煙臺黑豬SLA-DQA基因編碼區(qū)多態(tài)性及生物信息學分析
2016-11-17 08:02:35黃曉宇袁軍虎楊巧麗馬艷萍滾雙寶
華北農學報 2016年5期
關鍵詞:結構
黃曉宇,袁軍虎,楊巧麗,馬艷萍,滾雙寶,3
(1.甘肅農業(yè)大學 動物科學技術學院,甘肅 蘭州 730070;2.河南省漯河市畜牧局,河南 漯河 462000;3.甘肅省現代養(yǎng)豬工程技術研究中心,甘肅 蘭州 730070)
?
煙臺黑豬SLA-DQA基因編碼區(qū)多態(tài)性及生物信息學分析
黃曉宇1,袁軍虎2,楊巧麗1,馬艷萍1,滾雙寶1,3
(1.甘肅農業(yè)大學 動物科學技術學院,甘肅 蘭州 730070;2.河南省漯河市畜牧局,河南 漯河 462000;3.甘肅省現代養(yǎng)豬工程技術研究中心,甘肅 蘭州 730070)
采用 PCR-SSCP、克隆測序和生物信息學等方法對煙臺黑豬SLA-DQA基因多態(tài)性進行研究,預測其蛋白質結構并分析其功能特征。結果顯示:SLA-DQA基因 exon 2、exon 3、exon 4 分別檢測出4 種等位基因,5 種基因型、6 種等位基因,8 種基因型、4 種等位基因,7 種基因型,包括3種新等位基因。SLA-DQA基因共編碼 255 個氨基酸,編碼區(qū)共發(fā)現 32 個核苷酸突變和 13 個氨基酸變異,exon 2 多態(tài)性最為豐富。蛋白質結構預測結果顯示二級結構中無規(guī)則卷曲和延伸鏈比例最高,α 螺旋數量多且集中,β 折疊最少。功能預測結果顯示,SLA-DQA基因蛋白質在能量代謝、結構蛋白和生長因子等方面幾率較高。本研究結果可為SLA-DQA基因在豬抗病分子育種中的應用提供理論依據。
煙臺黑豬;SLA-DQA基因;編碼區(qū);多態(tài)性;蛋白質結構
豬的主要組織相容性復合體即豬白細胞抗原復合體(Swine lymphocyte antigen,SLA),是豬染色體上一組緊密連鎖、與免疫應答和抗病性密切相關的高度多態(tài)的基因群。SLA抗原定位于 7p12-q12,按其抗原結構和功能主要分為三大類,其中 Ⅱ 類抗原位于SLA-D區(qū),是由34 kDa 的 α 鏈和 29 kDa 的 β 鏈以非共價鍵結合的異質二聚體,主要包括DRA、DRB、DQA、DQB、DOB、DPA、TAP、LMP等基因[1-3]。SLAⅡ類DQA和DRA基因位于抗原呈遞細胞(APC)表面,能夠將抗原呈遞給 CD4+T細胞,從而參與并調控機體的免疫應答反應。基因產物間的變異是特異的抗原結合和排斥反應的基礎,抗原結合位點的多態(tài)性可確定免疫識別相關的功能性多態(tài),在研究機體的抗病能力方面具有非常重要的作用。SLA基因是豬染色體中最富有特性的區(qū)域,大量研究表明,具有SLAⅡ 類基因不同基因型的個體對疾病抵抗能力具有明顯差異,不同DQA和DRA基因型顯著影響仔豬腹瀉情況[4-9]。研究顯示不同地域品種豬有各自獨特的生物多樣性和遺傳穩(wěn)定性特征,使得不同地域特征性的豬種SLAⅡ 基因表現出不同程度的多態(tài)性,人們通過克隆測序等技術對廣西巴馬豬、海南五指山豬、八眉豬、榮昌豬及湖南沙子嶺豬等不同地域豬種SLAⅡ類基因DQ和DR基因的多態(tài)性水平、蛋白質結構和功能、同源性進行分析[10-14],發(fā)現豬SLA與人類HLA基因之間具有高度的同源性,這種相對低度的免疫排斥反應,將有助于豬-人異種器官移植免疫排斥反應的研究,但目前尚未見到對煙臺黑豬SLAⅡ類基因多態(tài)性的報道。
