基于轉(zhuǎn)錄組信息的艾納香牻牛兒基牻牛兒基焦磷酸合成酶基因(BbGGPS)的克隆及序列分析
2016-11-09 06:16:21夏奇峰劉紅昌官玲亮龐玉新
山地農(nóng)業(yè)生物學(xué)報(bào) 2016年4期
關(guān)鍵詞:分析
夏奇峰,趙 致,劉紅昌,官玲亮,龐玉新
(1.貴州大學(xué) 生命科學(xué)學(xué)院,貴州 貴陽 550025;2.貴州大學(xué) 農(nóng)學(xué)院貴州省中藥材重點(diǎn)實(shí)驗(yàn)室,貴州 貴陽 550025;3.中國(guó)熱帶農(nóng)業(yè)科學(xué)院熱帶作物品種資源研究所/農(nóng)業(yè)部華南作物基因資源與種質(zhì)創(chuàng)制重點(diǎn)開放實(shí)驗(yàn)室/海南省艾納香工程技術(shù)研究中心,海南 儋州 571737)
?
基于轉(zhuǎn)錄組信息的艾納香牻牛兒基牻牛兒基焦磷酸合成酶基因(BbGGPS)的克隆及序列分析
夏奇峰1,趙致2*,劉紅昌2,官玲亮3,龐玉新3
(1.貴州大學(xué) 生命科學(xué)學(xué)院,貴州 貴陽 550025;2.貴州大學(xué) 農(nóng)學(xué)院貴州省中藥材重點(diǎn)實(shí)驗(yàn)室,貴州 貴陽 550025;3.中國(guó)熱帶農(nóng)業(yè)科學(xué)院熱帶作物品種資源研究所/農(nóng)業(yè)部華南作物基因資源與種質(zhì)創(chuàng)制重點(diǎn)開放實(shí)驗(yàn)室/海南省艾納香工程技術(shù)研究中心,海南 儋州 571737)
采用RT-PCR和RACE(Rapid amplification of cDNA ends)技術(shù),從艾納香(Blumea balsamifera L·DC)的葉片中克隆到二萜化合物合成的關(guān)鍵酶牻牛兒基牻牛兒基焦磷酸合成酶(BbGGPS)基因。結(jié)果顯示:BbGGPS基因的cDNA全長(zhǎng)1475 bp,包含開放閱讀框(ORF)1002 bp,編碼334個(gè)氨基酸;亞細(xì)胞結(jié)構(gòu)定位于葉綠體,既非膜蛋白也非分泌性蛋白。疏水性分析顯示,BbGGPS是親水性蛋白。同源性比對(duì)結(jié)果顯示,BbGGPS蛋白與其他植物中GGPS蛋白具有高度的相似性。系統(tǒng)發(fā)育分析表明,所有序列被聚為5大類,BbGGPS與菊科植物刺菜薊聚(Cynara cardunculus var)為一類,表明與其親緣關(guān)系最近。
艾納香;牻牛兒基牻牛兒基焦磷酸合成酶;氨基酸序列;系統(tǒng)發(fā)育分析
艾納香(Blumea balsamifera L.DC.)為菊科艾納香屬多年生木質(zhì)草本植物,最早記載于公元741年(唐開元二十九年)陳藏器所編著 《本草拾遺》,此后宋代劉翰等編著《開寶本草》(公元 973 -974 年)也曾記載[1]。艾納香以根、嫩枝、葉入藥,其性微溫,味辛、微苦,具有祛風(fēng)消腫、活血散瘀之功效,可用于治療感冒、風(fēng)濕性關(guān)節(jié)炎、產(chǎn)后風(fēng)痛、痛經(jīng)、 外用跌打損傷、瘡癤痛腫、濕疹皮炎。另外,艾納香還具有抗氧化、抗癌和抗病毒的作用[2]。