福州左海湖細菌群落的季節性變化特征及典范對應分析
2016-10-28 01:28:27劉蘭英陳麗華李玥仁
福建農業學報 2016年7期
關鍵詞:環境
劉蘭英,呂 新,陳麗華,李玥仁*
(1.福建省農業科學院農業質量標準與檢測技術研究所,福建 福州 350003;2.福建省精密儀器農業測試重點實驗室,福建 福州 350003)
?
福州左海湖細菌群落的季節性變化特征及典范對應分析
劉蘭英1,2,呂新1,2,陳麗華1,2,李玥仁1,2*
(1.福建省農業科學院農業質量標準與檢測技術研究所,福建福州350003;2.福建省精密儀器農業測試重點實驗室,福建福州350003)
為揭示福州左海湖細菌群落的季節變化及其與環境因子的關系,通過構建16S rRNA 基因克隆文庫對全湖水體細菌群落進行了調查,應用典范對應分析(Canonical correspondence analysis,CCA)探討影響細菌群落季節性變化的環境因素。結果表明:左海湖細菌群落組成中β-變形菌門β-Proteobacteria和藍細菌門Cyanobacteria為優勢類群,α-變形菌門α-Proteobacteria、擬桿菌門Bacteroidetes和放線菌門Actinobacteria為亞優勢類群。細菌群落結構具有明顯的季節性變化特征,春、夏季Cyanobacteria為優勢類群,秋、冬季β-Proteobacteria為優勢類群。春季水體細菌群落結構主要受pH、葉綠素a(Chla)、透明度(SD)和總氮(TN)的影響,夏季水體受pH、Chla、SD、總磷(TP)影響較大,秋季水體受TP、Chla、TN影響較大,冬季水體則受水溫(WT)、pH、SD影響較大。本研究發現,左海湖細菌群落結構的季節動態與水溫及營養狀況密切相關。
左海湖;細菌群落;16S rRNA 基因克隆文庫;季節變化;典范對應分析
湖泊生態系統中細菌種類繁多且數量龐大,約105~108個·mL-1,占湖泊中生物量的絕大部分,是湖泊環境中食物網和食物鏈的重要組成部分[1- 2]。細菌對湖泊生態系統的穩定性具有調控作用,細菌群落結構是其整體功能的表現,湖泊細菌群落的組成與特性在一定程度上反映了湖泊生態系統的環境狀況[3]。因此研究湖泊細菌的群落組成、多樣性的季節差異,對于了解細菌在湖泊生態系統功能演替中的作用是非常必要的。
對水體細菌多樣性的研究已有很多報道,主要集中于湖泊、水庫、溪流、海洋等環境[4],尤其是湖泊細菌多樣性研究越來越受到研究者的關注。許多研究表明,湖泊營養狀況不同,湖泊細菌的群落結構組成也各不相同[5-6]。Zhang等[7]借助克隆文庫和基因芯片相結合的分子生物學技術,對青藏高原高山湖泊細菌多樣性的研究表明,鹽濃度和海拔與該湖泊中的細菌群落結構呈顯著性相關。Adams等[8]通過研究北極24個湖泊和溪流樣品中的細菌多樣性,發現水體的溫度在這些生境的細菌群落組成中起著關鍵作用。
左海湖位于福州市西北部,是典型的半封閉式淺水湖泊,是福州市民主要的休閑和娛樂場所之一。自2008年發現被稱為“左海水怪”的不名果凍狀生物體以來,左海湖的水體質量狀況已引起了廣泛的社會關注。“水怪”的出現可能與左海湖環境因子發生了變化有關,而湖泊中微生物群落的動態變化對湖泊環境具有較好的指示作用[9]。本研究在左海湖設置4個采樣點,基于構建16S rRNA基因克隆文庫的方法,對湖區細菌群落組成及季節變化規律進行了研究,并分析細菌群落組成與水體環境因子間的關系,以期為左海湖水環境質量評價及污染防治提供理論依據。
1 材料與方法
1.1樣品采集
分別于2012年2月(冬季)、4月(春季)、8月(夏季)和10月(秋季)在左海湖進行水樣采集。設置4個采樣點,具體采樣位置的經緯度信息見表1,每個取樣站位設置3個重復,所有樣品采集完成后立即放入帶有冰塊的保溫箱中運回實驗室,用于后續試驗分析。
1.2左海湖環境因子的測定
