不同嫩度羊肉中鈣蛋白酶的差異
2016-12-01 08:10:24杜曼婷張彩霞張德權(quán)
中國(guó)農(nóng)業(yè)科學(xué) 2016年17期
杜曼婷,李 欣,李 錚,高 星,張彩霞,張德權(quán)
(中國(guó)農(nóng)業(yè)科學(xué)院農(nóng)產(chǎn)品加工研究所/農(nóng)業(yè)部農(nóng)產(chǎn)品加工重點(diǎn)實(shí)驗(yàn)室,北京 100193)
不同嫩度羊肉中鈣蛋白酶的差異
杜曼婷,李 欣,李 錚,高 星,張彩霞,張德權(quán)
(中國(guó)農(nóng)業(yè)科學(xué)院農(nóng)產(chǎn)品加工研究所/農(nóng)業(yè)部農(nóng)產(chǎn)品加工重點(diǎn)實(shí)驗(yàn)室,北京 100193)
【目的】鈣蛋白酶是宰后肌肉嫩化過(guò)程的主要貢獻(xiàn)者,通過(guò)蛋白水解能力發(fā)揮其功能作用。鈣蛋白酶,尤其是μ-鈣蛋白酶(μ-calpain)的活性與宰后肌肉的嫩度密切相關(guān)。探究不同嫩度的羊肉中鈣蛋白酶的差異,確定羊宰后肌肉中μ-鈣蛋白酶的主要作用時(shí)間及其與嫩度的關(guān)系,為調(diào)控鈣蛋白酶酶活進(jìn)而調(diào)控嫩度提供理論基礎(chǔ)。【方法】選取烏珠穆沁大尾羊背最長(zhǎng)肌為樣品,通過(guò)測(cè)定肌原纖維小片化指數(shù)將樣品進(jìn)行高、低嫩度分組,分別測(cè)定嫩度差異樣品在宰后成熟過(guò)程中pH、μ-鈣蛋白酶大亞基存在狀態(tài),以及未自溶μ-/m-鈣蛋白酶的含量隨宰后時(shí)間的變化情況。【結(jié)果】高、低嫩度組宰后pH的變化規(guī)律一致,宰后各時(shí)間點(diǎn)兩組樣品pH均差異不顯著。μ-鈣蛋白酶80 kDa大亞基在宰后逐漸降解,宰后30 min,高嫩度組μ-鈣蛋白酶80 kDa大亞基含量顯著高于低嫩度組;但高嫩度組μ-鈣蛋白酶80 kDa大亞基在宰后48 h已基本完全降解,低嫩度組在宰后5 d才基本完全降解。未自溶μ-鈣蛋白酶的含量逐漸下降,宰后30 min和2 h,高嫩度組未自溶μ-鈣蛋白酶的含量顯著高于低嫩度組,宰后12 h、5 d和7 d,低嫩度組未自溶μ-鈣蛋白酶的含量顯著高于高嫩度組。宰后6 h,高嫩度組μ-鈣蛋白酶出現(xiàn)明顯的自溶和降解現(xiàn)象。【結(jié)論】與低嫩度組相比,高嫩度組宰后μ-鈣蛋白酶的初始含量較高,但高嫩度組μ-鈣蛋白酶自溶、失活的速率快,自溶及激活程度顯著大于低嫩度組。表明宰后μ-鈣蛋白酶的自溶及激活狀態(tài)直接影響肌肉的嫩度。宰后24 h,尤其是12 h內(nèi),μ-鈣蛋白酶的變化狀態(tài)對(duì)羊肉嫩度的影響最大。
羊肉;肌肉嫩度;μ-鈣蛋白酶;成熟
0 引言
【研究意義】嫩度是肉品的重要品質(zhì)特性之一,直接影響消費(fèi)者的購(gòu)買(mǎi)意向。宰后肌肉的嫩化主要發(fā)生在僵直后的解僵成熟階段,大量研究表明,這一過(guò)程主要受鈣蛋白酶系統(tǒng)的調(diào)控,尤其是μ-鈣蛋白酶(μ-calpain),對(duì)宰后肌原纖維蛋白及細(xì)胞骨架蛋白降解引起的肌肉嫩化具有重要作用。因此,研究不同嫩度羊肉在宰后成熟過(guò)程中鈣蛋白酶的差異,能夠?yàn)橐哉{(diào)控鈣蛋白酶活性為途徑的嫩化方法提供理論基礎(chǔ)。【前人研究進(jìn)展】TAYLOR等[1]在研究Z-線(xiàn)蛋白降解與嫩度的關(guān)系時(shí)發(fā)現(xiàn),μ-鈣蛋白酶在宰后早期被激活,而此期間內(nèi)肌肉中關(guān)鍵的肌原纖維蛋白也在大量降解。LIU等[2]通過(guò)研究僵直前期不同溫度對(duì)鈣蛋白酶活性的影響,證明鈣蛋白酶的激活程度與肌肉嫩度正相關(guān)。