煙臺黑豬始產于膠東半島,是以膠東地方灰皮黑豬為基礎,經選育而成的一種優(yōu)良豬種,具有耐粗飼、易飼養(yǎng)、抗逆性強、產仔率高等特點,對當地生豬生產和品種改良起到了重要作用,又因其豬肉品質、色澤、口感及風味較好,近年來更是倍受消費者青睞[15]。因此,探討煙臺黑豬腹瀉的抗病性或易感性對于更好地利用其品種資源優(yōu)勢具有重要意義。
試驗以甘肅紅古區(qū)引入的煙臺黑豬為研究對象,采用 PCR-SSCP、克隆測序和生物信息學軟件等方法,研究煙臺黑豬SLA-DQA基因編碼區(qū)(Coding region,CDS)多態(tài)性,氨基酸組成并對其CDS 蛋白質的結構和功能進行預測分析,以期探究煙臺黑豬SLA-DQA基因的遺傳特性、蛋白質結構和功能的關系以及其作為候選基因在豬抗病育種方面的潛力,為今后在豬遺傳育種候選基因的研究提供一定可靠的理論依據。
1 材料和方法
1.1 試驗材料
試驗對象為甘肅省紅古區(qū)黑豬飼養(yǎng)場飼養(yǎng)條件相同的 290頭煙臺黑豬。采集耳組織樣品約 3 g 置于盛有 75% 乙醇的 5 mL EP 管中,并在 DNA 降解前立即送到實驗室保存于-20 ℃。根據 Sambrook等[16]的常規(guī)酚/氯仿抽提法提取基因組 DNA,TE 溶解,用瓊脂糖凝膠電泳和紫外分光光度法檢測 DNA 的純度和濃度,再稀釋成終濃度為 100 ng/μL 置于-20 ℃ 保存?zhèn)溆谩?/p>
1.2 引物設計和 PCR 擴增
參照 NCBI 數據庫SLA-DQA基因序列 (登錄號 AY 303988),采用 Primer 5.0 引物設計軟件設計SLA-DQA基因 exon 2、exon 3、exon 4 擴增引物,預期擴增片段分別為 378,388,278 bp,引物由大連寶生物公司合成。引物序列參見表 1。
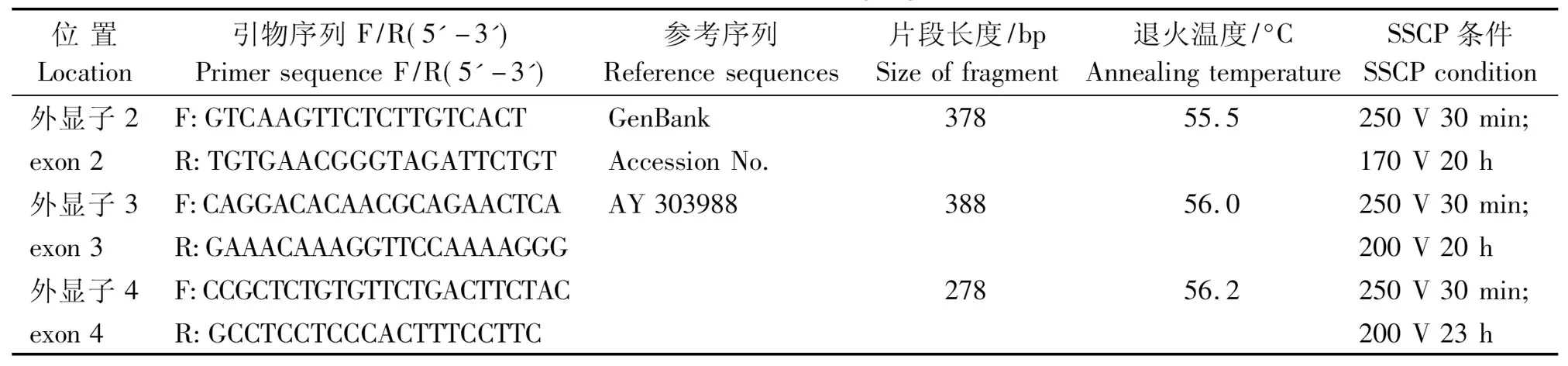
表1 SLA-DQA 基因 exon 2、exon 3、exon 4 的引物序列和 SSCP 反應條件
PCR 擴增采用 25 μL 的反應體系:10×Buffer 緩沖液 2.5 μL,dNTP 1 μL(2.5 mmol/μL),上下游引物各 0.5 μL(10 pmol/μL),TaqDNA聚合酶0.5 μL(5 U/μL),模板 DNA 1 μL(50~100 ng/μL),滅菌 ddH2O 19 μL。PCR 擴增條件:預變性 94 ℃ 3 min;變性 94 ℃ 30 s,退火溫度詳見表 1,延伸 72 ℃ 30 s,35 個循環(huán);最后延伸 72 ℃ 10 min,4 ℃ 保存,PCR 產物用2%的瓊脂糖凝膠電泳檢測。
1.3 PCR 擴增產物的 SSCP 檢測
分別取 3 μLSLA-DQAPCR 產物,加入 7 μL 變性劑(98% 去離子甲酰胺、0.025% 溴酚藍、0.025% 二甲苯青、10 mmol/L EDTA(pH 值0.8)混合),經 98 ℃變性 10 min,迅速冰浴 10 min,上樣于充分預冷的 10% 非變性聚丙烯酰胺凝膠(Acr∶Bis=39∶1),SLA-DQA基因3個外顯子 PCR 擴增產物混合液 4 ℃ 條件下進行電泳,電泳條件見表 1,電泳結束后銀染法顯色。