采用透明度盤對水體的透明度(SD)進行現場測定,其他環境因子均在實驗室內完成測定。水體的pH值通過酸度計(PE20,梅特勒)測定;采用90%丙酮萃取熒光分光光度法對水體的葉綠素a含量進行測定[10];總氮(TN)、總磷(TP)含量和高錳酸鉀指數(COD)測定方法均參照王心芳等[11]的報道。
1.3水體細菌的收集與基因組DNA的提取
采用8 μm孔徑的定性濾紙,將水樣中的大部分顆粒雜質及真核生物過濾除去,再通過聚碳酸酯微孔薄膜(Whatman,0.2 μm,47 mm) 進行真空抽濾,然后用無菌水洗滌濾膜2~3次,取下濾膜并將其剪碎后裝入1.5 mL離心管中,于-20℃保存備用。水體細菌總基因組DNA的提取參考Wu等[12]的報道。
1.4細菌16S rRNA基因克隆文庫構建與篩選
采用細菌16S rRNA序列通用型引物27f (5′-AGA GTT TGA TCC TGG CTC AG-3′) 和1492r (5′-TAC GGC TAC CTT GTT ACG ACT T-3′)[13]進行PCR擴增,設置3組試驗重復,PCR擴增方法參考Wu等[4]的報道。反應程序:①94℃,3 min;②94℃,30 s;③60℃,1 min;④72℃,60 s。共30個循環,最后再72℃延伸10 min。混合3 組平行的PCR擴增產物,經純化后的PCR產物連接到pMD18-T載體(TaKaRa公司),轉化E.coliDH5α感受態細胞。通過藍白斑法篩選陽性克隆,用測序引物M13f和M13r進行菌落PCR鑒定,PCR擴增體系和反應條件同上。
1.5酶切分析與測序
將驗證為陽性重組子的質粒DNA進行酶切鑒定,所用的限制性內切酶為Csp6 I和Hinf I[14],通過2.5%~3.0% 瓊脂糖凝膠電泳分離驗證酶切產物。借助Photoshop軟件分析酶切譜型,統計各泳道的酶切條帶數和大小,將完全相同的酶切譜型進行歸類,并計算每個酶切譜型出現的頻率,挑取具有不同譜型的克隆子到生工生物工程(上海)有限公司進行測序。測序獲得的16S rRNA基因序列通過DNAMAN 8.0軟件進行相互比較,將16S rRNA基因序列相似性大于或者等于97% 的劃為一個操作分類單元 (Operational Taxonomic Units,OTU)。
1.6數據分析
細菌多樣性根據Shannon-Wiener指數(H′)和Simpson指數(1/D)[15]進行分析,均勻度指數(E)可以反映群落中各個物種多度分布的均勻程度,其計算參考邱小琮等[16]的報道,采用文庫覆蓋率(Coverage C)評估所構建的文庫對目標環境微生物多樣性的體現程度[4]。具體計算公式如下:
(1)
(2)
式中,Pi=ni/N,ni為第i種OTU的克隆數,N為總克隆數。
E=H′/Hmax
(3)
式中,Hmax為最大Shannon指數,大小為log2S,S為克隆文庫中總OTU種類數。
C=(1-n1/N)×100%
(4)
式中,n1代表在16SrRNA基因克隆文庫中僅出現一次的OTU數量,N為總克隆數。
通過CanocoforWindows4.5應用軟件對群落組成數據和環境因子數據進行典范對應分析,探討細菌群落與環境因子之間的關系,分析時除了pH值,其余數據均進行log(x+1)轉換[17-18],采用Origin8.0軟件對試驗數據進行分析并作圖。
2 結果與分析
2.1環境因子季節動態
左海湖各環境因子指標呈現季節性的變化規律(表1)。湖水各季節的pH值介于7.1~7.9,均呈弱堿性;水溫(WT)變化幅度為12.8~32.2℃,四季分明;透明度(SD)的變化范圍介于20~48cm,在秋季呈現最低值;化學需氧量(COD)的變化幅度較小,波動范圍介于7.63~10.51mg·L-1;總氮(TN)含量的變化幅度較大,在0.39~1.28mg·L-1之間波動,其中夏季各采樣點的總氮含量均較高;總磷(TP)含量的波動范圍在0.023~0.052mg·L-1,在夏季呈現最高值,冬季呈現最低值。葉綠素a(Chla)含量的變化范圍介于0.012~0.048mg·L-1,在冬季呈現最低值,其余季節變化規律不顯著。