主要分布于我國(guó)海南、貴州、廣西、廣東、云南、臺(tái)灣等省。在黎族、苗族、壯族等少數(shù)民族地區(qū)有著悠久的藥用歷史,是一種重要的民間藥物。隨著現(xiàn)代藥學(xué)、分析化學(xué)等學(xué)科的發(fā)展,艾納香所含有成分的分析已有較多文獻(xiàn)報(bào)導(dǎo)[3-7]。研究表明,艾納香中所含成分主要是黃酮類物質(zhì)和揮發(fā)油。黃酮類化合物主要包括黃酮和黃酮苷類物質(zhì),揮發(fā)油主要包括一系列萜類化合物,從單體組成上劃分,可以分為單萜、倍半萜、二萜和三萜等,如L-龍腦、β-石竹烯、γ-桉葉油醇、樟腦、芳樟醇等。
二萜類化合物藥理活性廣泛,現(xiàn)代藥理學(xué)研究表面二萜類化合物具有抗腫瘤、抗菌、抗病毒、抑制超敏反應(yīng)等作用[8-9]。牻牛兒基牻牛兒基焦磷酸合成酶(geranylgeranyl pyrophosphate synthase,GGPS)在二萜生物合成途徑中起著關(guān)鍵性的作用,GGPS催化法尼基焦磷酸(FPP)與異戊烯焦磷酸(IPP)形成的牻牛兒基牻牛兒基焦磷酸(GGPP),它是二萜生物合成的共同前體[10]。在GGPP的結(jié)構(gòu)基礎(chǔ)上,經(jīng)過環(huán)化異構(gòu)形成各種陽離子中間產(chǎn)物,之后進(jìn)一步環(huán)化形成一系列二萜化合物。
對(duì)GGPS基因的研究已有較多的文獻(xiàn)報(bào)導(dǎo),如:馬利剛等[11]成功克隆到北葶藶子(Lepidium apetalum)的GGPS基因(LaGGPS),化文平等[12]克隆了丹參(Salvia miltiorrhiza Bunge)的GGPS基因(SmGGPS),李翠萍等[13]克隆了番茄(Solanum lycopersicon L)的GGPS 2基因(SlGGPS2),閔丹丹等[14]克隆了三七(Notoginseng Radix Et Rhizoma)的GGPS基因。目前還未見有關(guān)艾納香GGPS基因(BbGGPS)的研究報(bào)導(dǎo)。本課題組基于艾納香的轉(zhuǎn)錄組測(cè)序信息,以預(yù)測(cè)所得的一條可能的GGPS基因序列作為模板,設(shè)計(jì)特異引物克隆了一條BbGGPS基因,并進(jìn)行了基因序列與其編碼的蛋白質(zhì)序列的生物信息學(xué)分析。艾納香在分子生物學(xué)方面的研究尚未起步,對(duì)艾納香BbGGPS基因進(jìn)行研究,有望從分子水平揭示艾納香活性成分代謝途徑和調(diào)控機(jī)制,為提高艾納香活性成分的含量奠定理論基礎(chǔ)。
1 材料與方法
1.1材料
1.1.1植物材料
艾納香(Blumea balsamifera L·DC)栽培于海南省儋州市,中國(guó)熱帶農(nóng)業(yè)科學(xué)院熱帶作物品種資源研究所南藥種質(zhì)資源圃,試驗(yàn)材料為艾納香長(zhǎng)勢(shì)良好的葉片。
1.1.2試劑
總RNA提取試劑盒(Plant RNA Kit)、反轉(zhuǎn)錄試劑盒、載體連接試劑盒、異丙基硫代半乳糖苷(IPTG)、X-gal、多功能 DNA 純化回收試劑盒均購(gòu)自廣州飛揚(yáng)生物工程有限公司。引物合成與測(cè)序均由上海美吉(Majorbio)生物工程技術(shù)服務(wù)有限公司完成。
