番茄質膜H+-ATPase家族基因的鑒定和表達分析
2016-10-27 09:49:44劉健健劉俊麗季敏杰陳家棟楊曉峰陳愛群
浙江農林大學學報 2016年5期
劉健健,劉俊麗,季敏杰,陳家棟,楊曉峰,陳愛群
(南京農業大學 資源與環境科學學院,江蘇 南京210095)
番茄質膜H+-ATPase家族基因的鑒定和表達分析
劉健健,劉俊麗,季敏杰,陳家棟,楊曉峰,陳愛群
(南京農業大學 資源與環境科學學院,江蘇 南京210095)
植物質膜H+-ATPase(Ec3.6.1.3)是一類普遍存在于細胞質膜上通過水解三磷酸腺苷(ATP)產生能量,將細胞質中的氫離子(H+)逆濃度泵出細胞的運輸蛋白。植物中的質膜H+-ATPase由一個多基因家族所編碼,其功能涉及到植物生長發育的多個生理過程。通過對全基因組檢索在茄科Solanaceae植物番茄Solanum lycopersicum中共鑒定到8個編碼質膜H+-ATPase的同源基因(LHA1~8)。生物信息學分析顯示:這8個LHA基因具有較高的序列相似性和較為保守的外顯子/內含子結構特征。實時熒光定量聚合酶鏈式反應(qRT-PCR)分析顯示,LHA1~4在所有被檢測的組織器官中都有表達,LHA5~7幾乎只在花器官中高量表達,而LHA8在正常培養和養分(氮、磷、鉀和鎂)缺乏以及高鹽脅迫處理條件下幾乎都不表達,但能夠在被菌根真菌侵染的根系中強烈表達。將一段2 669 bp的LHA8的啟動子融合GUS報告基因轉入到煙草Nicotiana tabacum中發現,GUS基因幾乎只在被菌根真菌菌絲侵入形成叢枝的根系細胞中特異性表達。圖5表2參22
分子生物學;番茄;質膜H+-ATPase;表達模式;菌根真菌
H+-ATPase是廣泛存在于植物質膜和各種內膜系統中的一種功能蛋白,在細胞代謝過程中起非常關鍵的作用[1]。目前,被報道的H+-ATPase按結構可分為P型、F型和V型。其中F型H+-ATPase位于葉綠體內囊體膜和線粒體內膜上,V型H+-ATPase定位于液泡膜上,質膜H+-ATPase屬于P型ATPase[2]。質膜H+-ATPase能夠通過水解三磷酸腺苷(ATP)產生能量,將細胞質中的氫離子(H+)逆濃度泵出細胞(也稱為質膜質子泵),其功能主要是在細胞膜兩側產生氫離子(H+)濃度梯度和膜電位,為一系列次級轉運體和通道蛋白跨質膜轉運各種離子和小分子代謝產物提供驅動力和能量[3]。一系列的實驗結果證明:植物質膜H+-ATPase能夠參與胞內酸堿度的調節、離子平衡、細胞的伸長生長、氣孔的開閉等多種生理過程[4]。植物中的質膜H+-ATPase幾乎都是由一個多基因家族所編碼,且不同的成員在表達上具有一定的特異性和部分重疊性[5]。例如,在模式植物擬南芥Arabidopsis thaliana中共鑒定到11個編碼質膜H+-ATPase的同源基因(AHA1~11)[6]。其中,AHA1和AHA2在所有的組織器官中都有表達,其表達模式趨向于組成型[7];AHA3主要在維管組織和生殖器官中表達[8];AHA6,AHA8和AHA9幾乎只在花器官中表達[9];AHA10主要在發育中的種子種皮的內膜上表達[10]。這些結果暗示了在長期的進化過程中,不同的質膜H+-ATPase基因在植物發育的不同階段所分化形成的相對保守性和/或特異性的生理功能。植物質膜H+-ATPase活性在基因表達水平的調節還受到激素(如吲哚-3-乙酸等)和環境(如鹽害、病菌侵染、菌根真菌共生)因素的影響[11]。番茄 Solanum lycopersicum作為茄科Solanaceae作物中的一員,不僅是一種重要的經濟作物,而且由于其基因組相對比較簡單(大部分品種都為二倍體),以及全基因組測序和組裝工作已經接近完成,也已經成為生物學和遺傳學研究的一個理想的模式作物[12]。