GOLL[3]、ZIMMERMAN[4]等研究發(fā)現(xiàn),μ-鈣蛋白酶發(fā)生自溶后會(huì)降低激活其活性所需的Ca2+濃度,使μ-鈣蛋白酶的Ca2+需要量接近生理范圍。ZHANG等[5]在對(duì)牛肉成熟過(guò)程品質(zhì)特性的研究中發(fā)現(xiàn),μ-鈣蛋白酶自溶狀態(tài)出現(xiàn)的越早,可檢測(cè)到的肌間線(xiàn)蛋白、肌聯(lián)蛋白等肌原纖維蛋白的降解情況會(huì)越早。MELODY等[6]在研究宰后早期影響豬肉嫩度和持水力的生化因素時(shí)發(fā)現(xiàn),早期μ-鈣蛋白酶的自溶與μ-鈣蛋白酶的激活及關(guān)鍵蛋白的降解有關(guān)。μ-鈣蛋白酶由80 kDa的大催化亞基和28 kDa的調(diào)節(jié)亞基組成,80 kDa的大亞基包括自溶域、催化域、調(diào)節(jié)域和Ca2+結(jié)合域,其發(fā)揮蛋白水解活性而自溶主要體現(xiàn)為80 kDa大亞基的逐漸降解。GEESINK等[7]以檢測(cè)自溶狀態(tài)的μ-鈣蛋白酶來(lái)反映蛋白的水解情況,將μ-鈣蛋白酶的自溶速率作為反映鈣蛋白酶活性的理想指標(biāo)。【本研究切入點(diǎn)】目前關(guān)于μ-鈣蛋白酶與嫩度關(guān)系的研究主要集中在體外模型試驗(yàn)或間接的μ-鈣蛋白酶活性與嫩度相關(guān)性的研究等方面,對(duì)嫩度差異樣品中μ-鈣蛋白酶的研究也大多僅以μ-鈣蛋白酶活性或自溶狀態(tài)中的一個(gè)指標(biāo)來(lái)反映。而在宰后成熟過(guò)程中,嫩度差異樣品的μ-鈣蛋白酶活性及自溶狀態(tài)的變化情況是否存在直觀(guān)差異以及μ-鈣蛋白酶活性與自溶狀態(tài)的變化是否吻合目前并不清楚。【擬解決的關(guān)鍵問(wèn)題】本研究以品種、年齡、性別、生長(zhǎng)環(huán)境相同的烏珠穆沁大尾羊?yàn)樵囼?yàn)材料,在將樣品分為高、低嫩度組的基礎(chǔ)上,研究不同嫩度羊肉中鈣蛋白酶的差異,明確羊肉宰后成熟過(guò)程中μ-鈣蛋白酶的變化特點(diǎn)及發(fā)揮水解作用的時(shí)間,為通過(guò)調(diào)控μ-鈣蛋白酶活性改善宰后肌肉嫩度的方法途徑提供理論依據(jù)。
1 材料與方法
試驗(yàn)于2015年在中國(guó)農(nóng)業(yè)科學(xué)院農(nóng)產(chǎn)品加工研究所進(jìn)行。
1.1 試驗(yàn)材料
1.1.1 試驗(yàn)樣品 選取50只月齡為7—8個(gè)月的烏珠穆沁大尾羊,按伊斯蘭屠宰方式進(jìn)行屠宰,宰后羊胴體重量約為17 kg。在宰后迅速取下雙側(cè)背最長(zhǎng)肌,冷庫(kù)中快速剔除表面筋膜及脂肪,分別于宰后30 min、1 h、2 h、6 h、12 h、24 h、48 h、72 h、5 d及7 d取等量樣品分裝到速凍管并放入液氮中速凍,保存于-80℃?zhèn)溆茫徊⑼瑫r(shí)測(cè)定各時(shí)間點(diǎn)肉樣的pH。
1.1.2 試劑材料 蛋白濃度測(cè)定試劑盒、PVDF膜購(gòu)自美國(guó)Thermo公司;μ-鈣蛋白酶單克隆抗體、辣根過(guò)氧化酶標(biāo)記的羊抗鼠IgG、亮抑酶肽、三羥甲基氨基甲烷(Tris Base)、十二烷基硫酸鈉、丙烯酰胺、甲叉雙丙烯酰胺、過(guò)硫酸銨、四甲基乙二胺購(gòu)自美國(guó)Sigma公司;ECL顯色試劑盒購(gòu)自美國(guó)Bio-Rad公司;甲醇、乙醇、乙酸等為國(guó)產(chǎn)分析純?cè)噭?gòu)自國(guó)藥集團(tuán)化學(xué)試劑有限公司。
