支鏈氨基酸影響豬免疫功能的研究進(jìn)展
2016-11-08 04:51:50董小英李均祥任曉強(qiáng)唐勝球
中國(guó)飼料 2016年16期
董小英,李均祥,任曉強(qiáng),唐勝球*
(1.韶關(guān)學(xué)院英東農(nóng)業(yè)科學(xué)與工程學(xué)院,廣東韶關(guān)512005;2.韶關(guān)學(xué)院粵北生豬生產(chǎn)及疾病防控協(xié)同創(chuàng)新發(fā)展中心,廣東韶關(guān)512005;3.韶關(guān)學(xué)院英東生命科學(xué)學(xué)院,廣東韶關(guān)512005)
綜述
支鏈氨基酸影響豬免疫功能的研究進(jìn)展
董小英1,2,李均祥3,任曉強(qiáng)1,唐勝球1,2*
(1.韶關(guān)學(xué)院英東農(nóng)業(yè)科學(xué)與工程學(xué)院,廣東韶關(guān)512005;2.韶關(guān)學(xué)院粵北生豬生產(chǎn)及疾病防控協(xié)同創(chuàng)新發(fā)展中心,廣東韶關(guān)512005;3.韶關(guān)學(xué)院英東生命科學(xué)學(xué)院,廣東韶關(guān)512005)
支鏈氨基酸(BCAAs)作為豬體內(nèi)的必需氨基酸,具有營(yíng)養(yǎng)支持、調(diào)節(jié)免疫、促進(jìn)分泌和參與代謝等多種重要的生物學(xué)功能。本文主要就支鏈氨基酸影響豬免疫功能的新近成果作一綜述,并初步探討其作用機(jī)理,以期為相關(guān)研究提供科學(xué)參考。
支鏈氨基酸;豬;免疫功能;機(jī)理
支鏈氨基酸(BCAAs)又名分支氨基酸,是指一組化學(xué)結(jié)構(gòu)類似,在α-碳上含有分支脂肪烴鏈的中性氨基酸,為亮氨酸(Leu)、異亮氨酸(Ile)與纈氨酸(Val)三者的統(tǒng)稱。研究證實(shí),BCAAs均不能在人和動(dòng)物體內(nèi)自身合成,必須由外界食物供給,是機(jī)體維持正常生命功能和生長(zhǎng)發(fā)育所必需的氨基酸,在機(jī)體中具有提供能量、營(yíng)養(yǎng)支持、調(diào)節(jié)免疫,以及促進(jìn)激素分泌、參與蛋白合成和生理代謝等多種重要的生物學(xué)功能(劉春生等,2006;Harris等,2005)。BCAAs作為唯一在肝外代謝的氨基酸,主要分布在骨骼肌,約占骨骼肌蛋白質(zhì)中必需氨基酸的35%,是體內(nèi)氨基酸系統(tǒng)供能的重要來(lái)源。本文主要就BCAAs的免疫調(diào)節(jié)機(jī)制及其在提高豬免疫功能上的應(yīng)用進(jìn)行概述。
1 BCAAs調(diào)節(jié)免疫功能的機(jī)理
1.1影響免疫細(xì)胞功能動(dòng)物機(jī)體的免疫系統(tǒng)由免疫組織、免疫器官、免疫細(xì)胞及免疫活性分子等組成,具有免疫監(jiān)視、防御與調(diào)控等生理作用,能及時(shí)發(fā)現(xiàn)并清除異物、外來(lái)病毒或病原微生物等引起內(nèi)環(huán)境波動(dòng)的因素。