根據前期研究關于Southern印記雜交的結果推測,番茄中可能存在著至少7個編碼質膜H+-ATPase的基因LHA1~7[13]。現有研究僅對其中的3個基因(LHA1,LHA2和LHA4)進行了全長cDNA序列的克隆和表達調控分析,而其他4個可能的LHA基因只報道了約200 bp的DNA序列[14]。本研究通過對番茄全基因組序列文庫和EST/cDNA文庫進行檢索,共獲得了8個具有完整編碼區的質膜H+-ATPase基因LHA1~8,其中LHA8為從未報道過的新基因。對這8個基因的序列結構特征、進化關系以及組織表達模式做進一步分析,可為將來深入研究番茄質膜H+-ATPase基因家族的生物學功能奠定工作基礎。
1 材料與方法
1.1供試材料
番茄微型模式品種Solanum lycopersicum‘Micro-Tom’,成熟期為70~80 d,株高約為15 cm。
1.2試驗設計
番茄種子用體積分數為10%的過氧化氫溶液表面消毒10 min,自來水沖洗后置滅過菌的石英沙于25℃培養箱中萌發至真葉完全展開,用1/2濃度的營養液和完全濃度的營養液各培養1周后移栽進行后續實驗處理。營養液的配方及濃度為:1.00 mmol·L-1硝酸銨(NH4NO3),2.00 mmol·L-1硝酸鉀(KNO3),0.50 mmol·L-1硝酸鈣(Ca(NO3)2),1.00 mmol·L-1磷酸二氫鈉(NaH2PO4),0.25 mmol·L-1氯化鈣(CaCl2),0.50 mmol·L-1硫酸鎂(MgSO4),20.00 μmol·L-1乙二胺四乙酸鐵鈉(Fe-EDTA),9.00 μmol·L-1氯化錳(MnCl2),46.00 μmol·L-1硼酸(H3BO3),8.00 μmol·L-1硫酸鋅(ZnSO4),3.00 μmol·L-1硫酸銅(CuSO4),0.03 μmol·L-1鉬酸銨((NH4)6Mo7O24)。pH值調至pH 5.8。
養分缺乏和高鹽處理:試驗共設計4個養分缺乏(氮、磷、鉀和鎂)和1個高鹽(氯化鈉)脅迫處理。對照處理為完全營養液培養;養分缺乏處理中的磷、鉀和鎂的濃度分別降低到對照處理的1/20,其他養分元素濃度不變;高鹽處理為在完全營養液中額外加入200 mmol·L-1的氯化鈉。生物學重復3個·處理-1。
接種菌根真菌盆栽試驗:試驗前將洗凈的石英砂滅菌后裝到3 L的塑料盆中,種番茄小苗3株·盆-1,苗根部接入2.0 g·株-1左右的叢枝菌根真菌Rhizophagus irregularis孢子菌劑,共設3個生物學重復。
1.3實時熒光定量聚合酶鏈式反應(qRT-PCR)分析
取約2 μg的植物組織樣品的總RNA,嚴格按照Takara公司的PrimeScript RT reagent Kit With cD-NA Eraser反轉錄試劑盒的操作說明進行cDNA合成及后續的實時熒光定量PCR分析(ABI plus real-time PCR system。表1)。內參基因為組成型表達的Actin基因[15]。

表1 番茄8個LHA基因及Actin的定量引物Table 1 Gene-specific primers used for real-time RT-PCR amplification of tomato LHA and Actin genes
1.4進化樹分析
HA蛋白序列的比對分析和系統進化樹的繪制分別由ClustalX 1.83和MEGA 4.0軟件完成。構建進化樹的算法采用鄰接法(Neighbor-Joining)。
1.5啟動子片段融合pBI121表達載體的構建