Testo 205便攜式pH計(jì),德國(guó)德圖公司;Ultra TurraxDisperser S25分散器,德國(guó)IKA公司;Neofuge高效冷凍離心機(jī),上海力申科學(xué)儀器有限公司;T6紫外可見(jiàn)分光光度計(jì),北京普析通用儀器有限責(zé)任公司;ML204/02 電子天平,上海梅特勒-托利多有限公司;SpectraMax 190全波長(zhǎng)酶標(biāo)儀,美國(guó)Molecular Devices公司;電泳設(shè)備(Mini-PROTEAN Tetra System)、轉(zhuǎn)膜設(shè)備(Mini Trans-Blot)及ChemiDocTMMP凝膠成像系統(tǒng),美國(guó)Bio-Rad公司。
1.2 試驗(yàn)方法
1.2.1 蛋白提取 參考VEISETH等[8]的方法,取1 g冷凍的羊背最長(zhǎng)肌樣品,加入三倍體積的抽提液(100 mmol·L-1Tris base,10 mmol·L-1EDTA,0.05%巰基乙醇,6 mg·L-1亮抑酶肽,pH 8.3)后冰上勻漿30 s(每勻漿10 s,間隔30 s),所得的勻漿液在10 000×g、4℃條件下離心35 min,上清液即為所需的蛋白樣品。BCA法(Pierce BCA Protein Assay Kit,Thermo)測(cè)定蛋白濃度,保存于-80℃?zhèn)溆谩?/p>
1.2.2 pH 取樣過(guò)程中,使用便攜式pH計(jì)(Testo 205,德圖,德國(guó))分別于宰后30 min、1 h、2 h、6 h、12 h、24 h、48 h、72 h、5 d及7 d測(cè)定各時(shí)間點(diǎn)肉樣的pH。探頭插入深度為2 cm,連續(xù)測(cè)定3次,結(jié)果取平均值。
1.2.3 肌原纖維小片化指數(shù)(MFI) 參考CULLER等[9]的方法,并稍作修改。稱(chēng)取0.5 g肉樣,切碎后加入5 mL預(yù)冷的MFI緩沖液(100 mmol·L-1KCl、20 mmol·L-1K2HPO4、1 mmol·L-1EDTA、1 mmol·L-1MgCl2、1 mmol·L-1NaN3),高速勻漿3次,每次30 s(中間間隔1 min),勻漿后3 000×g、4℃下離心15 min;將沉淀用5 mL預(yù)冷的MFI緩沖液懸浮,再在3 000×g、4℃下離心15 min,棄去上清;用1.25 mL預(yù)冷的MFI緩沖液將沉淀充分懸浮,將懸浮液用20目篩網(wǎng)過(guò)濾除去結(jié)締組織,再加1.25 mL MFI緩沖液清洗離心管并幫助肌原纖維通過(guò)篩孔。過(guò)濾后的懸浮液用雙縮脲法測(cè)定蛋白濃度,然后用MFI緩沖液調(diào)整懸浮液蛋白濃度為(0.5±0.05)mg·mL-1,在540 nm測(cè)定吸光度,將所得結(jié)果乘200后即可得到MFI值。
1.2.4 蛋白質(zhì)免疫印跡 參考LOMIWES[10]等的方法并稍作修改。提取的肌漿蛋白樣品調(diào)整至統(tǒng)一濃度后,按1∶1(v/v)加入SDS-PAGE上樣緩沖液,沸水浴中加熱5 min,冷卻后12 000×g離心1 min,取上清液上樣,上樣量為60 μg。SDS-PAGE采用8%的分離膠和4%的濃縮膠(丙烯酰胺∶甲叉雙丙烯酰胺=37.5∶l(w/w))。電泳初始電壓為70 V,待蛋白進(jìn)入分離膠后調(diào)整電壓為110 V,至蛋白到達(dá)離凝膠底物5 mm處停止電泳。采用濕法將凝膠上的蛋白條帶轉(zhuǎn)移至PVDF膜上,100 V恒壓冰浴轉(zhuǎn)膜100 min。轉(zhuǎn)膜結(jié)束后將PVDF膜在TBS緩沖液(0.01 mol·L-1Tris、0.15 mol·L-1NaCl,pH 7.5)中清洗3次,每次1 min。封閉液(含0.05% Tween-20,3%牛血清白蛋白的TBS溶液)中室溫封閉1 h。免疫印跡的一抗使用μ-鈣蛋白酶單克隆抗體(Sigma,C0355),稀釋比為1∶300,4℃過(guò)夜孵育;隨后將膜用TBST1溶液(含0.1% Tween-20的TBS溶液)漂洗3次后與二抗(二抗為辣根過(guò)氧化酶標(biāo)記的羊抗鼠IgG,稀釋比為1∶5 000)室溫孵育1 h;孵育結(jié)束后用TBST2溶液(0.05 mol·L-1Tris、0.15 mol·L-1NaCl、0.1% Tween-20,pH 7.5)漂洗PVDF膜3次,ECL法顯色曝光,并使用凝膠成像儀拍照。通過(guò)Phoretix 1D(CLIQS,1.1)軟件分析蛋白免疫印跡條帶的光密度值。