1.2方法
1.2.1總RNA提取及 BbGGPS全長(zhǎng)cDNA的克隆
按照Plant RNA kit試劑盒說明書,提取艾納香葉片的總RNA,采用RT-PCR合成cDNA第一鏈。通過從本課題組前期對(duì)艾納香花和葉片轉(zhuǎn)錄組的信息,挖掘艾納香中GGPS相關(guān)基因的序列片段,設(shè)計(jì)特異引物擴(kuò)增cDNA。正向引物Pf:5’-GTAATTTGCA TCTCATCTGA TATTCG-3’、反向引物Pr:5’-CAAAACGCCA AAACCCAAG-3’,以 cDNA為模板進(jìn)行PCR擴(kuò)增。反應(yīng)體系:PCR反應(yīng)總體積為50 μL,1.5 mmol/L MgCl2,4種dNTP各200 μmol/L,引物各150 ng,1.5 U Taq plus DNA polymerase(高保真),100 ng cDNA。反應(yīng)條件:94℃、5 min;94℃、30 s,53℃、30 s,72℃、60 s,30個(gè)循環(huán);72℃后延伸10 min。并進(jìn)行克隆、測(cè)序及序列分析。
1.2.2BbGGPS 基因的測(cè)序與分析
BbGGPSPCR產(chǎn)物經(jīng)1.0%的瓊脂糖凝膠電泳檢測(cè),用膠回收試劑盒對(duì)目的片段進(jìn)行純化,純化后將其連接到pEASY-Blunt cloning vector載體上,轉(zhuǎn)化 E.coli Trans1-T1 感受態(tài)細(xì)胞,涂布于添加氨芐青霉素、IPTG、X-gal 的 LB 平板上,37℃培養(yǎng)箱中培養(yǎng)過夜,隨機(jī)挑取陽性克隆,使用M13Pf/Pr引物,經(jīng)PCR檢測(cè)后擴(kuò)菌培養(yǎng),再送樣進(jìn)行測(cè)序,獲得重組載體pEASY-Blunt-BbGGPS。
1.3艾納香BbGGPS 基因的生物信息學(xué)分析
使用DNAMAN軟件分析BbGGPS基因的cDNA序列,找到對(duì)應(yīng)的開放閱讀框(ORF)以及對(duì)應(yīng)編碼的氨基酸序列;利用DNAMAN軟件的多序列比對(duì)功能,將BbGGPS氨基酸序列與NCBI中BLAST檢索到的高同源性序列進(jìn)行比對(duì),對(duì)保守區(qū)域進(jìn)行考察;使用BioEdit等軟件對(duì)BbGGPS氨基酸序列的理化性質(zhì)進(jìn)行分析,并使用Swiss-model工具對(duì)BbGGPS的三維結(jié)構(gòu)在線進(jìn)行預(yù)測(cè);最后利用Mega 6.0將BbGGPS氨基酸序列與從NCBI中進(jìn)行BLAST比對(duì)搜索得到的序列進(jìn)行系統(tǒng)進(jìn)化分析。
2 結(jié)果與分析
2.1艾納香BbGGPS基因的克隆
以艾納香新鮮葉片的cDNA為模板,基于轉(zhuǎn)錄組測(cè)序預(yù)測(cè)的GGPS基因序列設(shè)計(jì)一對(duì)特異引物擴(kuò)增出的PCR產(chǎn)物在1450 bp左右,與預(yù)期片段大小一致,電泳結(jié)果見圖1。將目的條帶回收,構(gòu)建重組載體pEASY-Blunt-BbGGPS轉(zhuǎn)化大腸桿菌,將PCR鑒定為陽性的克隆送樣測(cè)序。