根據LHA8起始密碼子ATG上游的啟動子序列,設計5′端分別引入HindⅢ和BamHⅠ的正向引物(aagcttagaatccatcattggatcact)和反向引物(ggatccggtagctcaattgattgaaccc),以番茄基因組DNA為模板,進行PCR擴增,將擴增得到的2 669 bp的啟動子片段克隆到pEASY-Blunt載體(北京全式金公司產品)。測序驗證后用HindⅢ和BamHⅠ將啟動子片段從克隆載體中切下,回收。同樣將雙元表達載體pBI121用HindⅢ和BamHⅠ將35S啟動子片段切下并回收剩余的線性化的片段,與從克隆載體上切下的目的基因啟動子片段通過 T4 DNA Ligase定向連接。將重組后的表達載體質粒用電擊法轉化至根癌農桿菌A-grobacterium tumefaciens感受態細胞(EHA105)中備用。
1.6煙草Nicotiana tabacum轉基因
采用根癌農桿菌介導的煙草遺傳轉化方法(葉盤法)[16]。
1.7GUS染色
剪取不同轉基因株系的根系浸入Magenta-GUS染液中,37℃反應6 h后在FAA固定液[V(甲醛)∶V(冰醋酸)∶V(70%乙醇)=1∶1∶18混合配制而成]中脫色固定10 h,鏡檢成像(BX50 OLYPUS顯微鏡)。
2 結果與分析
2.1番茄質膜H+-ATPase編碼基因的鑒定及序列分析
為了確認番茄質膜H+-ATPase基因家族中的成員數目,我們以已報道的3個番茄質膜H+-ATPase基因LHA1,LHA2和LHA4的全長編碼序列(CDS)作為查詢序列,采用BLASTN和TBLASTN程序分別在番茄基因組數據庫和EST/cDNA數據庫中進行同源序列搜索。通過對所獲得的序列進行相似性和結構域分析,最終篩選到8個具有完整編碼區的LHA基因,其中7個與之前報道的LHA1~7序列完全一致,而另外一個(本研究中命名為LHA8)為從未報道過的新基因。序列分析顯示,這8個LHA基因具有相近大小的開放閱讀框(ORF),且編碼的氨基酸序列之間具有很高的相似性(表2)。其中,LHA1和LHA2以及LHA5和LHA6之間的氨基酸序列相似性分別高達91.5%和98.3%。
2.2番茄質膜H+-ATPase編碼基因的外顯子/內含子結構分析
將8個LHA基因的編碼序列(CDS)和DNA序列比對后分析發現,這些基因的編碼區中都包含有多個內含子(圖1)。其中LHA1,LHA2和LHA8含有的內含子最多,為20個;而LHA4含有的內含子最少,只有10個。對內含子在LHA基因中的分布位置進一步分析發現,這8個LHA基因具有較為保守的內含子/外顯子結構特征,即各LHA基因中的不同內含子相對于外顯子的位置較為一致。從圖1中還可以得知,LHA基因在進化過程中存在著多處較為明顯的內含子丟失(intron loss)現象,如LHA5,LHA6和LHA7相對于LHA1,LHA2和LHA8這3個基因而言在第5個和第7個內含子處出現了丟失,導致了相應位置的外顯子發生了序列合并。

表2 番茄8個LHA基因的氨基酸序列相似性分析Table 2 Similarity matrix for the predicted amino acid sequences of the eight tomato LHA genes

圖1 番茄LHA基因的外顯子/內含子結構分析Figure 1 Exon/intron structures of the eight tomato LHA genes
2.3番茄質膜H+-ATPase家族基因的進化分析