1.2.5 酪蛋白底物酶原分析法 參考VEISETH等[8]的方法并稍作調(diào)整。提取的肌漿蛋白樣品調(diào)整至統(tǒng)一濃度后,按3∶2(v/v)加入樣品緩沖液(150 mmol·L-1Tris-HCl(pH 6.8),20%甘油,0.75%巰基乙醇(MCE),0.02%(w/v)溴酚藍(lán)),混勻后備用。采用12.5%的酪蛋白分離膠(丙烯酰胺∶甲叉雙丙烯酰胺=75∶1(w/w),375 mmol·L-1Tris-HCl(pH 8.8),0.05%的TEMED(w/v),0.05%的APS(w/v),2.1 mg·mL-1酪蛋白)和4%的濃縮膠(丙烯酰胺∶甲叉雙丙烯酰胺=37.5∶l(w/w),125 mmol·L-1Tris-HCl(pH 6.8),0.05%的TEMED(w/v),0.05%的APS(w/v),凝膠厚度為0.75 mm。上樣前在4℃下100 V恒壓預(yù)跑15 min,上樣量為60 μg,相同電泳緩沖液(25 mmol·L-1Tris-HCl、0.05% MCE、192 mmol·L-1甘氨酸、1 mmol·L-1EDTA,pH 8.3)中4℃下100 V恒壓電泳7 h。電泳結(jié)束后更換3次孵育緩沖液(50 mmol·L-1Tris、0.05% MCE、4 mmol·L-1CaCl2,pH 7.5)在室溫下孵育1 h(每次20 min),同種緩沖液中室溫孵育16 h。凝膠經(jīng)考馬斯亮藍(lán)染色后脫色,使用凝膠成像系統(tǒng)照相,所得結(jié)果采用Phoretix 1D(CLIQS,1.1)軟件進(jìn)行分析。
1.3 數(shù)據(jù)處理及分析
采用IBM SPSS 17.0統(tǒng)計(jì)軟件進(jìn)行方差分析,用最小顯著差異法(least significant difference,LSD)和鄧肯氏總重比較法(Duncans Multiple-rang test)進(jìn)行差異顯著性分析。
2. 結(jié)果
2.1 羊肉嫩度分析及分組
根據(jù)宰后24 h及48 h的肌原纖維小片化指數(shù)(MFI值),從采集的50只羊中選出了8只羊分為高、低嫩度兩組,每組各4只羊。MFI值的測(cè)定結(jié)果顯示,宰后24 h及48 h,高嫩度組的MFI值均顯著高于低嫩度組(P<0.05)(表1)。

表1 肌原纖維小片化指數(shù)測(cè)定結(jié)果Table 1 Results of analysis of MFI
2.2 高、低嫩度組羊肉宰后pH的變化
宰后成熟過(guò)程中,高、低嫩度組羊背最長(zhǎng)肌的pH均呈現(xiàn)相同的變化趨勢(shì)(圖1),在達(dá)到極限pH值后,pH值逐漸增加。且在宰后各時(shí)間兩組間的pH值均差異不顯著(P>0.05)。
2.3 μ-鈣蛋白酶免疫印跡分析
宰后成熟過(guò)程中,μ-鈣蛋白酶80 kDa的大亞基逐漸降解(圖2、3)。在宰后7 d內(nèi),隨著μ-鈣蛋白酶被激活并發(fā)揮活性,μ-鈣蛋白酶的大亞基由80 kDa降解為78 kDa,最終降至76 kDa。宰后30 min,高嫩度組μ-鈣蛋白酶80 kDa大亞基含量顯著高于低嫩度組(P<0.05)。宰后6 h,高嫩度組μ-鈣蛋白酶80 kDa大亞基開(kāi)始降解,而低嫩度組μ-鈣蛋白酶80 kDa大亞基在宰后2 h出現(xiàn)明顯降解,但2—24 h基本保持不變。宰后48 h高嫩度組μ-鈣蛋白酶80 kDa大亞基已基本完全降解,而低嫩度組μ-鈣蛋白酶80 kDa大亞基在宰后5 d才基本降解完全。因此,高嫩度組μ-鈣蛋白酶80 kDa大亞基的初始含量高,但降解速度快,導(dǎo)致其蛋白水解活性高,主要在宰后2—48 h內(nèi)發(fā)揮作用。
