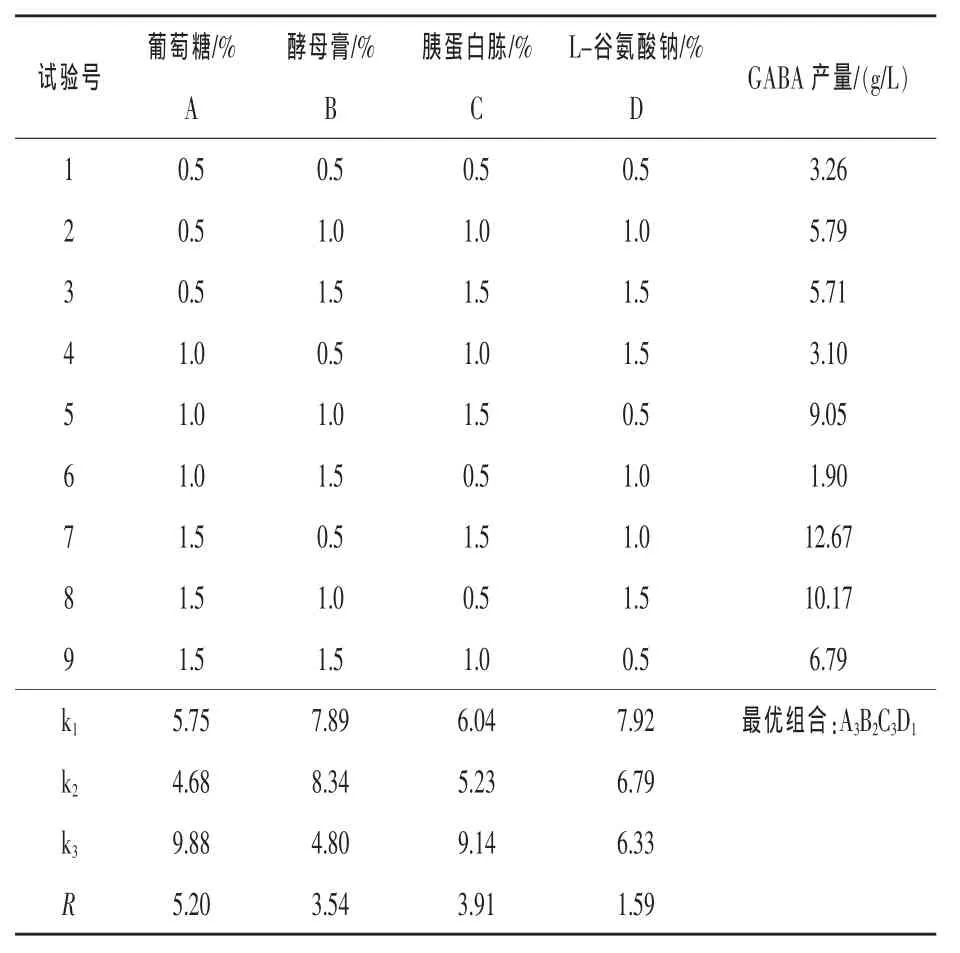
表2 L9(34)正交試驗設計及結果分析
由表2可知,葡萄糖添加量對于GABA的產量影響最大。各成分對GABA產量的影響順序為:葡萄糖>胰蛋白胨>酵母膏>L-谷氨酸鈉,極差分析的最優組合為A3B2C3D1。試驗中最優組合為A3B1C3D2,GABA產量為12.67 g/L。由于最佳的培養基組成在試驗中沒有進行,所以根據正交試驗給出的最佳培養基成分進行發酵,產量與7號試驗相比沒有明顯提高。考慮成本等因素,確定菌株Y-3發酵配方為:葡萄糖1.5%、酵母膏0.5%、胰蛋白胨1.5%、L-谷氨酸鈉1.0%、丁二酸鈉0.5%,pH 6.0。
2.4發酵液處理及凍干粉γ-氨基丁酸含量測定按照優化結果發酵培養菌株Y-3,在發酵結束后將發酵液凍干處理,計算100 mL發酵液的凍干粉比例,并以紙層析-比色法測定凍干粉中GABA的含量。結果顯示,100 mL發酵液凍干后可得到3.72 g凍干粉,凍干粉中GABA含量為4.6%左右。
2.5菌株Y-3的鑒定
2.5.1菌落及菌體形態該菌株在分離培養基上培養24 h,其菌落形態扁平,白色不透明,邊緣光滑,表面濕潤,顯微鏡下觀察菌體為短桿狀,大多成對出現,革蘭氏染色陽性。
2.5.216SrDNA序列和系統發育分析該菌株的PCR產物電泳圖見圖7,條帶在1000~2000 bp處,約為1500 bp,與預期結果一致。

圖7 菌株Y-3擴增產物瓊脂凝膠電泳圖
將菌株Y-3的16SrDNA序列在NCBI中進行BLAST比對分析,結果顯示菌株Y-3與Lactobacillus brevis的序列同源性達到100%。鑒定該菌株屬于短乳桿菌(Lactobacillus brevis)。
從Genebank中調取并下載與菌株Y-3同源性較高的該屬內其他相關菌株的16S rDNA,利用MEGA5.2軟件的Neighboring-joining方法,構建系統進化樹(圖8)。
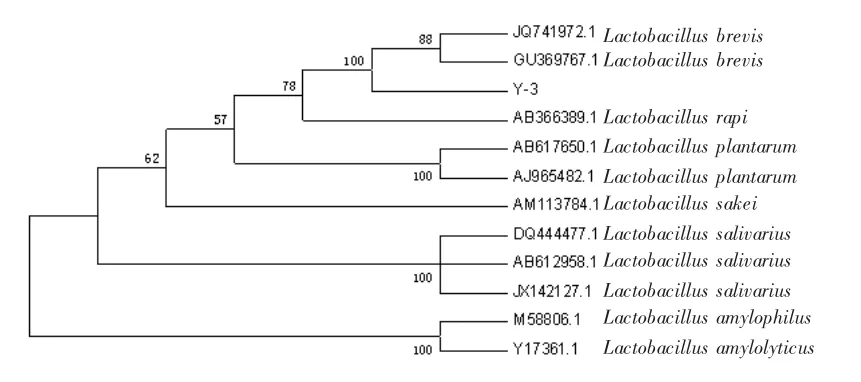
圖8 菌株Y-3與相關菌株的系統進化關系
3 小結
從泡菜、酸菜及不同品牌酸奶等分離源中分離純化得到113株乳酸菌,并以TYG培養基為基礎發酵,篩選到8株能產GABA的菌株。通過單因素試驗和正交試驗對培養基和發酵條件進行優化,確定菌株Y-3最佳培養基組合和發酵條件為:葡萄糖1.5%、酵母膏0.5%、胰蛋白胨1.5%、L-谷氨酸鈉1.0%、丁二酸鈉0.5%,初始pH為6.0,接種量6%,37℃靜置培養48 h,此時發酵液中GABA含量達12 g/L以上,比優化前產量增加了近5倍。對發酵液進行凍干處理,Y-3菌株100 mL發酵液凍干后可得4 g左右凍干粉,凍干粉中GABA含量為4.6%左右。通過16S rDNA鑒定,菌株Y-3為一株短乳桿菌。
[1]蔣冬花,后加衡,黃大年,等.酸菜中高產γ-氨基丁酸乳酸菌的篩選[J].微生物學雜志,2007,27(1):35~39.
[2]陸小雪,解春艷,顧振新.產γ-氨基丁酸乳酸菌的分離與鑒定[J].南京農業大學學報,2009,32(1):121~125.
[3]譚良溪,張銘,張復生.γ-氨基丁酸及其在畜禽生產中的研究進展[J].中國畜牧獸醫,2012,39(8):132~135.
[4]吳凡,譚青松.γ-氨基丁酸在動物營養中的研究進展[J].飼料研究,2015,2:61~65.
[5]吳俊鋒,李呂木.飼用γ-氨基丁酸研究進展[J].中國飼料,2011,16:28~30.
[6]徐秋良,張慶莉,唐國盤,等.γ氨基丁酸對高溫環境下生長肥育豬生產性能及激素水平的影響[J].家畜生態學報,2009,6:28~32.