對番茄LHA基因以及其他模式植物中同源基因序列進行的系統進化分析發現,植物質膜H+-ATPase基因家族可以大致分為5個亞家族(圖2),與之前其他課題組所報道的結果類似。除了番茄LHA3孤立于這5個亞家族之外,其他7個LHA基因分布于4個亞家族中,其中LHA1和LHA2分布在第I亞家族,且聚在同一進化分枝的末端,暗示了LHA1和LHA2的進化關系較近,復制產生這2個同源基因的事件發生在茄科植物與其他雙子葉植物分化之后。與此類似的是,聚集在第Ⅳ亞家族的LHA5,LHA6 和LHA7的進化關系也非常近。由于LHA5和LHA6之間的序列相似性非常高(表1),且外顯子/內含子的結構完全一致(圖1),暗示LHA5和LHA6可能是來源于同一個基因的復制,因此推測這2個基因可能具有相似的表達調控模式或行使相似的生物學功能。LHA4和LHA8分別處于第Ⅱ和第Ⅴ亞家族,與來自于其他雙子葉和單子葉植物中的同源基因具有相對較近的進化關系,暗示了LHA4和LHA8的進化起源較早,至少出現在雙子葉植物和單子葉植物分化之前。值得注意的是,番茄LHA基因的內含子/外顯子結構特征能夠很好地反應這些基因之間的進化關系,即在進化上具有較近關系的LHA同源基因,其內含子/外顯子的結構特征也較為接近。
2.4番茄質膜H+-ATPase家族基因的組織表達模式分析
圖3顯示為番茄LHA基因的組織表達特征。結果顯示:在正常培養條件下,除了LHA8在所有被分析的組織中都檢測不到表達信號外,其他7個LHA基因都能在某些或某個組織中被檢測到較為明顯的表達。具體而言,LHA1,LHA2和LHA4在所有的組織中都有表達,但在果實中的表達都相對較低,其中LHA2在所有組織中的表達豐度相對較高。盡管LHA3在所有被檢測的組織中也都有表達,但除了在花中有相對較高的表達之外,在其他組織中的表達都很微弱。LHA5,LHA6和LHA7這3個基因的表達模式比較特異,幾乎只在花中有表達,其中LHA5和LHA6的表達尤為強烈(圖3)。

圖2 番茄LHA基因及其他植物中同源基因的進化分析Figure 2 Phylogenetic analysis of tomato LHA genes and other plant homologs
2.5LHA8的表達調控模式分析
鑒于LHA8為新報道的基因,且在正常培養的番茄中不能被檢測到表達。為了驗證該基因的表達是否受到其他環境因素調控,本研究檢測了LHA8在不同養分(氮、磷、鉀和鎂)缺乏、高鹽(氯化鈉)脅迫以及接種菌根真菌處理條件下番茄根系中的表達水平(圖4)。結果表明:LHA8在養分缺乏和高鹽處理的番茄根系中也都檢測不到表達,但是在接種菌根真菌處理的番茄根系中能夠檢測到強烈的表達(圖4)。
為了更直觀地證實LHA8在被菌根真菌侵染根系中的表達部位,我們構建了LHA8啟動子片段(命名為pLHA8)融合GUS報告基因的雙元表達載體pLHA8-GUS,并通過農桿菌介導的轉基因方法轉化到了煙草中。對陽性轉基因煙草植株的GUS染色分析發現,LHA8的啟動子能夠驅動GUS報告基因在被菌根真菌侵染的根系中特異性表達,且表達部位主要集中在被菌根真菌菌絲侵入形成叢枝的細胞中表達,在未被菌絲侵染的細胞中檢測不到GUS表達(圖5)。
3結論與討論
質膜H+-ATPase是植物細胞膜上非常重要的功能蛋白,被認為是植物細胞代謝和生命活動過程的主宰酶[5]。迄今為止,質膜H+-ATPase的活性已被證實與植物生長發育過程中的很多生理過程(如細胞伸長生長、氣孔開閉、胞內pH調節、養分吸收等)密切相關[4]。鑒于質膜H+-ATPase功能的重要性,植物在漫長的進化過程中進化和保留了多個編碼質膜H+-ATPase的同源基因。如在模式植物擬南芥中已被報道存在有11個質膜H+-ATPase的編碼基因(AHA1~11)[7];在單子葉禾本科Gramineae水稻Oryza sativa中也已經被鑒定到10個具有轉錄活性的質膜H+-ATPase基因(OsA1~10)[6]。有意思的是,盡管番茄的基因組大小(約900 Mb)要比擬南芥(約125 Mb)和水稻的基因組(約460 Mb)要大得多[12],但通過對番茄全基因組序列搜索只獲得了8個質膜H+-ATPase編碼基因,序列分析表明,這些LHA基因具有較為保守的編碼序列和外顯子/內含子結構特征,其中新報道的LHA8與之前報道過的LHA1和LHA2的外顯子/內含子結構完全一致。