1.4 基因的克隆測序
根據不同 SSCP 模型條帶,每種基因型隨機挑取 3 個不同個體的 PCR 產物,用瓊脂糖凝膠 DNA 回收試劑盒回收純化,純化后的產物用載體pMD?19-T Vector 連接,構建重組質粒,并轉化大腸桿菌(Escherichiacoli)DH5α 菌株;挑選陽性克隆培養(yǎng),培養(yǎng)后進行菌液 PCR 擴增;菌液 PCR 產物與初始 PCR 產物同時進行 SSCP 檢測,再次通過 10% 的非變性聚丙烯酰胺凝膠電泳(39∶1),顯色后進行模型條帶比較,進一步確定測序菌落的正確性,每一種基因型挑選 2 個克隆的菌液 PCR 產物送上海生物工程有限公司進行測序。
1.5 數據統(tǒng)計分析
PopGene 32 軟件和 PIC軟件(Bostein)[17]計算基因的多態(tài)性信息,MEGA 5.0 軟件進行 DNA 分析和比對;NCBI dbSNP 數據庫(http://www.ncbi.nlm.nih.gov/SNP/index.html)分析單核苷酸多態(tài)性;利用 CBS的 Protfun在線軟件(http://www.cbs.dtu.dk/services/ProtFun)預測蛋白質的功能;ExPASY 服務器 SWISS-MODEL(http://us.expasy.org/)預測蛋白質二級結構,三維結構同源建模和模型分析。
2 結果與分析
2.1SLA-DQA基因核苷酸序列多態(tài)性分析
2.1.1 PCR 擴增及 SSCP 檢測SLA-DQA基因 exon 2、exon 3、exon 4 的 PCR 擴增產物經2%瓊脂糖凝膠電泳檢測,目的條帶清晰且無非特異擴增者,判斷其擴增產物為目的基因,可進行后續(xù)試驗分析。經 SSCP 檢測,exon 2 檢測到 4 種等位基因(A、B、C、D),共形成 5 種基因型(AA、BB、AB、AC、AD,圖 1-A);exon 3 檢測到了 6 種等位基因(A、B、C、D、E、F),共形成 8 種基因型(AA、BB、AB、CC、BC、DD、EF、CF,圖 1-B);exon 4 檢測到了 4 種等位基因(A、B、C、D),共形成 7 種基因型(AA、BB、AB、CC、BC、AD、BD,圖 1-C)。3個外顯子的等位基因頻率和基因型頻率見表 2。
2.1.2SLA-DQA基因 exon 2、exon 3、exon 4 的序列分析 根據參考序列(AY303988),將克隆測序得到的序列去除引物序列及內含子序列,得到有效的外顯子序列,exon 2、exon 3、exon 4 序列長度分別為 249,281,154 bp。將3個外顯子的等位基因序列利用 dbSNP 進行比對,結果發(fā)現 exon 2、exon 3、exon 4 序列分別檢測到 20,9,3 個核苷酸突變位點(表 3),共導致 13 個氨基酸發(fā)生變異。SLA-DQA基因3個外顯子中共包括3種新等位基因:exon 2 中 D等位基因含有7個核苷酸突變位點,exon 3 中 C等位基因包括一個新的突變位點(c.4828T>C),F等位基因包含一個新的突變位點(c.4617T>C)。各等位基因序列提交至GenBank,獲得的登錄號見表2。核苷酸的突變引起氨基酸的變異,exon 2中D等位基因含有7個核苷酸的突變位點,導致3個氨基酸發(fā)生變異,c.3945C>T導致了脯氨酸轉化為絲氨酸,c.4086A>G導致了蘇氨酸轉化為丙氨酸,c.4099A>C的突變導致了天冬酰胺轉化為蘇氨酸;exon 3中C等位基因c.4828T>C的變異導致半胱氨酸轉化為精氨酸,F等位基因c.4617T>C導致絲氨酸轉化為脯氨酸,均屬于錯義突變。