BCAAs之所以能有效調(diào)節(jié)動(dòng)物機(jī)體的免疫功能,其機(jī)理之一就是該類氨基酸為維持淋巴細(xì)胞敏感性與免疫細(xì)胞功能所必需的功能物質(zhì),可直接影響到免疫器官細(xì)胞的發(fā)生、分化、發(fā)育與成熟。免疫學(xué)研究指出,補(bǔ)充BCAAs可提高骨骼肌中該類氨基酸的能量代謝,同時(shí)促進(jìn)其向谷氨酰胺轉(zhuǎn)變;而谷氨酰胺是淋巴細(xì)胞和嗜中性粒細(xì)胞發(fā)揮正常功能的重要能源物質(zhì),可間接提高白細(xì)胞與淋巴細(xì)胞的增殖性能,以改善機(jī)體免疫作用(Negro等,2008)。
研究發(fā)現(xiàn),動(dòng)物攝入BCAAs不足可導(dǎo)致體內(nèi)免疫球蛋白水平升高,引起免疫功能異常亢進(jìn),進(jìn)而對(duì)自身器官或組織產(chǎn)生傷害,嚴(yán)重時(shí)可導(dǎo)致胸腺與脾臟等免疫器官萎縮,淋巴組織明顯受損;現(xiàn)已證實(shí),纈氨酸在免疫球蛋白中所占的比重較其他氨基酸都高,若動(dòng)物體內(nèi)缺乏該氨基酸還會(huì)導(dǎo)致補(bǔ)體C3和運(yùn)鐵蛋白水平急劇降低,顯著抑制胸腺與外周淋巴組織的正常生長(zhǎng)發(fā)育,并抑制嗜酸性粒細(xì)胞及嗜中性粒細(xì)胞的增殖分化,甚至出現(xiàn)淋巴細(xì)胞功能嚴(yán)重耗竭的后果(陳熠等,2008)。還有文獻(xiàn)報(bào)道,纈氨酸具有促進(jìn)骨骼T細(xì)胞轉(zhuǎn)化為成熟T細(xì)胞的作用(徐琪壽,1996;Petro等,1981)。Dunshea等(2005)利用細(xì)胞培養(yǎng)研究發(fā)現(xiàn),亮氨酸是淋巴細(xì)胞合成蛋白、DNA、RNA和分化應(yīng)激反應(yīng)所必需的;經(jīng)鼠的在體試驗(yàn)表明,亮氨酸還能限制機(jī)體免疫功能多方面消弱,加強(qiáng)病原體的受損傷程度。
此外,靳繼德等(2001)報(bào)道,適量增加BCAAs攝入有助于改善創(chuàng)傷后機(jī)體的營(yíng)養(yǎng)代謝狀況,可有效增強(qiáng)免疫防御功能,并能促進(jìn)小腸黏膜細(xì)胞的增殖。最近研究發(fā)現(xiàn),在飲水中添加0.5%或2.5%異亮氨酸可提高鼠血清抗氧化能力,促使脾臟脾小結(jié)增大、增多,還能增厚動(dòng)脈周圍的淋巴鞘,并且淋巴細(xì)胞的排列更為密集(宮碧霜等,2015)。
1.2調(diào)節(jié)免疫因子活性BCAAs參與機(jī)體免疫機(jī)能的生理調(diào)節(jié),其作用機(jī)制還與它促進(jìn)免疫因子活性具有密切關(guān)系。陳熠等(2008)報(bào)道,BCAAs可通過(guò)影響免疫活性物質(zhì)中免疫球蛋白的形成或活性等途徑,以實(shí)現(xiàn)對(duì)機(jī)體免疫功能的調(diào)控作用。如纈氨酸缺乏會(huì)顯著阻礙胸腺與外周淋巴組織細(xì)胞的正常生長(zhǎng),進(jìn)而抑制嗜中性白細(xì)胞的增殖,可明顯降低免疫球蛋白水平、補(bǔ)體C3和運(yùn)鐵蛋白的水平(Calder,2006;Petro等,1981;Chevalier等,1977)。