測(cè)序之后得到1475 bp的cDNA序列,通過在線ORF Finder 分析,發(fā)現(xiàn)其中包含一個(gè)1002 bp的ORF,編碼333個(gè)氨基酸,堿基序列見圖2。將此氨基酸序列在NCBI blastp對(duì)比nr數(shù)據(jù)庫(kù),比對(duì)結(jié)果表明該序列與菊科植物刺菜薊(Cynara cardunculus var.scolymus)的聚戊烯合成酶序列相似性最高,為86%;其次與地黃(Rehmannia glutinosa)的GGPS序列相似性高達(dá)78%。從而可以推測(cè),克隆所得的cDNA序列編碼GGPS蛋白,將此基因命名為BbGGPS。
2.2BbGGPS氨基酸序列分析
2.2.1理化性質(zhì)分析
通過BbGGPScDNA序列預(yù)測(cè)得到其編碼的氨基酸序列,使用 Prot Param tool工具分析其理化性質(zhì)[11-14],選擇軟件默認(rèn)參數(shù)。結(jié)果顯示BbGGPS肽鏈包含有334個(gè)氨基酸殘基,分子量為36710.9,等電點(diǎn)(pI)為5.32(圖3)。氨基酸的種類分布如 圖4 所示,該肽鏈總共含有20種氨基酸,包括天冬酰胺和谷氨酰胺。含量最多的氨基酸種類為亮氨酸和纈氨酸,其次是亮氨酸和谷氨酸。而半胱氨酸、組氨酸的含量則相對(duì)較少,色氨酸僅有一個(gè)。帶負(fù)電荷氨基酸殘基總數(shù)(Asp+Glu)為45個(gè),帶正電荷氨基酸殘基總數(shù)(Arg+Lys)為36個(gè)。總體而言,非極性氨基酸的含量相對(duì)較高。不穩(wěn)定系數(shù)為37.92,屬于穩(wěn)定蛋白。
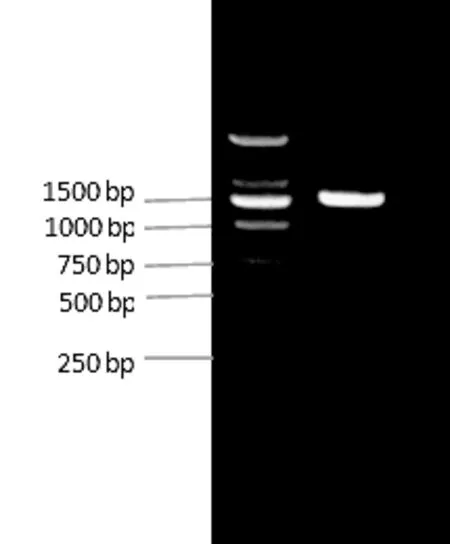
圖1 從艾納香中克隆所得的GGPS基因片段Fig.1 The GGPS gene clone fragment from Blumea balsamifera leaves

圖2 艾納香葉片中BbGGPS基因的cDNA序列(圖中框內(nèi)分別是起始密碼子與終止密碼子)Fig.2 cDNA sequence ofBbGGPS in Blumea balsamifera leaves(The shadows are initiation codon and termination codon)

圖3 BbGGPS氨基酸序列Fig.3 Amino acid sequence of BbGGPS
2.2.2BbGGPS跨膜區(qū)、信號(hào)肽、亞細(xì)胞定位分析
使用TMHMM Sever v.2.0預(yù)測(cè)BbGGPS的跨膜區(qū)域[11-14],輸出格式選擇為默認(rèn)的圖形化顯示(Extensive,with graphics)。分析結(jié)果表明,BbGGPS無跨膜區(qū),屬于膜外在蛋白;利用ExPASy SignalP 4.0 Serve分析BbGGPS蛋白,并沒有發(fā)現(xiàn)信號(hào)肽,表明該蛋白為非分泌蛋白;用WoLFPSORT工具對(duì)BbGGPS進(jìn)行亞細(xì)胞結(jié)構(gòu)定位預(yù)測(cè),預(yù)測(cè)結(jié)果表明,BbGGPS最可能定位于葉綠體。