2.4 μ-/m-鈣蛋白酶酪蛋白底物酶原分析
在酪蛋白底物酶原分析法檢測(cè)肌肉中的μ-/m-鈣蛋白酶時(shí),在孵育過(guò)程使用了4 mmol·L-1的Ca2+,在此濃度條件下,凝膠上所有的μ-/m-鈣蛋白酶均可被激活,發(fā)揮其活性進(jìn)而降解所在條帶位置處的酪蛋白,因此,本試驗(yàn)中,凝膠上檢測(cè)到的為肌肉中尚未發(fā)揮蛋白水解活性/未自溶的μ-/m-鈣蛋白酶的含量。由試驗(yàn)結(jié)果可知,宰后成熟過(guò)程中,高、低嫩度組羊背最長(zhǎng)肌中未自溶的μ-鈣蛋白酶的含量均逐漸降低(圖4、5)。高嫩度組宰后12 h內(nèi)未自溶μ-鈣蛋白酶的含量快速下降,12 h后變化較為緩慢,而低嫩度組樣品中未自溶的μ-鈣蛋白酶在宰后12 h含量仍較高。宰后30 min、2 h高嫩度組未自溶的μ-鈣蛋白酶含量顯著高于低嫩度組(P<0.05),宰后12 h、5 d及7 d低嫩度組未自溶的μ-鈣蛋白酶含量顯著高于高嫩度組(P<0.05)。宰后成熟過(guò)程中,兩組的m-鈣蛋白酶酪蛋白水解活性變化不明顯,m-鈣蛋白酶并未在宰后7 d內(nèi)發(fā)生自溶降解。因此,高嫩度組未自溶的μ-鈣蛋白酶初始含量高,但自溶/失活速度快,導(dǎo)致其活性作用強(qiáng),主要在宰后2—24 h發(fā)揮作用。
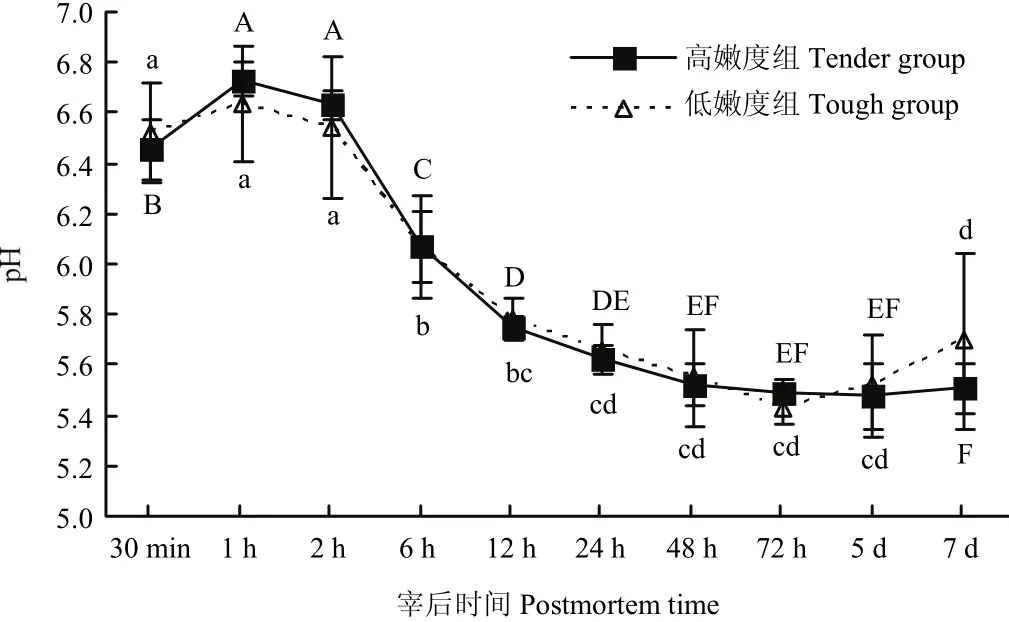
圖1 宰后高、低嫩度組羊肉pH的變化Fig. 1 Changes of pH at postmortem in tender/tough groups
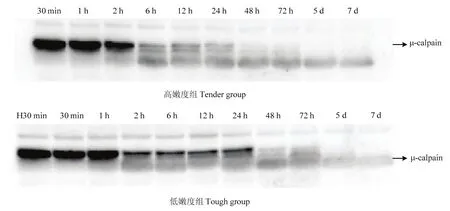
圖2 宰后高、低嫩度組羊肉中μ-鈣蛋白酶的免疫印跡成像Fig. 2 Western blots profiles showing the state of μ-calpain at postmortem in tender/tough groups
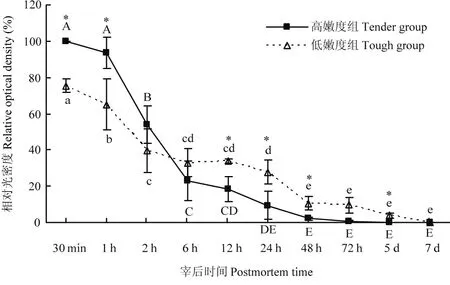
圖3 宰后高、低嫩度組羊肉中μ-鈣蛋白酶80 kDa大亞基的變化Fig. 3 Changes of 80 kDa large subunit of μ-calpain at postmortem in tender/tough groups
3 討論
3.1 羊肉嫩度分析
MFI值是反映宰后肌原纖維蛋白、細(xì)胞骨架蛋白降解程度的一個(gè)重要標(biāo)志[11],能體現(xiàn)宰后I帶關(guān)鍵結(jié)構(gòu)蛋白的降解及肌纖維連接作用的削弱程度,與宰后肌肉的嫩度顯著正相關(guān)[12],是反映宰后肌肉嫩度的主要指標(biāo)之一。本研究測(cè)定了50只羊宰后24 h和48 h背最長(zhǎng)肌的MFI值,并對(duì)結(jié)果進(jìn)行了排序。選擇了24 h及48 h MFI值高和低的羊各4只作為高、低嫩度組的樣品。試驗(yàn)結(jié)果表明,兩組羊背最長(zhǎng)肌樣品的嫩度具有顯著差異,可用于后續(xù)試驗(yàn)的進(jìn)一步分析。
3.2 不同嫩度羊肉宰后pH的變化
