
畢赤酵母異源表達幾丁質酶對大豆核盤菌的生防潛能分析
2016-11-30 11:15:08張福麗和玉婷張可陳晨劉暢樊小寧
天津農業科學 2016年11期
張福麗+和玉婷+張可+陳晨+劉暢+樊小寧
摘 要:將來自于棘孢木霉的幾丁質酶基因整合到了畢赤酵母基因組中,并對表達產物進行了生防潛能分析。結果表明,pH值為6和溫度為40 ℃時幾丁質酶活性最高,表達產物能夠顯著抑制大豆核盤菌的生長,并通過誘導ROS積累和抗性酶活性提高大豆對核盤菌的抗性。
關鍵詞:幾丁質酶;異源表達;酶學特性;生防潛能
中圖分類號:S476 文獻標識碼:A DOI 編碼:10.3969/j.issn.1006-6500.2016.11.002
Abstract: Chitinase from Trichoderma viride was expressed in Pichia pastoris, and the biocontrol potential was analyzed. The results showed that Chitinase had the highest activity when pH was 6 and temperature was 40 ℃. Furthermore, Chitinase significantly inhibited the growth of Sclerotinia sclerotiorum and improved the resistance to sclerotiorum by inducing ROS accumulation and the activity of resistant enzymes.
Key words: Chitinase; heterologous expression; biocontrol potential
大豆(Glycine max (L.) Merrill)是世界上重要的植物油和蛋白來源。核盤菌(Sclerotinia sclerotiorum (Lib.) de Bary)引起的大豆菌核病是一種世界性的大豆病害,是影響大豆產量的第二大病害因素[1]。與化學防治相比,生物防治可以有效克服污染環境、危害人畜健康等缺點,并因此成為當前植物病害防治研究的熱點之一。
木霉(Trichoderma spp.)是世界上公認的生防菌,抗真菌代謝物在木霉的生防功能中起著重要作用。幾丁質酶就是一類重要的抗真菌代謝物,能夠水解真菌細胞壁的主要成分幾丁質[2-3],從而抑制真菌的生長。已有研究表明,該基因的表達能夠增強植物對多種病原菌的抗性。與非轉基因植株相比,表達木霉幾丁質酶的轉基因煙草(Nicotiana tabacum)和土豆(Solanum tuberosum)對交鏈孢霉(Alternaria alternata)、茄鏈格孢(Alternaria solani)和灰葡萄孢菌(Btrytis cinerea)、立枯絲核菌(Rhizoctonia solani)[4],蘋果(Malus pumila)對黑星病菌(Venturia inaequalis)[5],檸檬(Citrus limon)對莖點霉(Phoma tracheiphila)和灰霉病菌(Botrytis cinerea)[6]的抗性都顯著增強,表明幾丁質酶對于增強植物的抗病性具有重要作用。筆者研究了畢赤酵母異源表達的棘孢木霉幾丁質酶對大豆核盤菌的生防機制,為幾丁質酶應用于植物病害防治提供理論依據。