然而,Petro(1980)在研究BCAAs影響小鼠血中免疫球蛋白、補(bǔ)體C3及運(yùn)鐵蛋白水平時(shí)發(fā)現(xiàn),BCAAs中任何一種氨基酸缺乏均可使免疫球蛋白水平有所升高,而補(bǔ)體C3僅受異亮氨酸缺乏的影響,運(yùn)鐵蛋白對(duì)支鏈氨基酸缺乏卻不敏感,但異亮氨酸缺乏可降低小鼠腹腔滲出液中的運(yùn)鐵蛋白水平;此外,當(dāng)給動(dòng)物接種熱處理過(guò)的傷寒沙門(mén)氏菌后,纈氨酸缺乏可使補(bǔ)體C3、血凝集素、運(yùn)鐵蛋白水平顯著下降,亮氨酸缺乏僅使補(bǔ)體C3水平明顯下降,而異亮氨酸缺乏卻無(wú)顯著影響。Calder(2006)報(bào)道,小鼠缺乏亮氨酸可使補(bǔ)體C3水平降低,缺乏異亮氨酸還能使動(dòng)物腹腔滲出液中運(yùn)鐵蛋白水平下降。另有研究證實(shí),腹腔注射纈氨酸可增加綿羊紅細(xì)胞免疫后的脾免疫球蛋白M(IgM)分泌細(xì)胞數(shù)(Belokrylov等,1990)。
黃紅英等(2007)報(bào)道,與對(duì)照組相比,在泌乳母豬日糧中添加BCAAs可極顯著提高仔豬血清中免疫球蛋白G(IgG)的濃度(P<0.01),研究結(jié)果揭示在母豬日糧中添加BCAAs能通過(guò)母體效應(yīng)影響仔豬的生長(zhǎng)發(fā)育與免疫功能,且其作用效果與添加水平有密切關(guān)系。研究證實(shí),IgG是動(dòng)物自然感染和人工主動(dòng)免疫后機(jī)體產(chǎn)生的主要抗體,在體內(nèi)可發(fā)揮抗菌、抗病毒、抗毒素等免疫學(xué)功能(陶義訓(xùn),1999)。還有試驗(yàn)指出,在母豬日糧中添加纈氨酸,可顯著提高其泌乳第10、18 d血清白蛋白濃度(P<0.05),同時(shí)增加血清IgM和IgG濃度,減少腹瀉發(fā)生率(陳熠等,2009)。
2 BCAAs對(duì)豬免疫功能的影響
2.1提高母豬免疫力多年來(lái)的研究顯示,BCAAs對(duì)于母豬免疫功能具有重要的生理學(xué)意義。黃紅英等(2008)研究發(fā)現(xiàn),在哺乳母豬飼糧中添加BCAAs,可顯著提高仔豬血清中白蛋白、血糖水平及乳酸脫氫酶活性,明顯降低血清尿素氮含量,同時(shí)顯著增加仔豬血清IgG濃度,明顯減少腹瀉率,說(shuō)明BCAAs可通過(guò)母體效應(yīng),對(duì)提高仔豬的免疫力、抗貧血和促進(jìn)生長(zhǎng)等方面產(chǎn)生積極作用。而王勇(2010)在哺乳母豬日糧中添加適量纈氨酸時(shí)發(fā)現(xiàn),與對(duì)照組相比,處理組可提高母豬哺乳14 d和21 d血液中淋巴細(xì)胞和白細(xì)胞個(gè)數(shù),以及7 d血液中淋巴細(xì)胞個(gè)數(shù)(P<0.05),同時(shí)提高哺乳14 d血清中IgG和補(bǔ)體C3水平(P<0.05)以及21 d母豬血清中補(bǔ)體C3和補(bǔ)體C4含量(P<0.01);此外,還能提高母豬血清中IgM濃度,其中在哺乳7、14、21 d分別增加22.2%(P<0.01)、14%(P<0.05)和11.9%(P<0.05)。