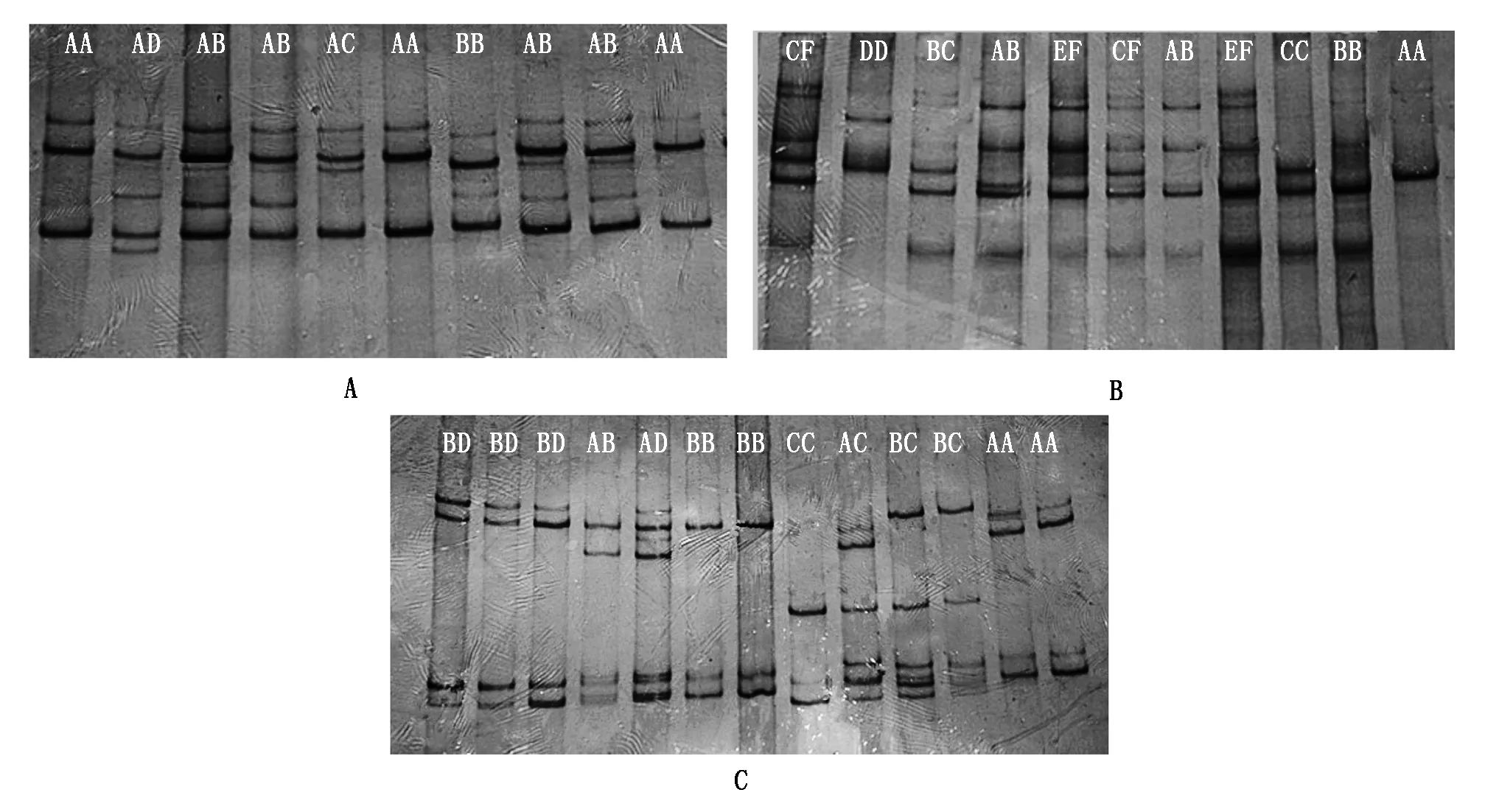
SLA-DQA基因exon 2(A)、exon 3(B)、exon 4(C)分別檢測到 5,8,7 個SSCP不同的帶型。每種帶型代表一種基因型。
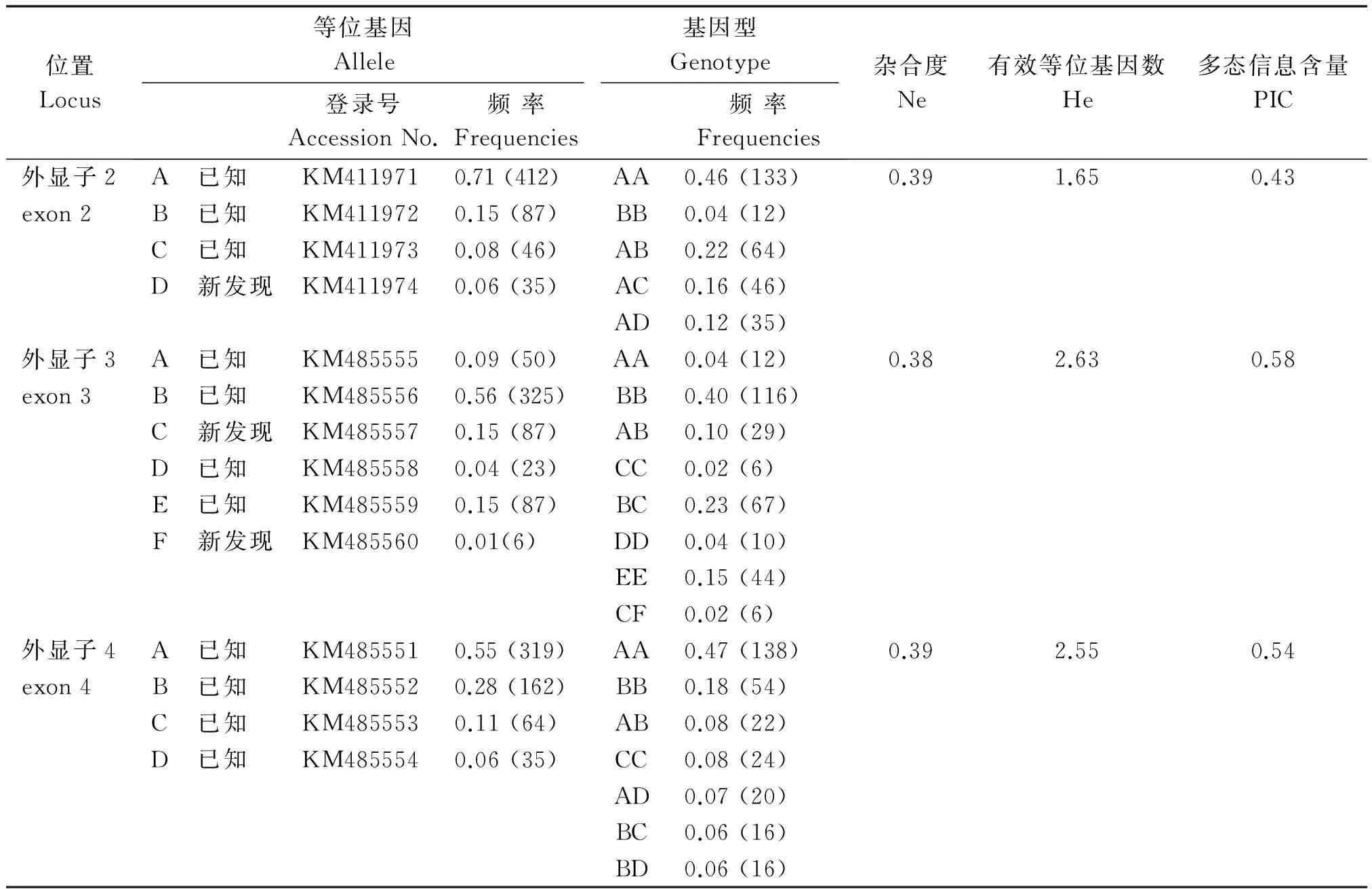
位置Locus等位基因Allele基因型Genotype登錄號AccessionNo.頻率Frequencies頻率Frequencies雜合度Ne有效等位基因數He多態(tài)信息含量PIC外顯子2A已知 KM4119710.71(412)AA0.46(133)0.391.650.43exon2B已知 KM4119720.15(87)BB0.04(12)C已知 KM4119730.08(46)AB0.22(64)D新發(fā)現KM4119740.06(35)AC0.16(46)AD0.12(35)外顯子3A已知 KM4855550.09(50)AA0.04(12)0.382.630.58exon3B已知 KM4855560.56(325)BB0.40(116)C新發(fā)現KM4855570.15(87)AB0.10(29)D已知 KM4855580.04(23)CC0.02(6)E已知 KM4855590.15(87)BC0.23(67)F新發(fā)現KM4855600.01(6)DD0.04(10)EE0.15(44)CF0.02(6)外顯子4A已知 KM4855510.55(319)AA0.47(138)0.392.550.54exon4B已知 KM4855520.28(162)BB0.18(54)C已知 KM4855530.11(64)AB0.08(22)D已知 KM4855540.06(35)CC0.08(24)AD0.07(20)BC0.06(16)BD0.06(16)
2.1.3SLA-DQA基因 exon 2、 exon 3、 exon 4 遺傳多態(tài)性分析 煙臺黑豬SLA-DQA基因 exon 2、exon 3、exon 4 的基因型和等位基因頻率見表 2。遺傳雜合度(He)、有效等位基因數(Ne)和多態(tài)信息含量(PIC)是評價群體遺傳變異的重要指標,不同的遺傳參數體現各群體的遺傳差異性。由表 2可以看出,煙臺黑豬SLA-DQA基因3個外顯子的 He 值為1.65~2.63,雜合度較高;根據 Vaiman 等[18]的研究結論,exon 2的PIC值為 0.43,表現為中度多態(tài)性(PIC<0.50),exon 3、exon 4 的PIC值分別為 0.58,0.54,表現為高度多態(tài)性(PIC>0.50)。

表3 煙臺黑豬 SLA-DQA 基因 exon 2、exon 3、exon 4 突變位點
注:方框標記為新突變位點。
Note:The box marked are the novel mutation sites.