2.2.3疏水性分析
使用ExPASy-ProtScale工具中的Kyte &Doolittle法(K-D法)對(duì)艾納香GGPS蛋白質(zhì)的疏水性進(jìn)行在線分析[11-14],窗口參數(shù)(Window size)設(shè)定為11。分析結(jié)果表明,BbGGPS親水性略大于疏水性,BbGGPS屬于親水蛋白。在第100~150個(gè)氨基酸殘基之間,有一個(gè)強(qiáng)親水區(qū)域,大約在第270個(gè)氨基酸殘基處,還有一個(gè)很強(qiáng)的親水區(qū)(圖5)。

圖4 BbGGPS的氨基酸組成Fig.4 Amino acid number of BbGGPS
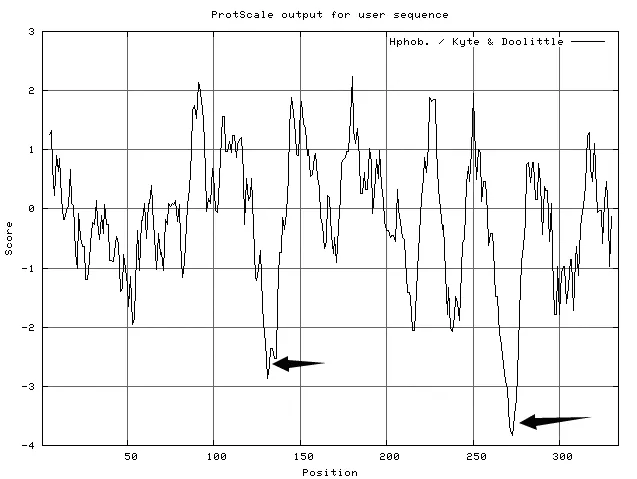
圖5 ProtScale分析BbGGPS疏水性的結(jié)果(圖中箭頭所指位置為強(qiáng)親水區(qū))Fig.5 Hydrophobic analysis of BbGGPS(Strong hydrophilic regions are marked in the picture)
2.2.4蛋白質(zhì)二級(jí)結(jié)構(gòu)預(yù)測(cè)和三維建模
利用SSpro 4.0(http://download.igb.uci.edu/sspro4.html)對(duì)BbGGPS的二級(jí)結(jié)構(gòu)進(jìn)行在線分析[11-14]。結(jié)果表明該蛋白二級(jí)結(jié)構(gòu)中α-螺旋(H)占 64.8%,β-折疊(E)占 1.7%,無規(guī)則卷曲(C)占 33.5%。
使用Swiss-model工具對(duì)BbGGPS的三維結(jié)構(gòu)在線進(jìn)行預(yù)測(cè)(圖6),不使用指定模版,默認(rèn)原始參數(shù)[11-14]。從圖中可以看到,BbGGPS的三維結(jié)構(gòu)主要由α-螺旋構(gòu)成,其次是無規(guī)則卷曲。β-折疊只占有很少的一部分,三維結(jié)構(gòu)預(yù)測(cè)的結(jié)果與蛋白質(zhì)二級(jí)結(jié)構(gòu)預(yù)測(cè)相吻合。分析結(jié)果表明,BbGGPS屬于α-螺旋型蛋白。其三維結(jié)構(gòu)的“口袋”區(qū)域,可能是BbGGPS起催化作用的活性中心所在。
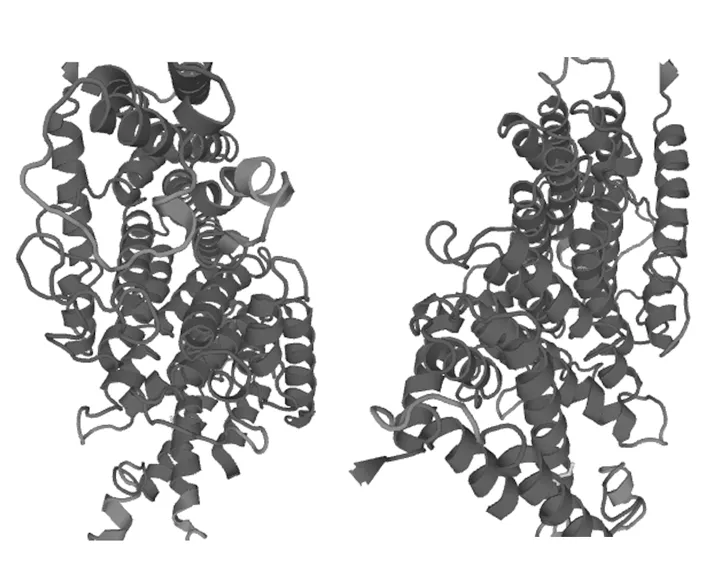
圖6 BbGGPS三維結(jié)構(gòu)預(yù)測(cè)Fig.6 Three dimensional structure prediction of BbGGPS
2.3多序列比對(duì)與系統(tǒng)進(jìn)化分析
2.3.1多序列比對(duì)
使用NCBI的BLAST工具,將BbGGPS序列在蛋白質(zhì)數(shù)據(jù)庫(kù)中進(jìn)行比對(duì)搜索。BbGGPS在蛋白質(zhì)數(shù)據(jù)庫(kù)中比對(duì)上不同物種的19條蛋白質(zhì)序列,其中刺菜薊(Cynara cardunculus var.scolymus)的聚戊烯合成酶序列相似性最高,為86%;其次與地黃(Rehmannia glutinosa)的GGPS序列相似性高達(dá)78%。利用 DNAMAN 將BbGGPS氨基酸序列與從NCBI 中挑選的部分同源性較高的已知序列進(jìn)行多序列比對(duì),結(jié)果表明BbGGPS蛋白與其他植物中GGPS蛋白具有高度的同源性(圖7)。