2.2提高仔豬免疫力Petro等(1981)證實(shí),當(dāng)斷奶仔豬缺乏BCAAs時(shí),雖對(duì)細(xì)胞免疫無(wú)影響,但會(huì)導(dǎo)致合成特異性抗體的能力下降,并發(fā)現(xiàn)纈氨酸具有促進(jìn)骨骼T細(xì)胞轉(zhuǎn)化為成熟T細(xì)胞的作用(Petro等,1981)。黃紅英等(2007)報(bào)道,哺乳母豬日糧中添加BCAAs能通過(guò)增加仔豬血清IgG濃度改善仔豬機(jī)體免疫功能,進(jìn)而降低其腹瀉率。另有研究發(fā)現(xiàn),在斷奶仔豬日糧中添加0.27%的亮氨酸,與對(duì)照組相比可顯著提高其采食后1、2、3、4 h血漿免疫球蛋白A(IgA)和IgM的質(zhì)量濃度,但對(duì)IgG的質(zhì)量濃度無(wú)顯著影響(李奇等,2012)。
Ren等(2015)研究報(bào)道,在斷奶仔豬日糧中添加BCAAs,有利于小腸腸絨毛形態(tài)的保護(hù)與發(fā)育,并能增加十二指腸絨毛高度,減少上皮內(nèi)淋巴細(xì)胞數(shù)量,增加空腸IgA、IgM水平,以及回腸I-gA、IgG的水平,從而全面增強(qiáng)斷奶仔豬腸道免疫防御功能。
3 應(yīng)用BCAAs的注意事項(xiàng)
3.1適宜的添加量豬對(duì)BCAAs的生理需要量除受BCAAs間拮抗作用的程度,以及與其他營(yíng)養(yǎng)素的互作效應(yīng)影響外,還受到其自身生產(chǎn)性能、日糧能量水平及其生理狀況等因素的影響。Gatnau等(1995)研究指出,影響B(tài)CAAs及其轉(zhuǎn)氨基作用的中間產(chǎn)物對(duì)動(dòng)物免疫刺激調(diào)節(jié)效果的因素,主要來(lái)自于BCAAs的添加形式、添加量與適用動(dòng)物的種類等方面,如過(guò)量的亮氨酸(3.12%)對(duì)斷奶仔豬的生長(zhǎng)和免疫均將產(chǎn)生有害的負(fù)面影響;若在4%蛋白質(zhì)水平的飼料中添加過(guò)量亮氨酸,可使斷奶仔豬的免疫反應(yīng)受到嚴(yán)重抑制;添加適量的異亮氨酸或/和纈氨酸可大大減弱過(guò)量亮氨酸所造成的免疫抑制作用。該課題組還在以斷奶仔豬為研究對(duì)象的試驗(yàn)中發(fā)現(xiàn),向含l.12%亮氨酸的飼糧中分別添加1.12%亮氨酸、l.12%α-酮異己酸或0.4%β-羥基-β-甲基丁酸均對(duì)仔豬生長(zhǎng)和免疫功能無(wú)不良影響,但向含1.56%亮氨酸的飼糧中添加1.56%的亮氨酸則對(duì)仔豬生長(zhǎng)和免疫功能均有不利的影響(Gatnau等,1995)。
Richert等(1997a)研究報(bào)道,哺乳母豬對(duì)異亮氨酸的需要量為0.85%,該需要量比NRC(1988)標(biāo)準(zhǔn)量約高0.2%。Franco等(2013)發(fā)現(xiàn),妊娠母豬從早期到晚期對(duì)異亮氨酸的需要量是逐漸增加的。Htoo(2014)研究認(rèn)為,使18~40 kg仔豬增重達(dá)最大的回腸表觀可消化異亮氨酸需要量為日糧的0.51%,而飼料轉(zhuǎn)化率達(dá)最高的相應(yīng)值為0.55%;在亮氨酸不超量的基礎(chǔ)上,體重為10~ 22 kg與24~39 kg豬對(duì)異亮氨酸∶賴氨酸平均最佳標(biāo)準(zhǔn)回腸可消化(SID)比率分別約為51%與54%。