1 材料和方法
1.1 材 料
PDA培養基:土豆200 g,葡萄糖20 g,瓊脂粉15 g,pH 值6.0定容至1 L;幾丁質酶基因(Tachi)來自于棘孢木霉(T. asperellum),酵母表達載體pPIC9K本實驗室保存。
1.2 方 法
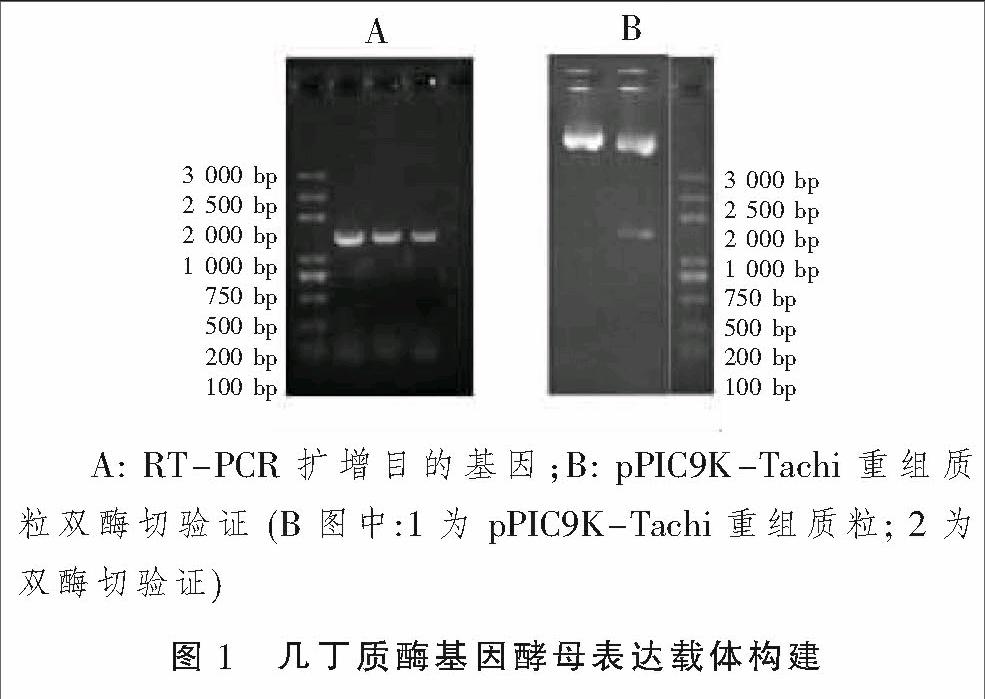
1.2.1 畢赤酵母表達載體構建 在棘孢木霉幾丁質酶基因(Tachi)的CDS序列兩端引入BamHI和KpnI酶切位點,進行PCR擴增并回收目的片段。采用BamHI和KpnI雙酶切,將目的片段連入pPIC9K載體,獲得pPIC9K-Tachi重組質粒,并對重組質粒進行PCR和雙酶切檢測。根據pPIC9K載體的多克隆位點和Tachi基因序列設計并合成以下引物序列:Chi1:5—ATCGGAATTCATGGTCCCTCAGTCTCGAGCC—3 EcoRI (劃線部分為BamHI酶切位點);Chi25—CGATGGTACCTTATTCGTCAAGCCCTCTCTT—3(劃線部分為KpnI酶切位點)。
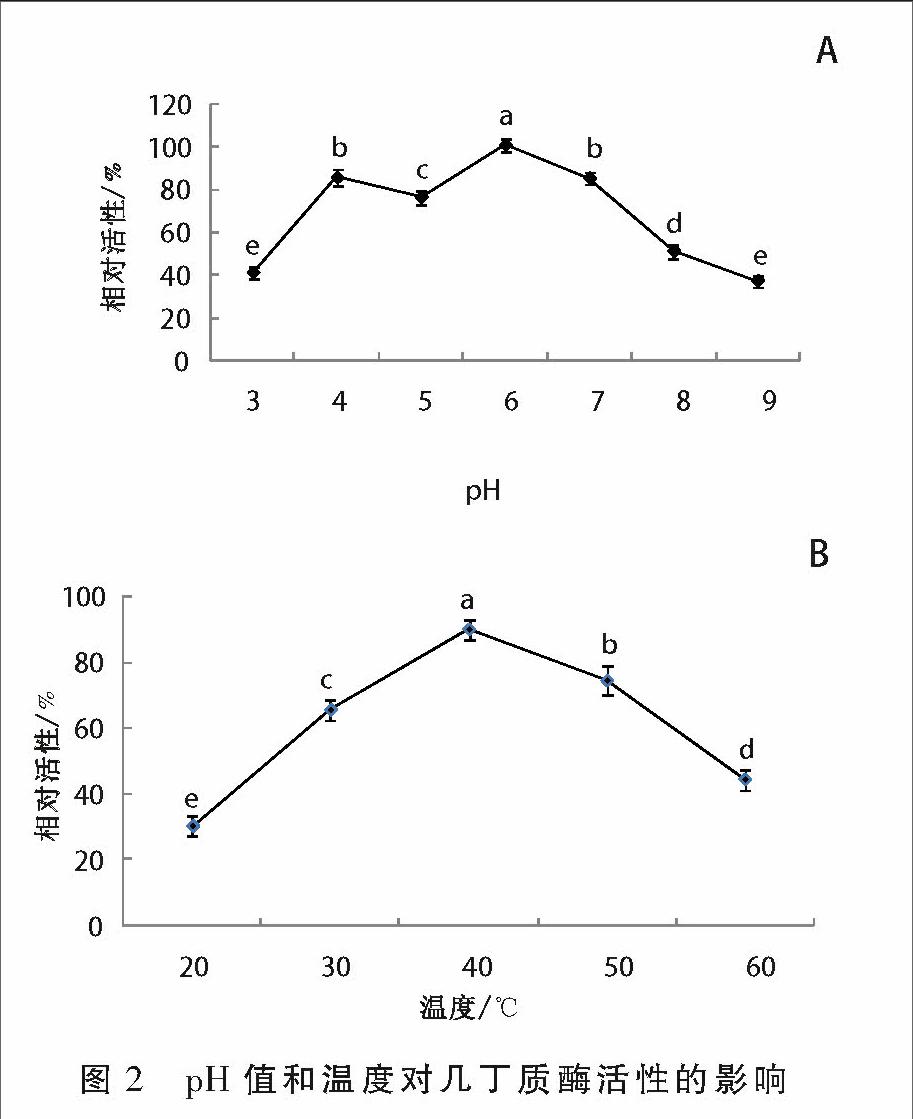
1.2.2 pH值和溫度對幾丁質酶活性的影響 在分析pH值和溫度對幾丁質酶活性的影響時,將酶液分別加入不同pH值(3,4,5,6,7,8,9)的磷酸緩沖液中,首先找出最適pH值,然后在最適pH值條件下,反應溫度分別設定為20,30,40, 50,60 ℃,分別測定溫度對酶活性的影響。酶活性測定參照文獻[7]的方法。
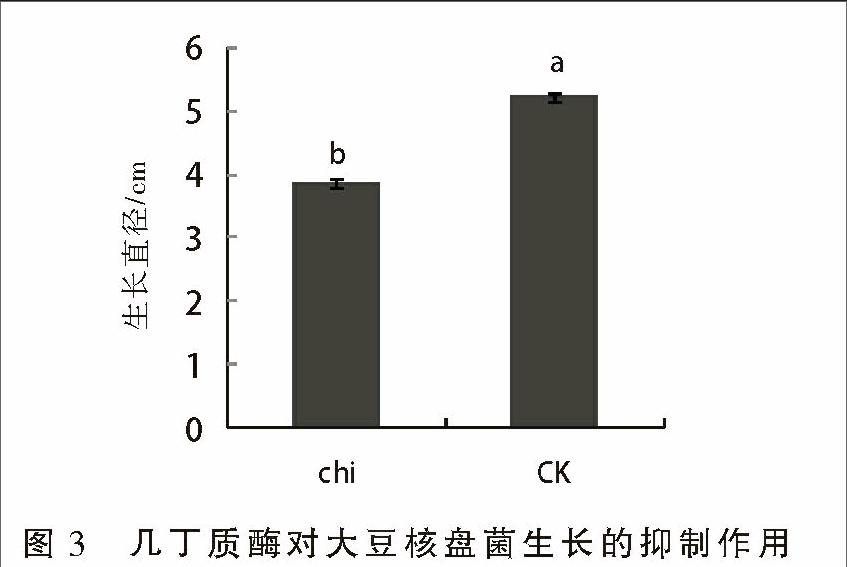
1.2.3 幾丁質酶抑菌活性測定 將培養在PDA培養基上的大豆核盤菌菌塊用打孔器切成直徑為5 mm的菌盤,接種到含有幾丁質酶液的PDA培養基平板的中央,28 ℃培養5 d后,測量菌盤生長直徑,同時以菌盤接種到不含粗酶液的為對照。抑菌率=(對照菌塊直徑-處理菌塊直徑)/對照菌塊直徑×100%。
1.2.4 抗性酶活性及H2O2和O2-含量測定 12 d的大豆幼苗用幾丁質酶液誘導1 d后接種病原菌S.sclerotiorum孢子懸液(1×106孢子·mL-1),26 ℃培養3 d后進行H2O2和O2-含量測定,4 d后測定抗性酶活性。POD和SOD活性測定采用文獻[8-9]的方法。H2O2和O2-含量測定分別采用Mukherjee和Choudhuri的方法[10]。
1.2.5 數據統計分析 采用SPSS 17.0進行數據分析,P<0.05表示差異顯著。
2 結果與分析
2.1 幾丁質酶酵母表達載體的構建及酵母轉化子鑒定