[7]Dhakal R,Bajpai V K,Baek K H.Production of GABA(γ-aminobutyric acid)by microorganisms:A review[J].Brazilian Journal of Microbiology,2012,43(4):1230~1241.
[8]Li H,Qiu T,Huang G D,et al.Production of gamma-aminobutyric acid by Lactobacillus brevis NCL912 using fed-batchfermentation[J].Microbial Cell Factories,2010,9(9):85.
γ-amino butyric acid(GABA)as a putative inhibitory in the vertebrate nervous system,has many important physiological functions.Using filter paper chromatography method,a lacto-bacteria Y-3 highly yielding GABA was isolated from different foods such as pickled vegetable and yoghourt.Y-3 was identified as Lactobacillus brevis by its cultural characteristics,morphological characteristics and 16S rDNA sequence.The optimal fermentation medium composition by single factor and orthogonal test showed as follows:glucose 1.5%,yeast extract 0.5%,tryptone 1.5%,L-sodium glutamate 1.0%,sodium succinate 0.5%.The better fermentation conditions were as follows:initial pH 6.0,inoculation amount 6%,temperature 37℃,fermentation 48 h.Using this optimization conditions,the GABA yield was over 12 g/L,which increased nearly 5 times than that before optimization.
γ-amino butyric acid;isolation;Lactobacillus brevis;fermentation
S816.7
A
1004-3314(2016)04-0027-05
10.15906/j.cnki.cn11-2975/s.20160407
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(2021年10期)2021-12-05 16:31:48
今日農業(2021年14期)2021-11-25 23:57:29
今日農業(2021年13期)2021-08-14 01:37:56
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中國果業信息(2019年10期)2019-11-13 01:21:34
中國化肥信息(2019年2期)2019-01-18 15:24:35
中國化肥信息(2019年1期)2019-01-17 21:31:12
中國化肥信息(2019年4期)2019-01-17 18:47:06