前期有研究表明,植物中的質膜H+-ATPase基因在進化上可以被大致分為5個亞家族(Ⅰ~Ⅴ)[17]。對擬南芥、煙草等植物中質膜H+-ATPase基因的表達分析顯示,處于第Ⅰ和第Ⅱ亞家族中的成員,其表達模式偏向于組成型,即幾乎在所有的組織器官中都有表達;而處于其他幾個亞家族中的成員的表達模式較為特異,只能在少數組織器官中有所表達[18]。在本研究中對番茄LHA基因的表達模式分析的結果也獲得了類似的結論。LHA1,LHA2和LHA4在所有被檢測的組織器官中都有表達,暗示了這幾個基因在番茄植株中可能參與了一系列的生理過程。LHA5,LHA6和LHA7表達模式比較特異,幾乎只在花中有表達,暗示了這幾個基因的功能主要與花器官和生殖器官發育相關,且這些基因之間可能存在著一定程度的功能冗余。之前有研究顯示,質膜H+-ATPase基因在轉錄水平上的表達受到很多因素(如養分缺乏、鹽害、病菌侵害和菌根真菌共生等)的影響[11]。LHA8無論是在正常培養條件下,還是在養分缺乏和高鹽處理中都不表達,但能夠在菌根真菌侵染的根系中強烈表達。菌根真菌是土壤中一類屬于球囊霉門Glomeromycota的能夠與大多數植物根系形成互惠共生關系的真菌[19]。形成菌根共生體后,菌根真菌的一端(根內菌絲)定殖于根系皮層細胞內,另一端(根外菌絲)穿越根際養分耗竭區。植物根系借助于菌根真菌的根外菌絲可數十倍地擴展在土壤中的吸收空間,增加對土壤中養分(主要是磷)的吸收利用;作為交換,植物提供給菌根真菌多達20%的光合產物以維持其生長繁殖[20]。LHA8在接種菌根真菌的番茄根系中特異性表達,暗示了該基因可能參與了植物與菌根真菌共生過程中的養分和信號交換過程。最近在蒺藜苜蓿Medicago truncatula和水稻中也各發現了1個菌根真菌共生特異誘導表達的質膜H+-ATPase基因(MtHA1和OsHA1/OsA8)[21-22]。在水稻中突變OsHA1/OsA8能夠顯著影響水稻植株根系與菌根真菌共生,并抑制水稻對土壤中磷素營養的吸收[22]。從進化樹上分析可知,LHA8與MtHA1和OsHA1/OsA8同處于第Ⅴ亞家族,暗示了LHA8在進化中保留了與MtHA1和OsHA1/OsA8相似的生物學功能。本研究通過對番茄質膜H+-ATPase基因家族各成員的鑒定、序列結構、進化關系和表達模式分析可以為將來深入研究質膜H+-ATPase在番茄植物生長發育過程中的作用機制提供理論依據。

圖3 番茄LHA基因的表達模式分析Figure 3 Expression analysis of the tomato LHA genes

圖4 LHA8在不同處理條件下番茄根系中的表達分析Figure 4 Expression analysis of LHA8 in roots of the tomato plants under different treatments
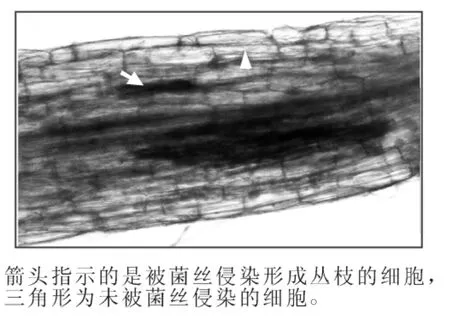
圖5 LHA8啟動子驅動GUS報告基因在轉基因煙草接種菌根真菌的根系中的表達分析Figure 5 Expression analysis of the GUS reporter driven by LHA8 promoter in transgenic tobacco roots colonized by arbuscular mycorrhizal fungi
[1]MORSOMME P,BOUTRY M.The plant plasma membrane H+-ATPase:structure,function and regulation[J].Biochim Biophysic Acta Biomembr,2000,1465(1/2):1-16.