圖7 BbGGPS 多序列比對(duì)結(jié)果Fig.7 Multiple sequence alignment of BbGGPS
2.3.2系統(tǒng)進(jìn)化分析
利用Mega 6.0軟件將BbGGPS氨基酸序列與從NCBI中BLAST比對(duì)搜索得到的高分值序列(表1)進(jìn)行系統(tǒng)進(jìn)化分析,使用除權(quán)配對(duì)法(UPGMA)構(gòu)建系統(tǒng)進(jìn)化樹,用差異位點(diǎn)距離(P-distance)估計(jì)進(jìn)化距離。所構(gòu)建進(jìn)化樹的置信度顯著性檢驗(yàn)使用自助法(Bootstrap method)。由結(jié)果可知位于系統(tǒng)發(fā)育樹最下方的田七(Panax notoginseng,PnGGPS)、毛果楊(Populus trichocarpa,PtGGPS)、菜豆(Phaseolus vulgaris,PvGGPS)成為一個(gè)獨(dú)立的初級(jí)分支,表明這一個(gè)分支的物種與其余物種之間親緣關(guān)系較遠(yuǎn);擬南芥(Arabidopsis thaliana,AtGGPS)在進(jìn)化樹中作為樹根,表明擬南芥相對(duì)進(jìn)化樹中的其他物種進(jìn)化時(shí)序最早,進(jìn)化地位最原始。艾納香(Blumea balsamifera,BbGGPS)、刺菜薊(Cynara cardunculus var,CvGGPS)同屬于菊科植物,位于一個(gè)分支上,表明其親緣關(guān)系較近(圖8),這與形態(tài)學(xué)植物分類結(jié)果相似,表明GGPS基因可為植物分類方面的分析或研究提供一定的佐證。
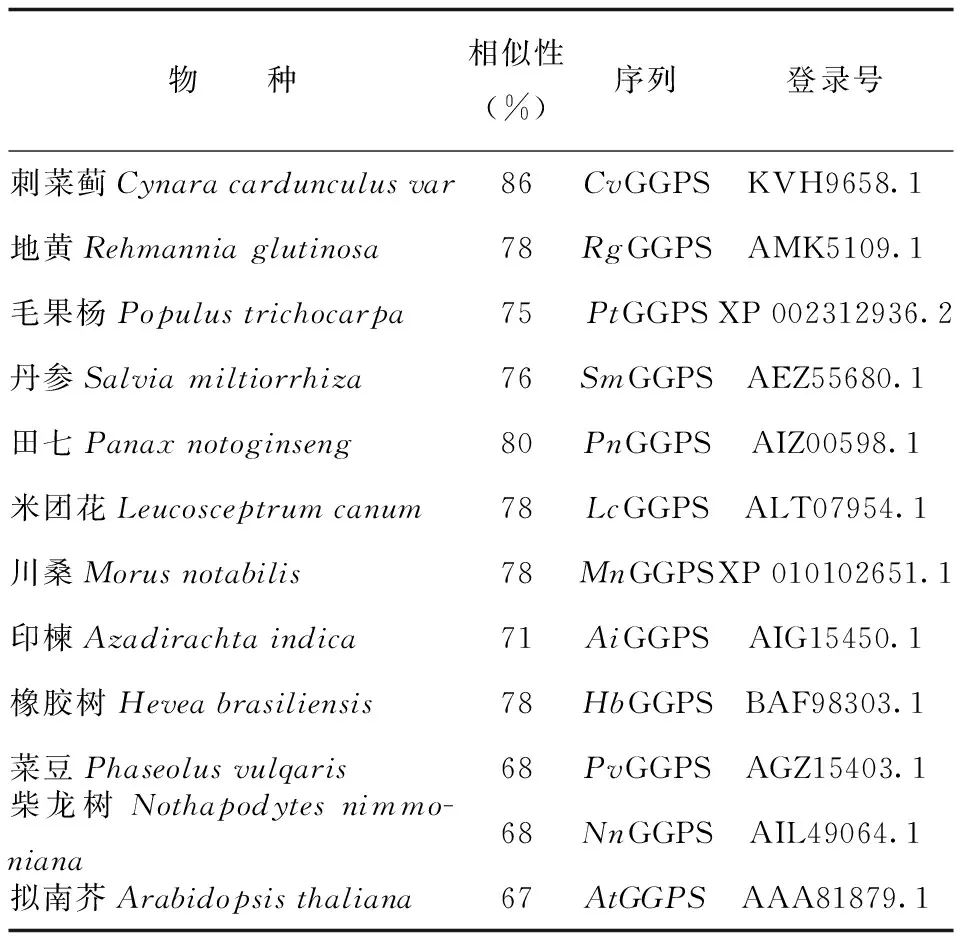
表1 BLAST比對(duì)高同源性GGPS序列
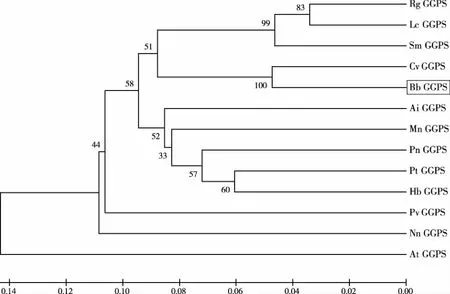
圖8 Mega對(duì)BbGGPS系統(tǒng)進(jìn)化分析結(jié)果(艾納香BbGGPS序列用方框標(biāo)出)Fig.8 system evolution analysis of BbGGPS(Gene sequence of BbGGPS is marked in frame)
3 結(jié)論與討論
本研究基于艾納香葉片的轉(zhuǎn)錄組數(shù)據(jù)信息,根據(jù)挖掘到的GGPS基因的序列片段,設(shè)計(jì)特異性引物成功從艾納香總RNA中擴(kuò)增BbGGPS的全長(zhǎng)cDNA序列。將擴(kuò)增得到的cDNA通過克隆、測(cè)序驗(yàn)證,擴(kuò)增得到的cDNA序列正確。