表1 左海湖水體理化指標
注:①S1~S4代表左海湖4個不同的采樣站位;②表格中的數值代表平均值±標準差(3組平行試驗的標準差)。
2.2克隆文庫分析
每個季節樣品分別構建1個16S rRNA基因克隆文庫,并在每個文庫中隨機挑取120個克隆子。通過菌落PCR驗證后共得到412個陽性克隆子,其中春季115個,夏季105個,秋季101個,冬季91個。通過限制性內切酶Csp6Ⅰ和HinfⅠ對4個季節文庫中獲得的陽性克隆子進行酶切鑒定,共計得到298個酶切結果。
分別計算4個季節文庫的覆蓋率Coverage(C)、Shannon-Wiener指數(H′)、Simpson 指數(1/D)和均勻度指數(E)(表2),以比較各季節樣品的細菌群落多樣性與豐富度。從表2可以看出,各季節樣品的文庫覆蓋率(C)為69.5% ~79.1%。張勝男等[19]在梁素海浮游細菌群落結構研究中指出,當湖區克隆文庫的覆蓋率為65.2%~75.4%時,表明文庫基本可代表細菌群落結構的組成。因此,由表2中的文庫覆蓋率范圍可知,本研究所構建的克隆文庫較為理想,可以很好地反映左海湖中細菌群落的多樣性。
多樣性分析表明,春、夏、秋、冬各季節文庫的Shannon-Wiener指數分別為2.85、3.30、3.29和3.52,其中冬季樣品的HShannon-Wiener指數和HSimpson指數均為最高,說明其細菌群落多樣性最豐富;相比之下,春季樣品的HShannon-Wiener指數和HSimpson指數均較低,說明其細菌群落多樣性較低。從均勻度指數(E)可以看出,各克隆文庫中細菌群落的均勻度較為接近,和多樣性指數變化趨勢一致。
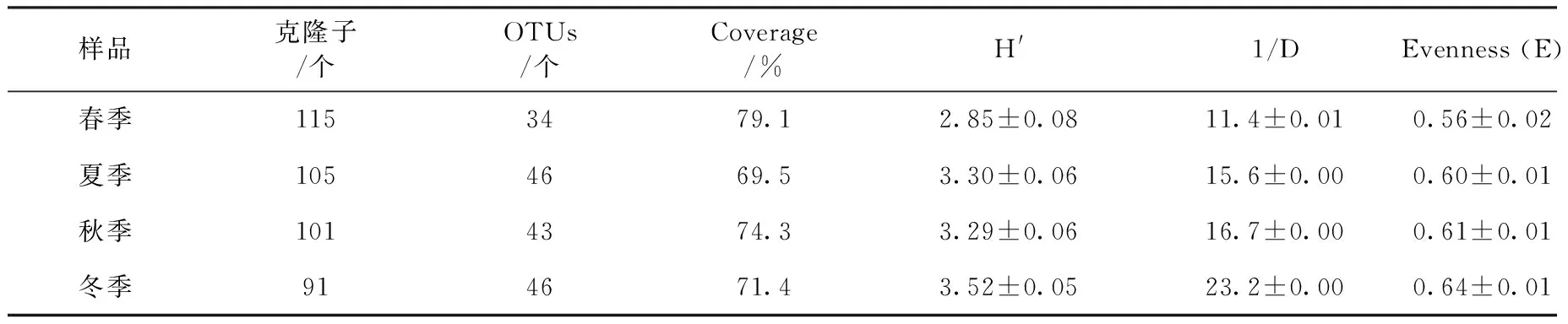
表2 左海湖細菌16S rRNA基因克隆文庫分析結果
2.3細菌群落組成的季節動態
將所構建克隆文庫的16S rRNA基因片段送上海生工進行雙向測序,測序結果中的上、下游序列通過Invitrogen軟件拼接。根據97%序列相似性原則,將所有季節文庫中得到的克隆子劃分為169個OTUs類型。BLAST比對結果顯示,大部分OTUs(90%以上)的16S rRNA基因序列與GenBank 中已知序列具有很高的相似度(97%~100%)。本試驗中獲得的412個陽性克隆子代表的169種基因型歸屬為11門一級的細菌分類單元,分別來自變形菌門的α-,β-,γ-和δ-亞族、放線菌門Actinobacteria、擬桿菌門Bacteroidetes、藍細菌門Cyanobacteria、綠彎菌門Chloroflexi、酸桿菌門Acidobacteria、疣微菌門Verrucomicrobia和厚壁菌門Firmicutes。
不同細菌類群組成的季節性變化較大(圖1),在各季節中均大量分布的細菌類群是β-Proteobacteria和Cyanobacteria。其中,β-Proteobacteria在秋、冬季比例較高,在春季比例相對較低,為秋、冬季文庫的優勢細菌類群;Cyanobacteria在春、夏季最豐富,尤其在春季,該細菌類群占了絕對的優勢地位。α-Proteobacteria在4個不同季節中均有發現,春冬季比例較高,夏秋季比例較低。Actinobacteria和Bacteroidetes在不同季節里都有分布,冬季數量較多,其他季節較少。除了上述幾大主要的細菌類群,一些非優勢類群,如Chloroflexi、Verrucomicrobia、Firmicutes等并未在所有季節里均有檢測到,而且所占的比例也較少,這可能與研究中所采用的16S rRNA基因文庫構建方法無法很好地獲取環境中的非優勢細菌類群有關。