哺乳母豬對(duì)BCAAs需要量的多少還與其哺仔數(shù)的數(shù)量有關(guān),一般情況下哺仔數(shù)多的母豬要比哺仔數(shù)少的母豬需要更高的BCAAs總需要量。Richert等(1997b)進(jìn)一步研究報(bào)道,當(dāng)飼糧BCAAs比例平衡時(shí),高產(chǎn)哺乳母豬對(duì)BCAAs的總需要量至少占飼糧的3.27%(每天203 g),其中異亮氨酸為0.85%、纈氨酸為1.07%、亮氨酸為1.35%;此外,飼糧中BCAAs含量不平衡時(shí)將增加哺乳母豬對(duì)其總的需要量。由此可見(jiàn),飼糧中BCAAs的含量及比例(或模式)是同等的重要,倘若過(guò)量添加BCAAs反而不利于豬免疫力的正常發(fā)揮。
此外,Mavromichalis等(1998)報(bào)道,仔豬在體重為10 kg階段采食低蛋白質(zhì)(13.5%)玉米-大豆粕日糧時(shí),纈氨酸對(duì)仔豬生產(chǎn)性能的限制程度與色氨酸、蘇氨酸或蛋氨酸相似,但其限制程度要高于異亮氨酸;結(jié)果還提示,10kg仔豬對(duì)纈氨酸的需要量可能高于NRC(1998)的推薦標(biāo)準(zhǔn)(0.79%)。綜合眾多研究發(fā)現(xiàn),在8~25kg體重階段,仔豬標(biāo)準(zhǔn)回腸可消化纈氨酸(SID)需要量最高可達(dá)到0.92%(Gaines等,2006),而最低僅為0.53%(Barea等,2009)。
綜上可知,豬對(duì)BCAAs的生理需要量及其組成模式除了與BCAAs間的拮抗效應(yīng)有關(guān)外,還受到豬的基因類型、飼糧能量水平與生理階段等因素的影響。表1概括了美國(guó)NRC(1998)標(biāo)準(zhǔn)中豬對(duì)BCAAs的推薦需要量及其模式,可為BCAAs在豬日糧中的應(yīng)用提供科學(xué)參考。
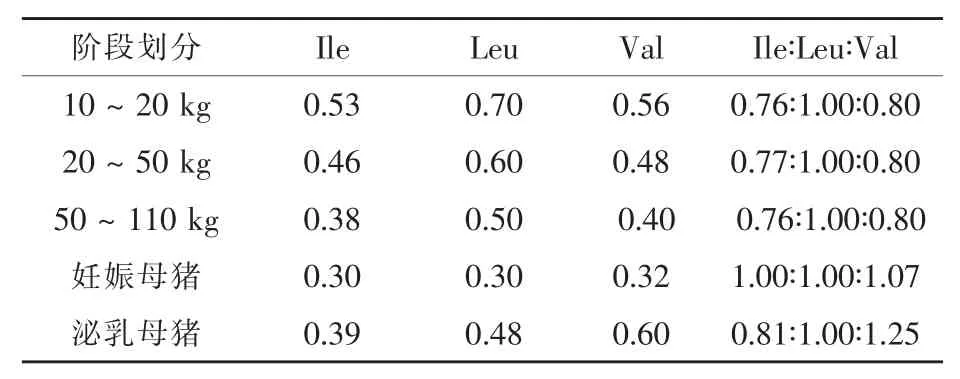
表1 美國(guó)NRC(1998)標(biāo)準(zhǔn)中豬對(duì)BCAAs的需要量及其模式%
3.2BCAAs的拮抗反應(yīng)BCAAs均具有相似的側(cè)鏈結(jié)構(gòu),故在小腸壁吸收轉(zhuǎn)運(yùn)與通過(guò)各種細(xì)胞膜時(shí)會(huì)相互競(jìng)爭(zhēng)轉(zhuǎn)運(yùn)體系,而且3種BCAAs在分解代謝過(guò)程中還會(huì)競(jìng)爭(zhēng)相同的降解酶。研究發(fā)現(xiàn),當(dāng)日糧中任一種BCAAs含量升高或過(guò)量或比例失衡時(shí),均會(huì)增加BCAAs轉(zhuǎn)氨酶的活力,促使其他BCAAs分解加劇,導(dǎo)致其他兩種BCAAs不平衡性加劇(代臘等,2012;Mello等,1970)。因此,三種BCAAs間存在一定的拮抗效應(yīng),并已得到了多種動(dòng)物試驗(yàn)的研究證實(shí)。Peter等(1988)報(bào)道,羔羊日糧中添加亮氨酸水平過(guò)高,動(dòng)物機(jī)體對(duì)異亮氨酸與纈氨酸的轉(zhuǎn)化效率將大幅度降低;同時(shí),隨著異亮氨酸的減少,亮氨酸的氧化將逐漸增加。此外研究還發(fā)現(xiàn),異亮氨酸過(guò)低可導(dǎo)致細(xì)胞間質(zhì)內(nèi)異亮氨酸含量減少,進(jìn)一步降低白蛋白與纖維蛋白的合成率(杜宗亮,2006)。