萜類化合物在植物體中普遍存在,有研究表明,許多高等植物中的藥用活性成分均屬于萜類化合物。萜類化合物包括單萜、二萜、倍半萜等。目前關(guān)于艾納香中二萜化合物的成分分析與合成機(jī)制等方面均未見文獻(xiàn)報(bào)道,尚屬空白。艾納香GGPS是二萜類物質(zhì)合成途徑中的關(guān)鍵酶,在艾納香二萜合成機(jī)制的研究中具有重要意義。GGPS屬于蛋白超家族,在高等植物不同物種之間具有較高的保守性。高等植物不同物種GGPS研究已有較多的文獻(xiàn)報(bào)導(dǎo),不論是GGPS cDNA序列長(zhǎng)度或是編碼的GGPS氨基酸個(gè)數(shù)都有較大的相似性。例如:北葶藶子GGPS的cDNA全長(zhǎng)1146 bp,編碼381個(gè)氨基酸[11];三七GGPS的cDNA全長(zhǎng)1203 bp,編碼344個(gè)氨基酸[14]。GGPS基因具有的保守結(jié)構(gòu)域主要是聚丙烯合成酶(polyprenylsynthetase);萜類合成酶(terpenoid synthase);聚丙烯相關(guān)合成酶(polyprenyl synthetase-related)。使用DNAMAN軟件,將艾納香BbGGPS與同源性較高的多條序列進(jìn)行比對(duì),發(fā)現(xiàn)BbGGPS氨基酸序列存在多個(gè)保守結(jié)構(gòu)域,與前人對(duì)其他物種的GGPS氨基酸序列研究成果相一致,證明BbGGPS同屬于GGPS蛋白超家族中的一員。GGPS在不同高等植物中的進(jìn)化上具有保守性,因此GGPS氨基酸序列的局部差異可以作為物種進(jìn)化時(shí)序的證據(jù)之一。
在本次研究中,通過不同物種間GGPS的氨基酸位點(diǎn)差異構(gòu)建了系統(tǒng)進(jìn)化樹,結(jié)果表明艾納香與菊科植物刺菜薊親緣關(guān)系最近,BbGGPS序列的系統(tǒng)發(fā)育分析結(jié)果與艾納香已有的種屬分類相一致,進(jìn)一步說明本次分析的結(jié)果是具有較高真實(shí)性和可信度的。植物中通常含有數(shù)目不等的多個(gè)GGPS基因,不同的GGPS基因亞細(xì)胞定位可能不同,表達(dá)模式也不盡相同,在不同的代謝途徑中起作用。化文平等已經(jīng)從丹參中克隆到了三條GGPS基因,實(shí)時(shí)定量PCR表明丹參的GGPS基因在不同發(fā)育時(shí)期不同器官中表達(dá)差異顯著[12]。本次測(cè)序得到的艾納香葉片轉(zhuǎn)錄組數(shù)據(jù)中也有多個(gè)不同序列的GGPS基因,有些基因序列并不完整,有待將來通過RACE技術(shù)獲取全長(zhǎng)序列之后進(jìn)一步研究其生物學(xué)功能。本實(shí)驗(yàn)克隆得到了艾納香二萜化合物合成的關(guān)鍵酶基因 GGPS,并對(duì)所得序列進(jìn)行了比較詳細(xì)的生物信息學(xué)分析,以期從分子生物學(xué)的角度揭示艾納香二萜生物合成的內(nèi)在機(jī)制,為艾納香的后續(xù)深入研究奠定了基礎(chǔ)。
[1]袁媛,龐玉新,王文全,等.艾納香屬系統(tǒng)分類研究進(jìn)展[J].熱帶農(nóng)業(yè)科學(xué),2011,31(3):81-86.
[2]官玲亮,龐玉新,王丹,等.中國(guó)民族特色藥材艾納香研究進(jìn)展[J].植物遺傳資源學(xué)報(bào),2012,13(4):695-698.