2.4細菌群落與環境因子的CCA分析
不同季節細菌群落組成與環境因子之間的關系如圖2所示。依據除趨勢對應分析(DCA),物種分布的最大梯度長度為2.040(<3.0),具有明顯的線性模型特征[20],因此選擇冗余分析(RDA)探討細菌群落組成與環境因子之間的相關性。本研究中環境因子第1、2主軸間的相關系數為0,說明排序結果是可信的,圖中箭頭所處的象限表示環境因子與排序軸間的正負相關性[21-22]。從RDA分析圖中可以看出,不同季節細菌群落受環境因子影響不同,本研究選取的7 個環境因子對細菌群落結構都有一定程度的影響。春季水體細菌的分布主要受Chla、pH、SD和TN的影響,排序軸1與Chla(r= -0.959 1)、pH(r=-0.873 7)、SD(r=-0.814 4)呈負相關,軸2與TN(r=-0.767 0)呈負相關;夏季水體細菌的分布受pH、SD、TP、Chla的影響較大,軸1與pH(r=-0.960 7)、TP(r=-0.769 3)、Chla(r=-0.742 7)呈負相關,軸2與SD(r=-0.796 1)負相關;秋季水體細菌的分布主要受TP、Chla、TN的影響,環境因子與主軸1的相關性較大,其中呈正相關的為TP(r=0.995 3)、TN(r=0.920 8),與Chla(r=-0.941 1)呈負相關;冬季水體細菌的分布受WT、pH、SD影響較大,環境因子與主軸2的相關性較大,其中呈正相關的為pH(r=0.985 3),呈負相關的為WT(r=-0.983 3)和SD(r=-0.793 4)。
3 討論與結論
與其他已知淡水生境細菌類群的組成情況[4-5]相似,本研究結果表明,左海湖水體中常見的細菌類群為α-變形菌門α-Proteobacteria、β-變形菌門β-Proteobacteria、放線菌門Actinobacteria、擬桿菌門Bacteroidetes和藍細菌門Cyanobacteria,其季節分布特征表現為β-Proteobacteria和Cyanobacteria在所有季節里所占的比例均最高。春、夏季Cyanobacteria為優勢類群,秋、冬季β-Proteobacteria為優勢類群。β-Proteobacteria和Cyanobacteria通常被認為在富營養化水體中占優勢[4, 6],因此在左海湖檢測到豐富的β-Proteobacteria和Cyanobacteria細菌類群在某種程度上說明了左海湖水質狀況發生了變化。而優勢類群顯著的季節性變化,原因可能為春、夏季氣溫高,有利于Cyanobacteria的生長,因此Cyanobacteria數量通常有所上升,甚至占了絕對的優勢地位,由于種間競爭的加劇,導致β-Proteobacteria數量有所下降。另外,β-Proteobacteria和Cyanobacteria在季節性變化過程中存在明顯的此消彼長的規律,還可能與水體中營養物質發生變化有關[23],水體中營養物質的高低會導致一些細菌的生長受到促進或抑制。
本研究對福州左海湖水體的細菌群落組成、環境因子進行典范對應分析,結果表明環境因子對不同季節里細菌群落組成的影響程度明顯不同。造成左海湖細菌群落結構變化的環境因子包括水體的pH、水溫、總氮、總磷和葉綠素a,表明左海湖細菌群落結構具有明顯的季節變化,并且與湖泊中的環境因子有著不可分割的密切關系,這與Pollet等[24],Du等[25],Wang等[26]的研究結果類似。氮、磷是水體主要營養污染物[27],通過環境因子季節動態分析發現,春、夏、秋季左海湖水的總氮、總磷含量均高于冬季,表明這期間湖水富營養化程度比冬季湖水嚴重。已有研究表明,富營養化湖泊中碳酸鹽的化學平衡決定著水體pH的變化,而碳酸鹽體系的化學平衡又受到藻類種群生長、轉化,光合、呼吸作用等過程的影響,藻類初級生產力與水體的pH相關性顯著[28-29]。