RT-PCR擴增得到全長1 194 bp TachiCDS序列,PCR擴增產物和酵母表達載體pPIC9K分別用BamHI和KpnI雙酶切并進行連接后,得到pPIC9K-Tachi重組質粒。采用PCR的方法用載體構建引物從轉化子中擴增出了大小為1 000~2 000 bp的片斷(圖1A),初步驗證了目的基因連接到了表達載體。提取質粒,再用BamHI和KpnI雙酶切后電泳檢測目的基因片斷,進一步驗證了幾丁質酶基因構建到pPIC9K表達載體中,后經過對所構建載體的序列測定,證實了幾丁質酶Tachi基因已成功構建到酶母表達載體pPIC9K。
2.2 pH值和溫度對幾丁質酶活性的影響
pH值對酶活的影響如圖2A所示,在pH值3~9的范圍內,幾丁質酶均具有一定活性,但pH值為6時酶活性最高。溫度對幾丁質酶活性的影響如圖2B如示,在溫度20~60 ℃范圍內幾丁質酶均有活性,在40 ℃時幾丁質酶酶活性最高。
2.3 幾丁質酶對大豆核盤菌的生長抑制作用
如圖3所示,畢赤酵母表達的幾丁質酶對大豆病原核盤菌具有明顯的抑制作用,培養5 d后,添加幾丁質酶的培養基上核盤菌的生長直徑僅為3.86 cm,而對照培養基上核盤菌的直徑達到5.23 cm,差異顯著。
2.4 幾丁質酶處理誘導了病原脅迫下ROS在大豆中的積累和抗性酶活性
幾丁質酶處理增加了病原脅迫下ROS在大豆幼苗中的積累(圖4)。病原脅迫前,水處理和幾丁質酶分別處理的大豆ROS含量并沒有產生顯著區別(P<0.05)。病原脅迫條件下,水和幾丁質酶處理大豆的ROS含量均有明顯增加,但是水處理的大豆中ROS含量增加較少,與幾丁質酶處理植株之間有顯著區別(P<0.05)。
為了研究抗性酶在清除過多ROS方面的作用,對大豆葉片的抗性酶(POD和SOD)活性進行了測定。結果表明,幾丁質酶處理誘導了大豆葉片抗性酶活性的提高,其中POD活性在幾丁質酶處理和非處理大豆植株之間具有顯著區別(P<0.05)。病原S. sclerotiorum脅迫后大豆葉片的抗性酶活性進一步提高,但幾丁質酶預處理的植株中抗性酶的活性提高較快,與無菌水預處理植株間形成顯著區別(P<0.05)。以上結果表明,幾丁質酶處理誘導了大豆植株抗性酶活性的提高,進而有效清除了病原S. sclerotiorum脅迫下ROS在大豆植株內的過量積累。
2.5 幾丁質酶處理減輕了病原脅迫對大豆的傷害
從圖5可以看出,幾丁質酶處理減少了大豆植株MDA含量和降低了電導率。病原脅迫前,幾丁質酶處理和非處理植株間的MDA和電導率并沒有顯著區別,而在病原脅迫情況下,幾丁質酶預處理大豆植株的電導率和MDA含量顯著低于對照植株(P<0.05),表明幾丁質酶預處理誘導了大豆對病原S. sclerotiorum的抗性,進而減輕了病原脅迫對大豆細胞的傷害。
3 結論與討論