2.2SLA-DQA基因 CDS 區(qū)蛋白質的結構功能預測
2.2.1SLA-DQA基因蛋白質功能特性預測 對比 GenBank 數據庫SLA-DQA基因的參考序列登錄號AY303988,煙臺黑豬SLA-DQA基因CDS核苷酸序列長度為768 bp,共編碼255個氨基酸,SLA-DQA基因與AY303988的CDS氨基酸對比結果詳見圖 2。SLA-DQA基因與 AY303988 的 CDS 蛋白質功能預測詳見表 4,由表 4可以看出,預測的磷酸化位點,轉移數差異較大,能量代謝、結構蛋白、脂肪酸代謝和等電點數值均較高。SLA-DQA基因蛋白質的二級結構預測見圖 3,其中 α 螺旋率 22.35%(57個),β 折疊率6.67%(17個),無規(guī)則卷曲率 40.00%(102個),延伸鏈率為 30.98%(79個)。

箭頭標記的氨基酸為配位體,由氨基酸序列的第144位天冬酰胺(N),192位谷氨酸(E)和193位組氨酸(H)組成。
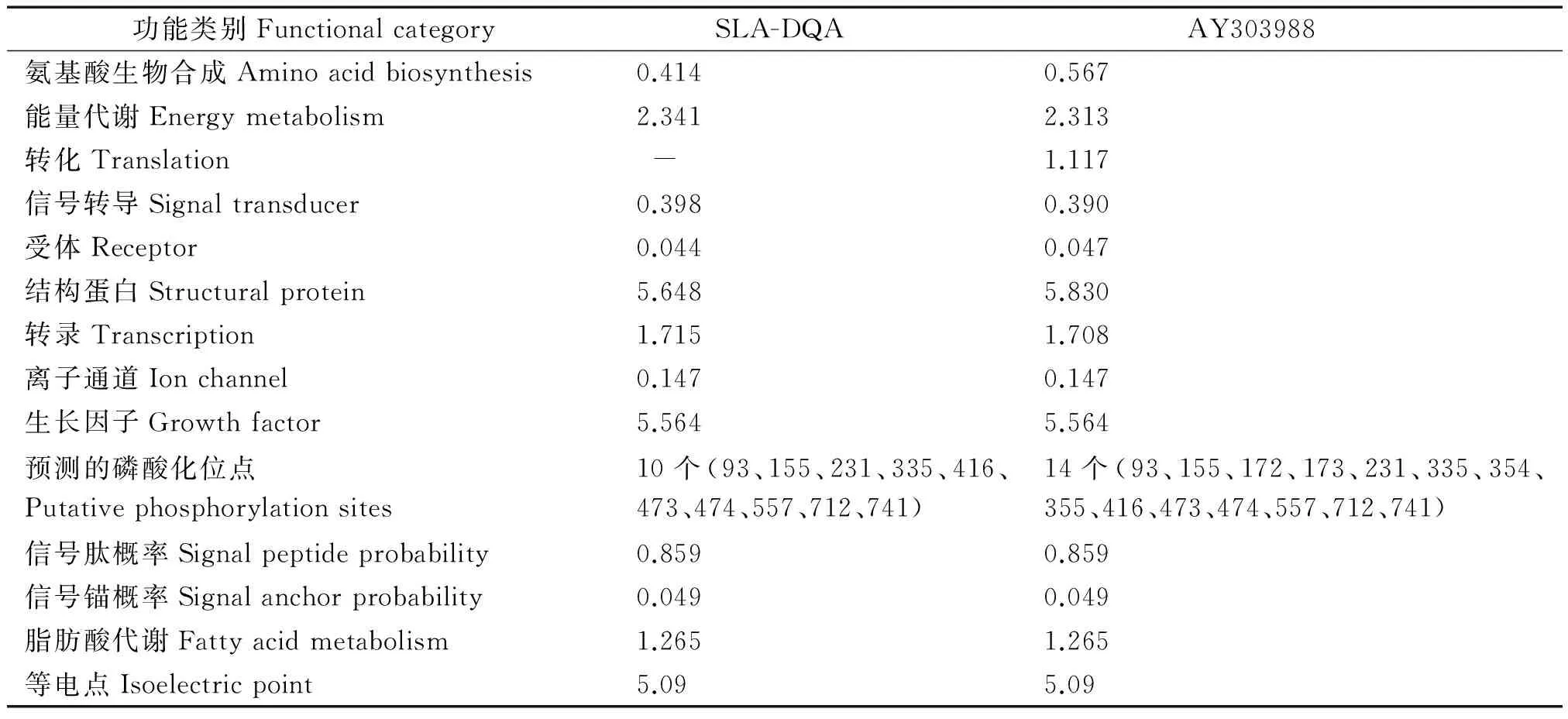
功能類別FunctionalcategorySLA-DQA AY303988 氨基酸生物合成Aminoacidbiosynthesis0.4140.567能量代謝Energymetabolism2.3412.313轉化Translation -1.117信號轉導Signaltransducer0.3980.390受體Receptor0.0440.047結構蛋白Structuralprotein5.6485.830轉錄Transcription1.7151.708離子通道Ionchannel0.1470.147生長因子Growthfactor5.5645.564預測的磷酸化位點Putativephosphorylationsites10個(93、155、231、335、416、473、474、557、712、741)14個(93、155、172、173、231、335、354、355、416、473、474、557、712、741)信號肽概率Signalpeptideprobability0.8590.859信號錨概率Signalanchorprobability0.0490.049脂肪酸代謝Fattyacidmetabolism1.2651.265等電點Isoelectricpoint5.095.09

c.無規(guī)則卷曲區(qū);e.延伸鏈區(qū);h.α-螺旋區(qū);t.β-折疊區(qū)。
2.2.2SLA-DQA基因蛋白質三級結構的同源建模 通過在線軟件 SWISS-MODEL 數據庫 Blast 進行分析,以序列 3pl6.1.A(2.55?)[19]為模板,對提交的SLA-DQA基因氨基酸序列進行蛋白質三維結構同源建模,預測了第 25-206 位氨基酸的三維結構,占氨基酸總長度的70.6%,模型結果見圖 4。SLA-DQA基因與模板序列相似性為 0.55%,序列覆蓋率為 76%,序列一致性達到 80.93%。全球模型質量評估(GMQE)得分為 0.69[20-21]。試驗發(fā)現1個 N-糖蛋白配位體(N-Acetyl-D-Glucosamine,NAG),分子式為 C8H15NO6,分子量為 221.208 g/mol。在SLA-DQA基因蛋白質的三維結構模型中還預測到一個配位體,該配位體是由蛋白質序列上的第144位N(天冬酰胺),第192位E(谷氨酸)和第193位H(組氨酸)3個氨基酸組成(圖2, 4)。
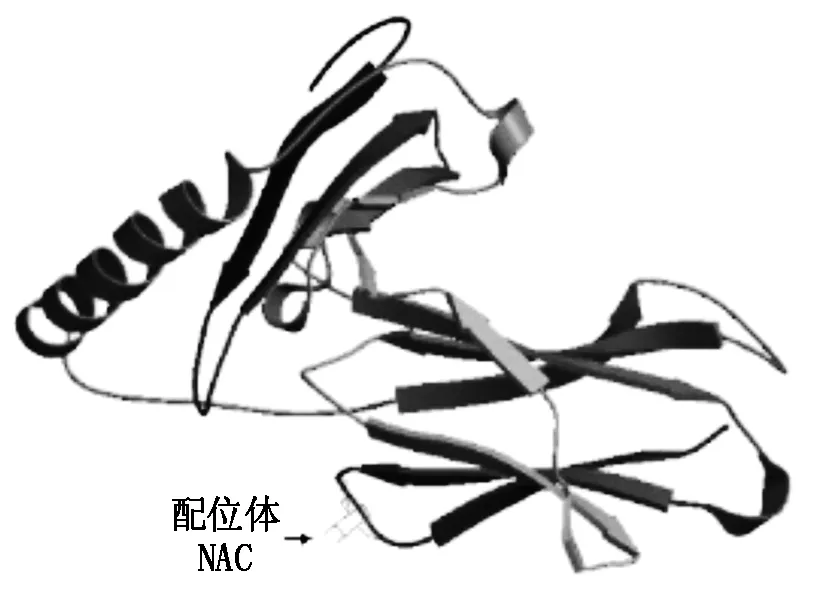
圖4 煙臺黑豬 SLA-DQA基因蛋白三級結構模型預測
2.2.3SLA-DQA基因三維結構合理性分析 運用 DeepView 軟件拉式構圖檢測模擬SLA-DQA基因蛋白質三維模型的合理性。模擬蛋白質三維模型包括 182 個氨基酸,其中 178 個氨基酸殘基(97.8%)的二面角落在允許的范圍內(a區(qū)域),僅有 4 個氨基酸殘基(2.2%)的二面角落在不允許范圍內(a區(qū)域外,b區(qū)域內為不完全允許區(qū),c區(qū)域內為不允許區(qū))(圖 5)。結果表明,SLA-DQA基因的三維模型的二面角分布和立體構象合理,均符合立體化學的二面角 (φ,ψ)分布要求。

a.允許區(qū);b.臨界限制區(qū);c.不允許區(qū)。
3 討論與結論
豬的SLA基因是基因組中最具遺傳多態(tài)性的基因,截至目前,在豬的 MHC數據庫中僅定義了 20DQA、44DQB、13DRA、82DRB1 種等位基因,相較于 β 鏈DQB和DRB基因的高度多態(tài)性,α 鏈DQA基因一般呈現出中等程度多態(tài)性,這種核苷酸的多態(tài)性將會導致相應氨基酸的變化,可能會影響SLAⅡ類 α 鏈基因的功能[22],較低的DQA基因多態(tài)性可能是由降低的積極選擇壓力造成的[2]。自然選擇過程中等位基因的積累和融合、生存環(huán)境、個體對外來抗原的選擇壓力的差異以及病原體驅動下的核苷酸替換都有利于等位基因多態(tài)性的進化[23],通過MHC基因的變異對蛋白質結構和功能進行分析和預測,可有效地評估種群遺傳多樣性水平,為更加深入的研究SLA-DQA基因作為疾病和生產的候選基因提供理論基礎。