宰后肌肉的糖酵解作用消耗ATP并產(chǎn)生乳酸,乳酸的累積使得肌肉的pH逐漸下降,當(dāng)乳酸含量趨于穩(wěn)定后,pH也逐漸趨于穩(wěn)定。MARSH等[13-14]研究表明,宰后pH的下降速率影響宰后肌肉的嫩度。此外,pH也是μ-鈣蛋白酶活性和自溶程度的重要影響因素[15],許多研究證實(shí),宰后pH下降速度快的肌肉中μ-鈣蛋白酶失活的速率也快[16-18]。CLAEYS等[19]在宰后低pH的肌肉中檢測(cè)到的μ-鈣蛋白酶活性也較低。POMPONIO等[20]在研究宰后早期豬肌肉中pH的下降對(duì)鈣蛋白酶活性的影響時(shí)發(fā)現(xiàn),宰后μ-鈣蛋白酶自溶及失活的速度與pH的下降速度呈高度正相關(guān)性,且pH的下降速度快,肌原纖維小片化的速度也快。本研究中,高、低嫩度兩組在宰后7 d內(nèi)pH的變化趨勢(shì)相同,且各時(shí)間點(diǎn)pH差異不顯著,說(shuō)明pH并不是造成兩組樣品嫩度差異的原因,宰后高、低嫩度兩組樣品中鈣蛋白酶的生理生化狀態(tài)也并不受pH的干擾。在本試驗(yàn)中,排除了pH對(duì)嫩度及鈣蛋白酶的影響作用。
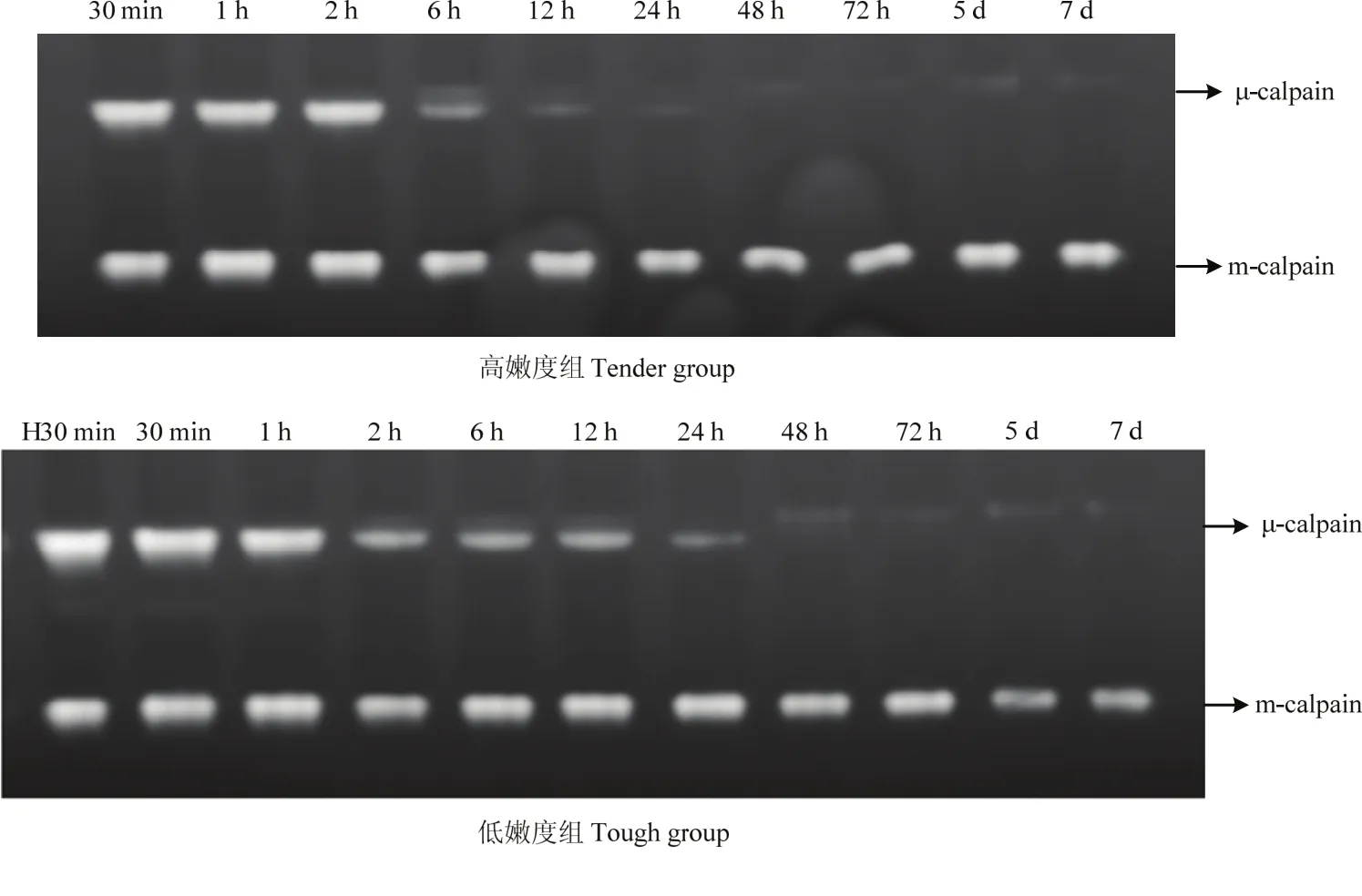
圖4 宰后高、低嫩度組羊肉中μ-/m-鈣蛋白酶酪蛋白底物酶原圖譜Fig. 4 Casein zymography profiles presenting the μ-/m-calpain at postmortem in tender/tough groups
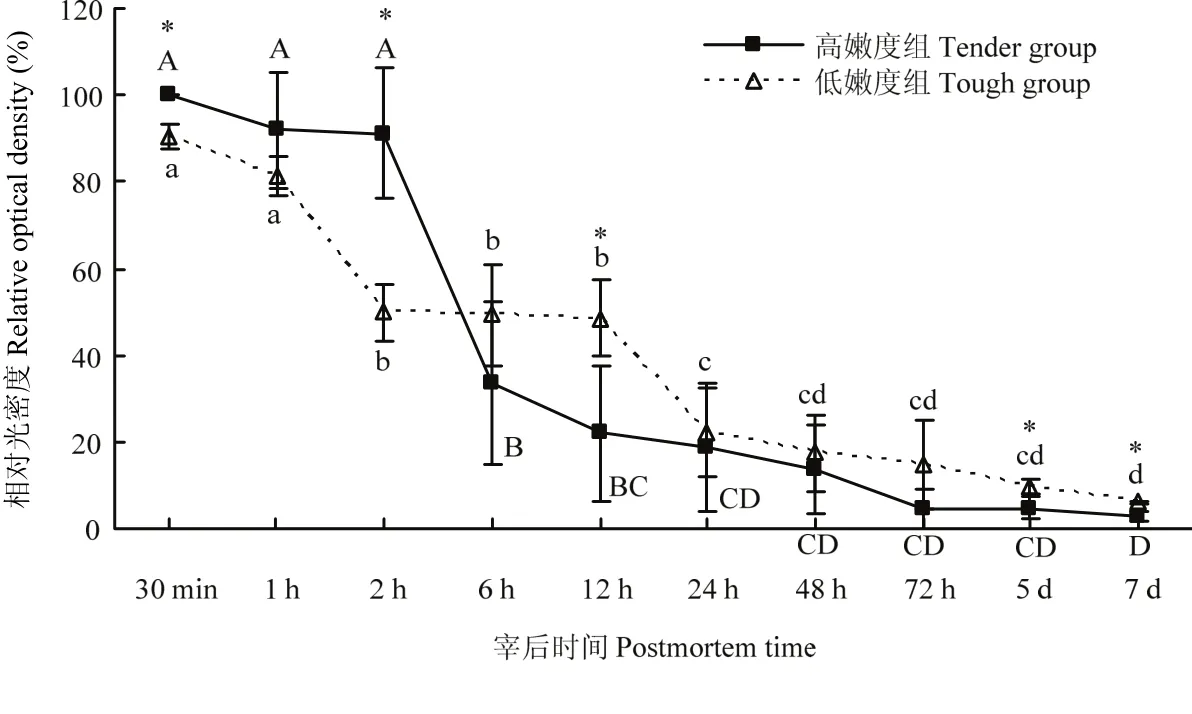
圖5 宰后高、低嫩度組羊肉中未自溶μ-鈣蛋白酶含量的變化Fig. 5 Changes of the content of native μ-calpain at postmortem in tender/tough groups
3.3 不同嫩度羊肉宰后μ-/m-鈣蛋白酶的差異
研究表明,宰后早期μ-鈣蛋白酶的自溶與活性的激活及早期肌原纖維蛋白的降解有關(guān)[6],宰后大部分蛋白質(zhì)的降解及肌肉嫩化進(jìn)程由μ-鈣蛋白酶的激活引起,μ-鈣蛋白酶自溶的速度快,會(huì)更早的被激活并較快的失活。YUAN等[21]通過(guò)蛋白質(zhì)免疫印跡法檢測(cè)羊宰后成熟過(guò)程中μ-鈣蛋白酶的自溶情況發(fā)現(xiàn),μ-鈣蛋白酶的自溶和激活程度與宰后蛋白降解有關(guān),自溶程度越大,肌原纖維蛋白的降解程度也越大。CRUZEN等[22]在不同肌肉宰后蛋白降解的研究中發(fā)現(xiàn),蛋白水解程度低的肌肉中μ-鈣蛋白酶完整的80 kDa大亞基的存在比例高,肌肉的嫩度差。本試驗(yàn)通過(guò)對(duì)比宰后高、低嫩度兩組樣品μ-鈣蛋白酶80 kDa大亞基的變化情況,明確μ-鈣蛋白酶的激活狀態(tài)及活性與羊肉嫩度的關(guān)系。由試驗(yàn)結(jié)果可知,宰后30 min,高嫩度組μ-鈣蛋白酶80 kDa大亞基的含量顯著高于低嫩度組,說(shuō)明高嫩度組羊肉中μ-鈣蛋白酶80 kDa大亞基的初始總量高。但隨著宰后時(shí)間的延長(zhǎng),高嫩度組μ-鈣蛋白酶自溶的速率顯著高于低嫩度組,宰后6 h高嫩度組μ-鈣蛋白酶出現(xiàn)明顯的78、76 kDa的降解條帶,在宰后12 h,高嫩度組μ-鈣蛋白酶80 kDa大亞基的含量顯著低于低嫩度組,表明高嫩度組μ-鈣蛋白酶的激活程度及活性都明顯高于低嫩度組。
在酪蛋白底物酶原分析法對(duì)高、低嫩度組羊肉中未自溶μ-/m-鈣蛋白酶含量的檢測(cè)中發(fā)現(xiàn),高、低嫩度兩組羊背最長(zhǎng)肌樣品中未自溶的μ-鈣蛋白酶的含量隨宰后時(shí)間的延長(zhǎng)均呈下降趨勢(shì),表明μ-鈣蛋白酶在宰后成熟過(guò)程中通過(guò)自溶逐漸發(fā)揮其嫩化作用。研究表明,μ-鈣蛋白酶對(duì)肌肉的嫩化作用主要發(fā)生在宰后前3 d,μ-鈣蛋白酶因自溶而被激活,在發(fā)揮蛋白水解活性的同時(shí),自身會(huì)降解失活,最終完全喪失活性。但宰后肌肉中Ca2+的濃度達(dá)不到m-鈣蛋白酶激活所需的水平,故m-鈣蛋白酶并不發(fā)揮作用[23-25]。O’HALLORAN等[26]研究表明,宰后3 h μ-鈣蛋白酶活性低的肌肉更嫩,骨骼肌蛋白水解的程度更大,即μ-鈣蛋白酶自溶失活速度快的肌肉嫩度更好。