電導率是反映植物抗逆性強弱的指標之一,植物組織外滲液電導率的大小反映了脅迫條件下植物質膜受損傷的程度[11-12]。本研究表明,與對照相比,幾丁質酶預處理后再進行病原脅迫,大豆幼苗的電導率顯著較低,表明幾丁質酶誘導了大豆對核盤菌的抗性,減弱了病原脅迫對大豆質膜的傷害。病原脅迫下,植物體內ROS代謝加強,產生的過氧化物自由基引起膜脂過氧化。丙二醛是植物細胞膜脂過氧化最重要的產物之一,它的產生能加劇植物細胞膜的損傷,是植物抗性生理研究中的一個常用指標,其含量的高低反映了膜脂質過氧化程度、植物細胞膜系統受損程度以及植物的抗逆性。MDA通過與酶蛋白發生鏈式反應聚合,使膜系統變性,造成膜脂流動性降低,影響膜結合酶的功能,引起膜傷害[13-14]。本研究中,與對照相比,幾丁質酶預處理顯著降低了大豆植株中丙二醛含量,表明幾丁質酶預處理提高了大豆植物對核盤菌的抗性,減弱了病原脅迫對大豆細胞膜的傷害。
生物脅迫造成植物體內ROS的快速大量積累,而過多的ROS的存在會對植物細胞造成傷害,為了有效對抗這種氧化脅迫,植物形成了一套有效的ROS清除系統,其中抗性酶在這一系統中起著重要作用。前人的研究表明,來自于綠木霉(T.virens)的幾丁質酶基因通過增加ROS積累、POD活性和萜類化合物含量,以及增加防御相關基因的表達,提高了對病原菌立枯絲核菌Rhizoctonia solani的抗性[15],這與筆者的研究結果一致。
總之,幾丁質酶預處理提高了ROS的積累,進而誘導了抗性酶活性,最終減輕了病原脅迫對大豆植株造成的傷害,表明來自于棘孢木霉的幾丁質酶對大豆核盤菌具有一定的生防潛能。
參考文獻:
[1]ZHAO X,HAN Y,LI Y,et al.Loci and candidate gene identification for resistance to Sclerotinia sclerotiorum in soybean (Glycine max L. Merr.) via association and linkage maps[J].The plant journal : for cell and molecular biology,2015,82(2):245-255.
[2]LINTHORST H. Pathogenesis-related proteins of plants[J]. Critical reviews in plant sciences, 1991,10(2): 123-150.
[3]LORITO M, SCALA F. Microbial genes expressed in transgenic plants to improve disease resistance[J]. Journal of plant pathology, 1999, 81(2): 73-88.
[4]LORITO M,WOO S L,GARCIA I,et al.Genes from mycoparasitic fungi as a source for improving plant resistance to fungal pathogens[J].Proceedings of the national academy of sciences of the United States of America,1998,95(14):7860-7865.