本試驗在SLA-DQA基因 CDS 區(qū)共檢測到 32 個SNPs,具有豐富的遺傳多態(tài)性。其中 exon 2 檢測 4 種等位基因和5種基因型(AA、BB、AB、AC和AD),沒有檢測到 BC 型,與孔晶晶[24]的研究結果相比,發(fā)現有2種基因型完全一致,2種等位基因不相同,進一步說明SLA-DQA基因外顯子 2 具有非常豐富的多態(tài)性。SLA-DQA基因的3個外顯子多態(tài)性分布極不平衡,exon 2 包括20 個核苷酸變異位點,A 等位基因頻率為 0.71,屬于優(yōu)勢等位基因,多態(tài)性最豐富,exon 3 和 exon 4 分別包括 9,3 個核苷酸變異位點,exon 3 中 B 等位基因和 exon 4 中 A等位基因的基因頻率分別為0.56和0.55,高于其他等位基因頻率,屬于優(yōu)勢等位基因,exon 3 和 exon 4 基因多態(tài)性較低,這種現象可能與基因的保守性有關。保守性機制是由基因在動物機體擔任重要功能所決定,具有調節(jié)和控制機體的免疫應答、抵御外源環(huán)境壓力、維持正常生理功能和自身進化的作用。煙臺黑豬作為地方性選育豬種,長期受到人工選擇的壓力作用,打破了個體交配的隨機性,造成基因分布的不平衡,使得某種基因型成為特定基因位點的優(yōu)勢基因型。SLA-DQA基因 exon 2 的 PIC值為 0.25~0.50。表現為中度多態(tài),這與Liu 等[5]、Yang等[8]、Ho等[22]和 Lunney等[1]的研究結果一致。MHC 基因組外顯子功能區(qū)堿基的變異將導致功能區(qū)氨基酸的突變,從而影響蛋白質的穩(wěn)定性,最終在抗原的識別與遞呈、免疫應答與調控等方面發(fā)揮作用。
本試驗中SLA-DQA基因氨基酸序列包含α螺旋、β折疊、無規(guī)則卷曲和延伸鏈的氨基酸數分別為 57,17,102,79個。α 螺旋和 β 折疊2種結構均含有氫鍵,鍵能較高,能夠牢固地維持蛋白質的高級結構且常位于蛋白質的內部;延伸鏈和無規(guī)則卷曲區(qū)域所占比例最高,屬柔型結構,易發(fā)生扭曲,常出現于蛋白表面并與表面抗原結合。SLA-DQA基因與 AY303988 編碼蛋白的能量代謝、脂肪酸代謝、信號轉導、生長因子的幾率均呈現相對較高現象(表 4),其中結構蛋白和生長因子的幾率最高,說明該基因可能對豬的抗原結合能力和生產力起到較高的調節(jié)作用[23]。
同源建模是一種根據蛋白結構保守穩(wěn)定性遠大于蛋白氨基酸序列的理論預測蛋白質三維結構的方法,如果氨基酸序列相似性達 30% 以上,則可用已知蛋白結構作為模板[21,25]模擬蛋白質三維結構。本試驗中預測的SLA-DQA基因蛋白質三維結構只含有一條 α 鏈,由第 25-206 位氨基酸組成 α 鏈的編碼區(qū),占編碼氨基酸總長度的 70.6%,是蛋白質行使生物功能的主要區(qū)域。SLA-DQA基因三維結構合理性分析中,α-螺旋主要出現在坐標軸的第三象限,β-折疊主要出現在第二象限,且均在a區(qū)域內,說明所形成的二面角(φ,ψ)構象能量最低且穩(wěn)定,符合立體化學所允許的構象圖。對蛋白質分子高級結構的研究有助于闡明SLA分子與多肽的結合規(guī)律及其免疫應答的特征,使人們對SLA-DQA基因蛋白的生物分子結構更加直觀、形象,便于今后深入探究SLA-DQA的致病機理、免疫機制等分子生物學功能的結構基礎。
配位體能夠提供電子與中心原子反應,保護中心原子的官能團而穩(wěn)定化合物。本試驗中預測到一個配位體,由 α 鏈上第 144 位 N(天冬酰胺),第 192 位 E(谷氨酸),第 193 位 H(組氨酸)形成,可與 NAG 接觸產生反應形成髓鞘堿性蛋白多肽(Myelin basic protein peptide,MBP)結合位點。MBP 是脊椎動物中樞神經系統(tǒng)髓鞘的主要蛋白質,位于髓鞘漿膜面,能夠維持髓鞘結構和功能的穩(wěn)定性,具有神經組織特異性。1962 年,Laatsch 等[26]首先從豚鼠腦中分離出 MBP,血清中 MBP 的含量可作為判斷 CNS 破壞程度的重要指標。信號肽是蛋白多肽鏈中用于指導蛋白質跨膜轉移(定位)的 N-末端的氨基酸序列,一般由 15~30 個氨基酸組成,蛋白成熟后信號肽將被剪切掉。該試驗中沒有預測到信號肽存在,這個結果可避免試驗中所預測到的 N-糖基化位點,磷酸化位點和抗原表位等落在信號肽區(qū)域而被剪切掉。蛋白質的糖基化是最重要的翻譯后修飾之一,與蛋白質結構和功能的關系密切。糖基化調控蛋白質在組織和細胞中的定位、功能和活性[27],糖基化位點發(fā)生變化可能與疾病的發(fā)生有關[28],在DQ基因 α 鏈上,第 82 和 121 位的氨基酸是2個糖基化位點[29],其中第 82 位氨基酸位置對應于 exon 2 區(qū)域 4 134~4 145 bp 的核苷酸,即為糖基化修飾區(qū)域。蛋白質的糖基化修飾能夠影響多肽的構象,使多肽鏈具有一定剛性,達到增強蛋白質穩(wěn)定性的作用。因此,SLA-DQA基因潛在的糖基化位點有可能與某些疾病密切相關。