[3]李璞,陳宇瓊,黃火強(qiáng).艾納香化學(xué)成分與藥理活性研究進(jìn)展[J].實(shí)用中醫(yī)內(nèi)科雜志,2012,26(10):3-6.
[4]嚴(yán)啟新,譚道鵬,康暉.艾納香中的黃酮類化學(xué)成分[J] .中國(guó)實(shí)驗(yàn)方劑學(xué)雜志,2012,18 (5):86-89.
[5]鄧芹英,丁叢梅,張維漢.艾納香中黃酮化合物的研究[J].波譜學(xué)雜志,1996,13(5):447-452.
[6]趙金華,康暉,姚光輝,等.艾納香化學(xué)成分研究[J].中草藥,2007,38(3):350-352.
[7]白志文,朱露,孫濟(jì)平,等.苗藥艾納香研究進(jìn)展[J].中國(guó)民族醫(yī)藥雜志,2012,7(7):65.
[8]Gershenzon J,Dudareva N.The function of terpenenatural products in the natural world [J].Nat Chem Biol,2007,3(7):408-414.
[9]海廣范,張慧,郭蘭青.二萜類化合物藥理學(xué)作用研究進(jìn)展[J].新鄉(xiāng)醫(yī)學(xué)院,2015,32(1):77-80.
[10]陳建,趙建剛,植物萜類生物合成相關(guān)酶類及其編碼基因的研究進(jìn)展[A].分子植物育種,2014,2(6):757-764.
[11]馬利剛,趙樂.北葶藶子GGPS基因的克隆、序列分析與原核表達(dá)[J].中醫(yī)藥現(xiàn)代化,2015,3(7):154-156.
[12]化文平,宋雙紅.丹參 SmGGGPS3 基因的克隆及表達(dá)分析[J].植物科學(xué)學(xué)報(bào) 2014,32(1):50-57.
[13]李翠萍,夏蓓蓓.番茄 GGPS2 基因的克隆與表達(dá)載體的構(gòu)建[J].西南大學(xué)學(xué)報(bào).2010,32(10):65-67.
[14]閔丹丹,唐美瓊.三七香葉基香葉基焦磷酸合酶基因的克隆及表達(dá)分析[J].中國(guó)中藥雜志.2015,40(11):54-58.
Analysis of geranylgeranyl pyrophosphate synthase (GGPS) sequence of Blumea balsamifera(L.)·DC based on transcriptome information
XIA Qi-feng1,ZHAO Zhi2*,LIU Hong-chang2,GUAN Ling-liang3*,PANG Yu-xin3
(1.Guizhou University,College of Life Sciences,Guiyang,Guizhou 550025,China;2.Guizhou University College of Agriculture,Guiyang,Guizhou 550025,China;3.Tropical Crops Genetic Resources Institute,Chinese Academy of Tropical Agricultural Sciences,Danzhou,Hainan 571737,China)
A geranylgeranyl diphosphate synthase gene,designatedBbGGPS,has been isolated from Blumea balsamifera(L.) DC using reverse transcription polymerase chain reaction approach (RT-PCR) and rapid amplification of cDNA ends (RACE) methods.The results showed that the BbGGPS cDNA had a full length of 1 475 bp,and contained an open reading frame predicting a polypeptide of 334 amino acids.Hydropathy and subcellular localization prediction showed that the BbGGPS belongs to hydrophilic protein and is located in Chloroplasts.It is neither a membrane protein nor secretory protein.BbGGPS protein showed a high similarity with other plant GGPS genes.Phylogenetic analysis indicated that all the amino acid sequence were divided into five categories and BbGGPS was grouped with that of Cynara cardunculus var.
Blumea balsamifera;geranylgeranyl diphosphate synthase;amino acid sequence;phylogenetic analysis
2016-05-21;
2016-06-14
國(guó)家自然科學(xué)基金(NO.81202910);中央級(jí)公益性科研院所基本科研業(yè)務(wù)費(fèi)專項(xiàng)資金(1630032014015);海南省中藥現(xiàn)代化專項(xiàng)(ZY201410)。
趙致(1957-),男,教授,主要研究方向:作物生理生態(tài)理論及其應(yīng)用;E-mail:zzhao@gzu.edu.cn。
Q756
A
1008-0457(2016)04-0023-07國(guó)際
10.15958/j.cnki.sdnyswxb.2016.04.005
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
當(dāng)代經(jīng)濟(jì)研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國(guó)中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學(xué)學(xué)報(bào)(社會(huì)科學(xué)版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