[2]KANCZEWSKA J,MARCO S,VANDERMEEREN C,et al.Activation of the plant plasma membrane H+-ATPase by phosphorylation and binding of 14-3-3 proteins converts a dimer into a hexamer[J].Proc Natl Acad Sci USA,2005,102(33):11675-11680.
[3]SONDERGAARD T E,SCHULZ A,PALMGREN M G.Energization of transport processes in plants.Roles of the plasma membrane H+-ATPase[J].Plant Physiol,2004,136(1):2475-2482.
[4]ALSTERFJORD M,SEHNKE P C,ARKELL A,et al.Plasma membrane H+-ATPase and 14-3-3 isoforms of Arabidopsis leaves:evidence for isoform specificity in the 14-3-3/H+-ATPase interaction[J].Plant Cell Physiol,2004,45(9):1202-1210.
[5]GAXIOLA R A,PALMGREN M G,SCHUMACHER K.Plant proton pumps[J].FEBS Lett,2007,581(12):2204-2214.
[6]BAXTER I,TCHIEU J,SUSSMAN M R,et al.Genomic comparison of P-type ATPase ion pumps in Arabidopsis and rice[J].Plant Physiol,2003,132(2):618-628.
[7]HARUTA M,BURCH H L,NELSON R B,et al.Molecular characterization of mutant Arabidopsis plants with reduced plasma membrane proton pump activity[J].J Biol Chem,2010,285(23):17918-17929.
[8]ROBERTSON W R,CLARK K,YOUNG J C,et al.An Arabidopsis thaliana plasma membrane proton pump is essential for pollen development[J].Genetics,2004,168(3):1677-1687.
[9]LAN Ping,LI Wenfeng,LIN Wendai,et al.Mapping gene activity of Arabidopsis root hairs[J].Genome Biol,2013,14(16):1-20.
[10]BAXTER I R,YOUNG J C,ARMSTRONG G,et al.A plasma membrane H+-ATPase is required for the formation of proanthocyanidins in the seed coat endothelium of Arabidopsis thaliana[J].Proc Natl Acad Sci USA,2005,102(7):2649-2654.
[11]SCHALLER A,OECKING C.Modulation of plasma membrane H+-ATPase activity differentially activates wound and pathogen defense responses in tomato plants[J].Plant Cell,1999,11(2):263-272.
[12]SATO S,TABATA S,HIRAKAWA H,et al.The tomato genome sequence provides insights into fleshy fruit evolution[J].Nature,2012,485(7400):635-641.
[13]ROSEWARNE G M,SMITH F A,SCHACHTMAN D P,et al.Localization of proton-ATPase genes expressed in arbuscular mycorrhizal tomato plants[J].Mycorrhiza,2007,17(3):249-258.
[14]EWING N N,BENNETT A B.Assessment of the number and expression of P-Type H+-ATPase genes in tomato[J]. Plant Physiol,1994,106(2):547-557.
[15]CHEN Aiqun,CHEN Xiao,WANG Huimin,et al.Genome-wide investigation and expression analysis suggest diverseroles and genetic redundancy of Pht1 family genes in response to Pi deficiency in tomato[J].BMC Plant Biol,2014,14(1):61.doi:10.1186/1471-2229-14-61.
[16]HORSCH R B,FRY J E,HOFFMAN N L,et al.A simple and general method for transferring genes into plants[J]. Science,1985,227(4691):1229-1231.
[17]ARANGO M,GéVAUDANT F,OUFATTOLE M,et al.The plasma membrane proton pump ATPase:the significance of gene subfamilies[J].Planta,2003,216(3):355-365.