SLA多態(tài)性的差異與豬對疾病抵抗能力的強弱有關。通過開展煙臺黑豬SLA-DQA 基因多態(tài)性研究,建立和探討了 CDS 區(qū)蛋白質的二級、三級結構模擬圖,并對蛋白質的功能和作用有了更進一步的了解,本研究結果為更好地了解 MHC 復合體及其在豬的抗病育種中的應用提供了重要的理論依據。
[1] Lunney J K,Ho C S,Wysocki M,et al.Molecular genetics of the swine major histocompatibility complex,the SLA complex[J].Developmental and Comparative Immunology,2009,33(3,SI):362-374.
[2] Moutou K A,Koutsogiannouli E A,Stanmatis C,et al.Domestication does not narrow MHC diversity inSusscrofa[J].Immunogenetics,2013,65(3):195-209.
[3] Luetkemeier E S,Malhi R S,Beever J E,et al.Diversification of porcine MHC class Ⅱ genes:evidence for selective advantage[J].Immunogenetics,2009,61(2):119-129.
[4] Bao W B,Ye L,Pan Z Y,et al.Microarray analysis of differential gene expression in sensitive and resistant pig toEscherichiacoliF18[J].Animal Genetics,2012,43(5):525-534.
[5] Liu L X,Zhao S G,Lu H N,et al.Association between polymorphisms of the swineMHC-DQAgene and diarrhoea in three Chinese native piglets[J].International Journal of Immunogenetics,2015,42(3):208-216.
[6] Huang X Y,Yang Q L,Yuan J H,et al.Polymorphism and haplotype analyses of swine leukocyte antigen DQA exons 2,3,4,and their associations with Piglet diarrhea in Chinese native pig[J].Genetics and Molecular Research,2015,14(3):10461-10472.
[7] Yang Q L,Zhao S G,Wang D W,et al.Association between genetic polymorphism in the Swine Leukocyte antigen-DRA gene and piglet diarrhea in three Chinese pig breeds[J].Asian-Australasian Journal of Animal Sciences,2014,27(9):1228-1235.
[8] Yang Q L,Kong J J,Wang D W,et al.Swine leukocyte antigen-DQA gene variation and its association with piglet diarrhea in large white,landrace and duroc[J].Asian-Australasian Journal of Animal Sciences,2013,26(8):1065-1071.
[9] 楊巧麗,孔晶晶,趙生國,等.豬SLA-DRA基因外顯子2多態(tài)性及其與仔豬腹瀉的關聯分析[J].畜牧獸醫(yī)學報,2012,43(7):1020-1027.