[5]BOLAR J P,NORELLI J L,WONG K W,et al.Expression of endochitinase from Trichoderma harzianum in transgenic apple increases resistance to apple scab and reduces vigor[J].Phytopathology,2000,90(1):72-77.
[6]GENTILE A,DENG Z, MALFA S L,et al.Enhanced resistance to Phoma tracheiphila and Botrytis cinerea in transgenic lemon plants expressing a Trichoderma harzianum chitinase gene[J].Plant breeding,2007,126(2):146-151.
[7]MCCREATH K J,GOODAY G W.A rapid and sensitive microassay for determination of chitinolytic activity[J].Journal of microbiology methods,1992,14(4):229-237.
[8]劉萍,李明軍,李成偉.植物生理學實驗技術[M].北京:科學出版社,2007.
[9]蔡慶生.植物生理學實驗[M].北京:中國農業大學出版社,2013.
[10]MUKHERJEE S P, CHOUDHURI M A. Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in vigna seedlings[J]. Physiologia plantarum,1983, 58(2): 166-170.
[11]邱全勝, 李林, 梁厚果,等. 水分脅迫對小麥根細胞質膜氧化還原系統的影響[J].植物生理學報, 1994(2): 145-151.
[12]張林剛,鄧西平.小麥抗旱性生理生化研究進展[J].干旱地區農業研究,2000,18(3):87-92.
[13]陳少裕.膜脂過氧化與植物逆境脅迫[J].植物通報,1989,6(4):211-217.
[14]王寶山. 生物自由基與植物膜傷害[J]. 植物生理學通訊, 1988 (2): 12-16.
[15]KUMAR V,PARKHI V,KENERLEY C M,et al.Defense-related gene expression and enzyme activities in transgenic cotton plants expressing an endochitinase gene from Trichoderma virens in response to interaction with Rhizoctonia solani[J].Planta,2009,230(2):277-291.