[18]ZHU Yiyong,DI Tingjun,XU Guohua,et al.Adaptation of plasma membrane H+-ATPase of rice roots to low pH as related to ammonium nutrition[J].Plant Cell Environ,2009,32:1428-1440.
[19]LIAO Dehua,CHEN Xiao,CHEN Aiqun,et al.The characterization of six auxin-induced tomato GH3 genes uncovers a member,SlGH3.4,strongly responsive to arbuscular mycorrhizal symbiosis[J].Plant Cell Physiol,2015,56(4):674-687.
[20]SMITH S E,SMITH F A,JAKOBSEN I.Functional diversity in arbuscular mycorrhizal(AM)symbioses:the contribution of the mycorrhizal P uptake pathway is not correlated with mycorrhizal responses growth or total P uptake[J]. New Phytol,2004,162(2):511-524.
[21]KRAJINSKI F,COURTY P E,SIEH D,et al.The H+-ATPase HA1 of Medicago truncatula is essential for phosphate transport and plant growth during arbuscular mycorrhizal symbiosis[J].Plant Cell,2014,26(4):1808-1817.
[22]WANG Ertao,YU Nan,BANO S A,et al.A H+-ATPase that energizes nutrient uptake during mycorrhizal symbioses in rice and Medicago truncatula[J].Plant Cell,2014,26(4):1818-1830.
Identification and expression analysis of tomato plasma membrane H+-ATPase family genes
LIU Jianjian,LIU Junli,JI Minjie,CHEN Jiadong,YANG Xiaofeng,CHEN Aiqun
(College of Resources and Environmental Sciences,Nanjing Agricultural University,Nanjing 210095,Jiangsu,China)
Plasma membrane(PM)H+-ATPase,which catalyzes ATP hydrolysis coupled with pumping out protons from cells and which is ubiquitously existent in all cell types of plants examined so far,has been characterized to be encoded by a multigene family involved in a number of plant physiological and developmental processes.The aim of this study is to identify and characterize the genes encoding PM H+-ATPase in Solanum lycopersicum(tomato).Through genomic sequence database hunting and bioinformatics analysis,eight putative PM H+-ATPase genes(named as LHA1-8)were identified,and their expression patterns were subsequently analysed by quantitative real-time reverse transcription polymerase chain reaction(qRT-PCR).Bioinformatics analysis revealed a high conservation among these paralogous genes in both coding sequences and Exon/Intron structures.Expression analysis showed that LHA1,LHA2,LHA3,and LHA4 were expressed in all tissues examined;whereas LHA5,LHA6,and LHA7 were predominantly expressed in flowers.LHA8,the newly identified gene in this study,with barely detectable transcripts in normal growth conditions or in nutrient(N,P,K,and Mg)-deficient conditions,was strongly induced in arbuscular mycorrhizal(AM)fungal-colonized roots.A 2 669 bp LHA8 promoter fragment could direct the β-glucuronidase(GUS)reporter expression specifically in transgenic tobacco roots colonized by AM fungi.The results provide new insights into the evolutionary conservation and functional divergence of the tomato PM H+-ATPase gene family during plant growth and development.[Ch,5 fig.2 tab.22 ref.]
molecular biology;Solanum lycopersicum(tomato);plasma membrane H+-ATPase;expression pattern;mycorrhizal fungi
S641.2;Q943.2
A
2095-0756(2016)05-0734-08
10.11833/j.issn.2095-0756.2016.05.002
2015-06-27;
2015-08-06
國家自然科學基金資助項目(31372121)
劉健健,從事叢枝菌根共生的分子機制研究。E-mail:jianjian2013103114@126.com。通信作者:陳愛群,副教授,博士,從事植物與微生物互作的分子機制研究。E-mail:chenaq8@163.com
浙 江 農 林 大 學 學 報,2016,33(5):734-741
Journal of Zhejiang A&F University
猜你喜歡
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
山東工業技術(2016年15期)2016-12-01 05:31:22
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