[10] 劉子展,王在貴,李 奎,等.廣西巴馬小型豬白細胞抗原經典Ⅱ類基因克隆及生物信息學分析[J].畜牧獸醫(yī)學報,2012,43(9):1353-1359.
[11] Zheng X L,Wang F,Wei L M,et al.Cloning and cDNA sequence analysis ofSLA-DQAgene from Hainan wuzhishan miniature pig[J].Animal Husbandry and Feed Science,2009,1(1):17-21.
[12] 劉麗霞,張 麗,趙生國,等.八眉豬DQA基因編碼區(qū)多態(tài)性及生物信息學分析[J].華北農學報,2015,30(1):103-108.
[13] 白小青,劉 文,黃 微,等.榮昌豬SLA-DQB基因β1結構域突變分析及蛋白質序列模式預測[J].畜牧獸醫(yī)學報,2012,43(8):1306-1309.
[14] 唐醫(yī)亞,邢曉為,薛立群,等.湖南沙子嶺豬SLA-DR基因克隆及生物信息學分析[J].遺傳,2007,29(12):1491-1496.
[15] 許剛璞,許瑜偉.煙臺黑豬的調查報告[J].中國畜禽種業(yè),2011,7(6):61-62.
[16] Sambrook J,Fritsch E F,Maniatis T.Molecular cloning:A laboratory manual[M].Beijing:Science Press,1996:464-467.
[17] Botstein D,White R L,Skolnick M,et al.Construction of a genetic linkage map in man using restriction fragment length polymorphisms[J].American Journal of Human Genetics,1980,32(3):314-331.
[18] Vaiman D,Mecier D,Moazmi-Goudarzi K,et al.A set of 99 cattle mircosatellites:characterization,synteny mapping and polymorphism[J].Mammalian Genome,1994,5(5):288-297.
[19] Sethi D K,Schubert D A,Anders A,et al.A highly tilted binding mode by a self-reactive T cell receptor results in altered engagement of peptide and MHC[J].Journal of Experimental Medicine,2011,208(1):91-102.
[20] Lee K H,Wucherpfennig K W,Wiley D C.Structure of a human insulin peptide HLA-DQ8 complex and susceptibility to type 1 diabetes[J].Nature Immunology,2001,2(6):501-507.
[21] Arnold K,Bordoli L,Kopp J,et al.The SWISS-MODEL workspace:a web-based environment for protein structure homology modelling[J].Bioinformatics,2006,22(2):195-201.
[22] Ho C S,Lunney J,Ando A,et al.Nomenclature for factors of the SLA system,update 2008[J].Tissue Antigens,2009,73(4):307-315.
[23] Garamszegi L Z,De Groot N G,Bontron R E.Correlated evolution of nucleotide substitution rates and allelic variation in MHC-DRB lineages of primates[J].BMC Evolutionary Biology,2009,9:73.
[24] 孔晶晶.三個豬品種SLA-DQA基因多態(tài)性與仔豬腹瀉的相關性分析[D].蘭州:甘肅農業(yè)大學,2012.
[25] Biasini M,Bienert S,Waterhouse A,et al.SWISS-MODEL:modelling protein tertiary and quaternary structure using evolutionary information[J].Nucleic Acids Research,2014,42(W1):W252-W258.
[26] Laatsch R L.Glycerol phosphate dehydrogenase activity of developing rat central nervous system[J].Journal of Neurochemistry,1962(9):487-492.
[27] Arnold J N,Wormald M R,Sim R B,et al.The impact of glycosylation on the biological function and structure of human immunoglobulins[J].Annual Review of Immunology,2007,25:21-50.
[28] Litynska A,Przybyuulo M,Pochec E,et al.Comparison of the lectin-binding pattern in different human melanoma cell lines[J].Melanoma Research,2001,11(3):205-212.
[29] Chardon P,Renard C,Vaiman M.The major histocompatibility complex in swine[J].Immunological Reviews,1999,167(1):179-192.
Polymorphism and Bioinformatics Analysis ofSLA-DQAGene CDs in Yantai Black Pig
HUANG Xiaoyu1,YUAN Junhu2,YANG Qiaoli1,MA Yanping1,GUN Shuangbao1,3
(1.College of Animal Science and Technology,Gansu Agricultural University,Lanzhou 730070,China;2.Henan Luohe Animal Husbandry Bureau,Luohe 462000,China;3.Gansu Research Center for Swine Production Engineering and Technology,Lanzhou 730070,China)
The PCR-SSCP,cloning sequencing and bioinformatics methods were used to explore the Yantai black pigSLA-DQAgene coding polymorphisms,structural prediction and functional characteristics of the protein.The results showed thatSLA-DQAgene exon 2,exon 3 and 4 were detected 4,6,4 alleles and 5,8,7 genotypes,respectively,and three new alleles were found for the first time.TheSLA-DQAcoding region encoded 255 amino acids,32 nucleotide mutations and 13 amino acid mutations were found,and exon 2 showed the most polymorphism.Protein prediction results showed that random coil and extended chain accounted for the highest level,alpha helix distribution concentrated,beta sheet appeared the least.The putative result ofSLA-DQAgene protein function showed that the odds of energy metabolism,structural protein and growth factor were higher.These results could provide the theory foundation for the application ofSLA-DQAgene to pig disease resistance molecular breeding.
Yantai black pig;SLA-DQAgene;Coding region;Polymorphism;Protein structure
2016-06-18
DQA和DRA基因與仔豬腹瀉死亡的分子標記技術項目(GNSW-2008-04);高繁殖力豬新品系選育研究項目(092NKDA036);甘肅省優(yōu)質豬肉生產配套技術研究與示范項目(0804NKCA065)
黃曉宇(1989-),女,河南蘭考人,在讀博士,主要從事動物遺傳育種與繁殖研究。
滾雙寶(1967-),男,甘肅張掖人,教授,博士,主要從事動物遺傳育種與繁殖研究。
S828;Q78
A
1000-7091(2016)05-0086-08
10.7668/hbnxb.2016.05.013
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現代企業(yè)(2015年9期)2015-02-28 18:56:50