宰后30 min,高嫩度組未自溶μ-鈣蛋白酶的含量顯著高于低嫩度組,而宰后6 h,低嫩度未自溶μ-鈣蛋白酶的含量高,高嫩度組μ-鈣蛋白酶出現(xiàn)明顯的自溶條帶。至宰后12 h時(shí),高嫩度組未自溶μ-鈣蛋白酶含量顯著低于低嫩度組,說(shuō)明高嫩度組μ-鈣蛋白酶自溶并發(fā)揮活性的速率更快,對(duì)肌肉嫩化的作用更為突出,這與蛋白質(zhì)免疫印跡法檢測(cè)到的宰后高、低嫩度組中μ-鈣蛋白酶降解發(fā)揮蛋白水解活性的變化結(jié)果一致。在宰后7 d內(nèi),高、低嫩度兩組樣品中m-鈣蛋白酶的活性沒(méi)有明顯變化,也未出現(xiàn)自溶降解現(xiàn)象,表明m-鈣蛋白酶并未在這一過(guò)程中發(fā)揮作用。本試驗(yàn)中,高、低嫩度組嫩度的差異與m-鈣蛋白酶的活性無(wú)關(guān)。
本研究中,高、低嫩度組嫩度的差異與宰后pH的變化無(wú)關(guān),兩組樣品在宰后的貯藏溫度及環(huán)境相同,表明溫度和pH并不是引起兩組樣品嫩度差異的原因,也排除了溫度及pH對(duì)高、低嫩度組整體μ-鈣蛋白酶活性差異的影響。但在本研究中,高、低嫩度組μ-鈣蛋白酶自溶激活的初始時(shí)間不同,高嫩度組μ-鈣蛋白酶在宰后2—6 h變化顯著,而低嫩度組在宰后1—2 h內(nèi)已開(kāi)始有明顯變化,根據(jù)pH的測(cè)定結(jié)果可知,雖然宰后不同成熟時(shí)間高、低嫩度兩組樣品的pH均差異不顯著,但宰后1 h及2 h低嫩度組樣品的pH較低,可能較早的激活了μ-鈣蛋白酶,造成兩組樣品μ-鈣蛋白酶激活起始時(shí)間存在差異。但總體來(lái)看,高嫩度組的含量高于低嫩度組,其自溶及激活的程度卻更大,引起兩組樣品中活性差異的主要調(diào)控因子仍需進(jìn)一步研究確定。
4 結(jié)論
宰后高、低嫩度組羊肉中μ-鈣蛋白酶80 kDa大亞基的存在狀態(tài)及未自溶的μ-鈣蛋白酶含量存在顯著差異,高嫩度組μ-鈣蛋白酶的自溶和激活程度顯著高于低嫩度組。宰后30 min,高嫩度組初始的μ-鈣蛋白酶含量高于低嫩度組,但隨著宰后時(shí)間的延長(zhǎng),其降解的程度也快于低嫩度組。高、低嫩度組樣品中的μ-鈣蛋白酶在宰后6 h及12 h的差異最為顯著。宰后24 h內(nèi),尤其是12 h內(nèi),μ-鈣蛋白酶的變化狀態(tài)對(duì)羊肉嫩度的影響最大。
[1] TAYLOR R G, GEESINK G H, THOMPSON V F, KOOHMARAIE M, GOLL D E. Is Z-disk degradation responsible for postmortem tenderization? Journal of Animal Science, 1995, 73(5): 1351-1367.
[2] LIU J, RUUSUNEN M, PUOLANNE E, ERTBJERG P. Effect of pre-rigor temperature incubation on sarcoplasmic protein solubility, calpain activity and meat properties in porcine muscle. LWT-Food Science and Technology, 2014, 55(2): 483-489.
[3] GOLL D E, THOMPSON V F, LI H Q, WEI W, CONG J Y. The calpain system. Physiological Reviews, 2003, 83(3): 731-801.
[4] ZIMMERMAN U P, SCHLAEPFER W W. Two-stage autolysis of the catalytic subunit initiates activation of calpain I. Biochimicae BiophysicaActa, 1991, 1078(2):192-198.