目前,以玉米為主要原料的基礎(chǔ)日糧中,亮氨酸水平偏高較為普遍,該配比容易導(dǎo)致異亮氨酸與纈氨酸相對(duì)性的缺乏。Taylor等(1977)研究證實(shí),將含0.38%異亮氨酸的生長(zhǎng)豬日糧中1.34%亮氨酸添加量提高到2.04%,可拮抗另外兩種BCAAs的正常功效,以致影響到生長(zhǎng)豬飼料轉(zhuǎn)化率、日增重與胴體品質(zhì)等生產(chǎn)性能;若將0.38%的異亮氨酸提高到0.45%,卻能顯著促進(jìn)豬的生長(zhǎng)(陳熠等,2008)。Gloaguen等(2012)研究也發(fā)現(xiàn),日糧中缺乏纈氨酸會(huì)抑制生長(zhǎng)豬的采食,而且攝入過(guò)多的亮氨酸會(huì)進(jìn)一步加劇以上采食抑制作用;該結(jié)果進(jìn)一步印證過(guò)量的亮氨酸會(huì)增加另外兩種BCAAs的分解代謝,進(jìn)而加重由纈氨酸和異亮氨酸缺乏所引起的生產(chǎn)抑制效應(yīng)。王彬等(2012)試驗(yàn)也指出,在日糧蛋白質(zhì)不足(9%酪蛋白)時(shí)添加3%的高水平亮氨酸可抑制老鼠生長(zhǎng),但能通過(guò)添加異亮氨酸得到部分緩解;而在蛋白質(zhì)充足(18%酪蛋白)日糧中,添加3%的亮氨酸并不引起老鼠生長(zhǎng)抑制,同時(shí)還未發(fā)現(xiàn)高水平異亮氨酸和纈氨酸的拮抗效應(yīng)。可見(jiàn),3種BCAAs不僅相互存在拮抗作用,而且在人類與動(dòng)物體內(nèi)主要由亮氨酸所引起。
另有文獻(xiàn)表明,BCAAs在生化代謝過(guò)程中所產(chǎn)生的拮抗作用,還會(huì)進(jìn)一步影響到動(dòng)物機(jī)體免疫功能的正常發(fā)揮(Beisel,1982)。
4 結(jié)語(yǔ)
BCAAs作為氮的載體,是豬的必需氨基酸,具有構(gòu)成肌肉蛋白、刺激胰島素分泌、促進(jìn)蛋白合成等多方面的生物學(xué)功能,特別是對(duì)豬免疫防御機(jī)能發(fā)揮著調(diào)節(jié)作用。但在豬日糧中添加BCAAs時(shí),要十分注意其不當(dāng)應(yīng)用所帶來(lái)的負(fù)面影響。如豬飼料中某種BCAAs含量過(guò)高或比例不當(dāng)都會(huì)引起B(yǎng)CAAs之間的拮抗現(xiàn)象和免疫抑制作用。相信隨著畜牧科學(xué)研究的不斷深入,養(yǎng)豬業(yè)中應(yīng)用BCAAs會(huì)日漸變得更加科學(xué)化與合理化。
[1]陳熠,賀建華,彭藝,等.豬的支鏈氨基酸營(yíng)養(yǎng)[J].飼料廣角,2008,9:23~25.
[2]陳熠,彭藝,賀建華,等.添加纈氨酸對(duì)泌乳母豬血清和乳中生化免疫指標(biāo)及激素水平的影響[J].中國(guó)飼料,2009,17:15~20.
[3]代臘,鄒曉庭.纈氨酸在畜禽生產(chǎn)中的應(yīng)用[J].飼料與畜牧,2012,1:20~23.
[4]杜宗亮.支鏈氨基酸在哺乳母豬營(yíng)養(yǎng)中的研究進(jìn)展[J].飼料工業(yè),2006,27(21):19~22.