本研究中細菌群落組成與水體pH的相關性可能是由不同水環境條件下藍細菌的生長繁殖變化導致的。藍細菌是一類光能營養微生物,其數量在春、夏季最豐富,這可能與春、夏季水體具有更為豐富的有機營養,較高的水體溫度以及較強的光照強度等有關。另有研究表明葉綠素a含量的高低體現了藻類生物量的多少[30],本研究中我們發現藍細菌在春、夏、秋季都比較豐富,而葉綠素a含量在這3個季節里也都比較高。因此,認為左海湖細菌群落與水體葉綠素a含量的相關性也可能是由不同季節時期藍細菌的生長繁殖變化導致的。
本研究通過構建16S rRNA基因克隆文庫的方法,探討了福州左海湖中不同季節細菌群落結構的組成,該方法對于細菌群落多樣性研究有一定幫助,但也存在一定的局限性。該方法主要采用隨機挑選克隆子的方法,可能會高估目標環境中優勢細菌群落的豐富度,不能了解環境中的非優勢細菌群落,而那些數量上不占優勢的細菌類群可能對水體環境也起著重要的作用。近年來流行的高通量測序技術可以更為精確的研究環境中微生物群落的多樣性,已逐漸得到廣泛應用,今后研究中可結合高通量測序技術對左海湖中的細菌群落組成進行更全面認識。
[1]SUN J G, JIANG R B, REN T Z, et al. Prospect for farm land and water pollution and microorganism repair in China[J]. Chinese Journal of Agricultural Resources and Regional Planning, 2008, 29(1): 41-47.
[2]LIU H X, SONG X Y, HUANG L M, et al. Progress in the mechanism of modulation on marine bacterial production[J]. Ecological Science, 2008, 27(1): 61-64.
[3]YANNARELL A C, KENT A D, LAUSTER G H, et al. Temporal patterns in bacterial communities in three temperate lakes of different trophic status[J]. Microb Ecol, 2003, 46: 391-405.
[4]WU L, GE G, ZHU G F, et al. Diversity and composition of the bacterial community of Poyang Lake (China) as determined by 16S rRNA gene sequence analysis[J]. World J Microbiol Biotechnol, 2012, 28: 233-244.
[5]LIU Z, HUANG S, SUN G, et al. Phylogenetic diversity, composition and distribution of bacterioplankton community in the Dongjiang River, China[J]. FEMS Microbiol Ecol, 2012, 80(1): 30-44.
[6]CHEN Z B, ZHOU Z Y, PENG X, et al. Effects of wet and dry seasons on the aquatic bacterial community structure of the Three Gorges Reservoir[J]. World J Microbiol Biotechnol, 2013, 29(5): 841-853.