[5]宮碧霜,任曼,靳二輝,等.異亮氨酸對(duì)大鼠血清抗氧化水平及脾臟組織結(jié)構(gòu)的影響[J].安徽科技學(xué)院學(xué)報(bào),2015,6:1~5.
[6]黃紅英,賀建華,范志勇,等.添加纈氨酸和異亮氨酸對(duì)哺乳母豬及其仔豬生產(chǎn)性能的影響[J].動(dòng)物營(yíng)養(yǎng)學(xué)報(bào),2008,3:281~287.
[7]黃紅英,賀建華,范志勇,等.母豬日糧中支鏈氨基酸水平對(duì)仔豬血液生化指標(biāo)和部分免疫指標(biāo)的影響[J].飼料工業(yè).2007,28(21):24~6.
[8]李奇,王彬,袁珍虎.亮氨酸對(duì)斷奶仔豬生長(zhǎng)和免疫性能的影響[J].飼料研究,2012,8:3~5.
[9]劉春生,張大鵬,劉文寬,等.支鏈氨基酸及其在泌乳母豬營(yíng)養(yǎng)中的研究現(xiàn)狀[J].飼料工業(yè),2006,1:25~26.
[10]靳繼德,鄭子新,王鋼樂(lè),等.Ala-Gln和BCAA對(duì)創(chuàng)傷大鼠營(yíng)養(yǎng)支持作用的實(shí)驗(yàn)研究[J].中國(guó)臨床營(yíng)養(yǎng)雜志,2001,1:19~23.
[11]陶義訓(xùn).免疫測(cè)定進(jìn)展[J].上海免疫學(xué)雜志,1999,2:2~5.
[12]王彬,李奇.亮氨酸的代謝及營(yíng)養(yǎng)生理作用研究進(jìn)展[J].飼料研究,2012,1:14~16.
[13]王勇.纈氨酸對(duì)高產(chǎn)哺乳母豬生產(chǎn)性能、免疫機(jī)能及氮利用率的影響研究:[碩士學(xué)位論文][D].杭州:浙江大學(xué),2010.
[14]徐琪壽.氨基酸藥理學(xué)研究進(jìn)展[J].氨基酸和生物資源,1996,1:30~32.
[15]Aschkenasy A.Prevention of the immunodepressive effects of excess dietary leucine by isoleucine and valine in the rat[J].J Nutr,1979,109(7):1214~22.
[16]Belokrylov G A,Molchanova I V,Sorochinskaya E I.Immunomodulatory properties of certain amino acids influence the immunostimulating properties of specific peptides[J].Int J Immunopharmacol,1990,12(8):841~5.
[17]Beisel WR.Single nutrients and immunity[J].Am J Clin Nutr.1982,35(2):417~68.
[18]Barea R,Brossard L,Le Floc'h N,et al.The standardized ileal digestible valine-to-lysine requirement ratio is at least seventy percent in postweaned piglets[J].J Anim Sci,2009,87(3):935~947.
[19]Calder P C.Branched-chain amino acids:metabolism,physiological function,and application:session III,branched-chain amino acids and immunity[J]. American Society for Nutrition,J Nutr,2006,136:288S~93S.
[20]Chevalier P,Aschkenasy A.Hematological and immunological effects of excess dietary leucine in the young rat[J].Am J Clin Nutr,1977,30:1645~54.
[21]Dunshea F R,Bauman D E,Nugent E A,et al.Hyperinsulinaemia,supplemental protein and branched-chain amino acids when combined can increase milk protein yield in lactating sows[J].British Journal of Nutrition,2005,93(3):325~332.
[22]Franco D J,Josephson J K,Moehn S,et al.Isoleucine requirement of pregnant sows[J].J Anim Sci,2013,91(8):3859~66.
[23]Gaines A M,Srichana P,Ratliff B W,et al.Evaluation of the true ileal digestible(TID)valine requirement of 8 to 20 kg pigs[J].J Anim Sci,2006,84(1):284.