[7]ZHANG R, WU Q, PICENO Y M, et al. Diversity of bacterioplankton in contrasting Tibetan lakes revealed by high-density microarray and clone library analysis[J]. FEMS Microbiol Ecol, 2013, 86(2): 277-287.
[8]ADAMS H E, CRUMP B C, KLING G W. Temperature controls on aquatic bacterial production and community dynamics in arctic lakes and streams[J]. Environmental Microbiology, 2010, 12(5): 1319-1333.
[9]TAMAKI H, SEKIGUCHI Y, HANADA S, et al. Comparative analysis of bacteria diversity in freshwater sediment of a shallow eutrophic lake by molecular and improved cultivation-based techniques[J]. Applied and Environmental Microbiology, 2005, 71: 2162-2169.
[10]楊彩根, 宋學宏, 孫丙耀. 浮游植物葉綠素a含量簡易測定方法的比較[J]. 海洋科學, 2007, 31(1): 6-9.
[11]王心芳, 魏復盛, 齊文啟. 水和廢水監測分析方法. [M]. 第4版. 北京: 中國環境科學出版社, 2002:216-219.
[12]WU Q L, ZWART G, SCHAUER M, et al. Bacterioplankton community composition along a salinity gradient of sixteen high-mountain lakes located on the Tibetan Plateau, China[J]. Applied and Environmental Microbiology, 2006, 72(8): 5478-5485.
[13]MWIRICHIA R, COUSIN S, MUIGAI A W, et al. Bacterial Diversity in the Haloalkaline Lake Elmenteita, Kenya[J]. Curr Microbiol, 2011, 62: 209-221.
[14]白藍, 趙明文, 賈軍偉, 等. 16S rDNA 克隆文庫法探索轉基因香石竹對土壤細菌群落的影響[J]. 微生物學通報, 2012, 39(4): 435-447.
[15]張繼強, 陳文業, 康建軍,等. 敦煌西湖鹽化草甸蘆葦群落特征及多樣性沿水分梯度的分布格局[J]. 水土保持通報, 2013, 33(2): 173-176.
[16]邱小琮, 趙紅雪. 寧夏沙湖浮游植物群落結構及多樣性研究[J]. 水生態學雜志, 2011, 32(1): 20-26.
[17]LIU X, LU X, CHEN Y. The effects of temperature and nutrient ratios on Microcystis blooms in Lake Taihu, China: An 11-year investigation[J]. Harmful Algae, 2011, 10(3): 337-343.
[18]MARIA J, MORA R, SCRANTON M I, et al. Bacterial community composition in a large marine anoxic basin: a Cariaco Basin time-series survey[J]. FEMS Microbiol Ecol, 2013, 84: 625-639.
[19]張勝男, 趙吉睿, 張曉軍, 等. 烏梁素海浮游細菌群落結構及其對富營養化因子的響應[J]. 微生物學通報,2014, 41(6): 1082-1093.
[20]鐘晶晶, 劉茂松, 王玉, 等. 太湖流域河流與湖泊間主要水質指標的空間關聯特征[J]. 生態學雜志, 2014, 33(8): 2176-2182.
[21]TER BRAAK C J F. Canonical correspondence analysis:A new eigenvector technique for multivariate direct gradient analysis[J]. Ecology, 1986, 67(5): 1167-1179.
[22]李玉華, 許其功, 趙越, 等. 松花湖水體中不同空間分布的細菌群落結構分析[J]. 農業環境科學學報, 2013, 32(4): 764-770.
[23]王曉丹, 翟振華, 趙爽, 等. 北京翠湖表流和潛流濕地對細菌多樣性的影響[J]. 環境科學, 2009, 30(1): 280-288.
[24]POLLET T, TADONLéKé R D, HUMBERT J F. Spatiotemporal changes in the structure and composition of a less-abundant bacterial phylum (Planctomycetes) in two perialpine lakes[J]. Applied and environmental microbiology, 2011, 77(14): 4811-4821.
[25]DU J, XIAO K, LI L, et al. Temporal and Spatial Diversity of Bacterial Communities in Coastal Waters of the South China Sea[J]. PLoS One, 2013, 8(6): e66968.