[24]Gatnau R,Zimmerman D R,Nissen S L,et al.Effects of excess dietary leucine and leucine catabolites on growth and immune responses in weanling pigs[J].J Anim Sci,1995,73(1):159~65.
[25]Gloaguen M,Le Floc'h N,Corrent E,et al.Providing a diet deficient in valine but with excess leucine results in a rapid decrease in feed intake and modifies the postprandial plasma amino acid and α-keto acid concentrations in pigs[J].J Anim Sci,2012,90(9):3135~42.
[26]Harris R A,Joshi M,Jeoung N H,et al.Overview of the molecular and biochemical basis of branched-chain amino acid catabolism[J].J Nutr,2005,135(6):1527S~30S.
[27]Htoo J K,Zhu C L,Huber L,et al.Determining the optimal isoleucine:lysine ratio for ten-to twenty-two-kilogram and twenty-four-to thirtynine-kilogram pigs fed diets containing nonexcess levels of leucine[J].J Anim Sci,2014,92(8):3482~90.
[28]Mavromichalis I L,Webel D M,Emm ert J L,et al.Limiting order of amino acids in a low-protein corn-soybean meal-whey-based diet for nursery pigs[J].J Anim Sci,1998,76(11):2833~7.
[29]Mello J P F,Lewis D.Amino acid interactions in chick nutrition[J].British Poultry Science,1970,11(3):367~385.
[30]Negro M,Giardina S,Marzani B,et al.Branched-chain amino acid supplementation does not enhance athletic performance but affects muscle recovery and the immune system[J].J Sports Med Phys Fitness,2008,48(3):347~51.
[31]NRC.Nutrient Requirements of Swine(10thED.).National Academy Press,Washington,DC.1998.
[32]Papet I,Breuille D,Glomot F,et al.Nutritional and metabolic effects of dietary leucine excess in preruminant lamb[J].J Nutr,1988,118(4):450~5.
[33]Petro T M,Bhattacharjee J K.Effect of dietary essential amino acid limitations upon susceptibility to Salmonella typhimurium and the effect upon humoral and cellular immune response in mice[J].Infect Immun,1981,32:251~9.
[34]Ren M,Zhang S H,Zeng X F,et al.Branched-chain Amino Acids are Beneficial to Maintain Growth Performance and Intestinal Immune-related Function in Weaned Piglets Fed Protein Restricted Diet[J].Asian-Australas J Anim Sci,2015,28(12):1742~50.
[35]Richert B T,Goodband R D,Tokach M D,et al.Increasing valine,isoleucine,and total branched-chain amino acids for lactating sows[J].J Anim Sci,1997a,75(8):2117~28.
[36]Richert B T,Tokach M D,Goodband R D,et al.The effect of dietary lysine and valine fed during lactation on sow and litter performance[J].J Anim Sci,1997b,75(7):1853~60.
[37]Taylor S J,Cole D J,Lewis D.An interaction of leucine,isoleucine and valine in the diet of the growing pig[J].Proc Nutr Soc,1977,36(1):36A.■
Branched-chain amino acids regarding as essential amino acids for pig,have many important biological functions,such as providing nutrition,regulating immune,promoting secretion and participating in metabolism.In this study,the latest research results of the branched-chain amino acids in pig with immune function were reviewed,and the mechanism of branched-chain amino acids in immunological functions was further discussed,to provide scientific references for the related research.
branched-chain amino acids;pig;immune function;mechanism
10.15906/j.cnki.cn11-2975/s.20161603
S816.7
A
1004-3314(2016)16-0010-04
國(guó)家自然科學(xué)基金項(xiàng)目(31372394);香港銘源基金(2011-165);廣東省自然科學(xué)基金(2014A030313699、2015A030310094)
猜你喜歡
中華詩(shī)詞(2022年6期)2022-12-31 06:41:24
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國(guó)語(yǔ)大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
中國(guó)科技論壇(2017年7期)2017-07-25 08:49:53
汽車工程學(xué)報(bào)(2017年2期)2017-07-05 08:13:02
媽媽寶寶(2017年2期)2017-02-21 01:21:24
國(guó)際漢語(yǔ)學(xué)報(bào)(2016年1期)2017-01-20 08:21:20