[26]WANG J J, ZHANG Y, LI Z G, et al. Higher seasonal variation of actinobacterial communities than spatial heterogeneity in the surface sediments of Taihu Lake, China[J]. Can J Microbiol, 2013, 59(5): 353-358.
[27]孫寓姣, 程陳, 丁愛中, 等. 官廳水庫水質特征及水體微生物多樣性的響應[J]. 中國環境科學, 2015, 35(5): 1547-1553.
[28]朱文昌, 陸敏, 石浚哲. 梅梁湖水體浮游植物與環境因子的關系[J]. 環境監測管理與技術, 2010, 22(3): 27-30.
[29]楊麗標, 韓小勇, 孫璞, 等. 巢湖藻類組成與環境因子典范對應分析[J]. 農業環境科學學報, 2011, 30(5): 952-958.
[30]馮勝, 李定龍, 秦伯強. 太湖水華過程中微生物群落的動態變化[J]. 寧波大學學報:理工版, 2010, 23(1): 7-12.
(責任編輯:林海清)
Seasonal Changes on and Environmental Factors Affecting Bacterial Community at Zuohai Lake in Fuzhou
LIU Lan-ying1, 2, Lü Xin1, 2, CHEN Li-hua1, 2, LI Yue-ren1, 2*
(1.InstituteforAgriculturalStandardsandTestingTechnology,FujianAcademyofAgriculturalScience,Fuzhou,Fujian350003,China; 2.KeyLaboratoryforPrecisionInstrumentTestsinAgriculturalFields,Fuzhou,Fujian350003,China)
To determine the seasonal changes on and the environmental factors that affected the bacterial community at Zuohai Lake in Fuzhou, a survey was conducted at 4 sampling sites on the lake from February to August in 2012. A genetic library of their 16S rRNA gene clones was established. Canonical correspondence analysis (CCA) was used to examine the correlation between the bacterial diversity and the environmental factors. The results indicated thatβ-ProteobacteriaandCyanobacteriawere the predominate lineages in the water, followed byα-Proteobacteria,Bacteroidetes, andActinobacteria. A significant seasonal variation in composition of the bacterial community was detected. Among various categories of bacteria,Cyanobacteriaappeared most dominant in the spring and summer; whileβ-Proteobacteria, in the autumn and winter. The results of CCA showed that the bacterial community in the water was mainly affected by pH, chlorophyll a (Chla), transparency (SD), and total nitrogen (TN) in the spring; pH, Chla, SD, and total phosphate(TP), in the summer; TP, Chla, and TN in the fall; and, water temperature, pH, and SD, in the winter. Thus, in general, the bacterial growth in the lake closely related to the temperature as well as the nutrients available in the water.
Zuohai lake; bacterial community; 16S rRNA gene clone library; seasonal changes; canonical correspondence analysis
2016-03-22初稿;2016-04-30修改稿
劉蘭英(1987-),女,研究實習員,研究方向:環境微生物多樣性
李玥仁(1966-),男,研究員,研究方向:微生物多樣性與生物安全(E-mail: yuerenli@yeah.net)
福建省自然科學基金項目(2011J01118);福建省科技計劃項目——省屬公益類科研院所基本科研專項(2015R1025-9)
Q 938.8; X 172
A
1008-0384(2016)07-758-07
劉蘭英,呂新,陳麗華,等.福州左海湖細菌群落的季節性變化特征及典范對應分析[J].福建農業學報,2016,31(7):758-764.
LIU L-Y,Lü X,CHEN L-H,et al.Seasonal Changes on and Environmental Factors Affecting Bacterial Community at Zuohai Lake in Fuzhou[J].FujianJournalofAgriculturalSciences,2016,31(7):758-764.
猜你喜歡
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
新世紀智能(英語備考)(2019年12期)2020-01-13 06:07:18
瘋狂英語·新策略(2019年9期)2019-10-17 01:51:34
中國生殖健康(2019年11期)2019-01-07 01:28:02
中國生殖健康(2018年6期)2018-11-06 07:09:28
濰坊學院學報(2017年2期)2017-04-20 08:44:31
中國環境監察(2016年5期)2016-10-24 05:25:52
中國商論(2016年33期)2016-03-01 01:59:38