[5] ZHANG W G, LONERGAN S M, GARDNER M A, HUFFLONERGAN E. Contribution of postmortem changes of integrin, desmin and μ-calpain to variation in water holding capacity of pork. Meat Science, 2006, 74(3): 578-585.
[6] MELODY J L, LONERGAN S M, ROWE L J, HUIATT T W,MAYES M S, HUFF-LONERGAN E. Early postmortem biochemical factors influence tenderness and water-holding capacity of three porcine muscles. Journal of Animal Science, 2004, 82(4): 1195-1205.
[7] GEESINK G H, KOOHMARAIE M. Postmortem proteolysis and calpain/calpastatin activity in callipyge and normal lamb biceps femoris during extended postmortem storage. Journal of Animal Science, 1999, 77(6): 1490-1501.
[8] VEISETH E, SHACKELFORD S D, WHEELER T L, KOOHMARAIE M. Effect of postmortem storage on mu-calpain and m-calpain in ovine skeletal muscle. Journal of Animal Science, 2001, 79(6): 1502-1508.
[9] CULLER R A, PARRISH F C, SMITH G C, CROSS H R. Relationship of MFI to certain chemical, physical, and sensory characteristics of bovine LD muscle. Journal of Food Science, 1978, 43: 1177-1180.
[10] LOMIWES D, FAROUK M M, WU G, YOUNG O A. The development of meat tenderness is likely to be compartmentalized by ultimate pH. Meat Science, 2014, 96(1): 646-651.
[11] LI K, ZHANG Y, MAO Y, CORNFORTH D, DONG P, WANG R, ZHU H, LUO X. Effect of very fast chilling and ageing time on ultra-structure and meat quality characteristics of Chinese yellow cattle M. longissimus lumborum. Meat Science, 2012, 92(4): 795-804.
[12] REES M P, TROUT G R, WARNER R D. The influence of the rate of pH decline on the rate of ageing for pork. II: Interaction with chilling temperature. Meat Science, 2003, 65(2): 805-818.
[13] MARSH B B, RINGKOB T P, RUSSELL R L, SWARTZ D R, PAGEL L A. Effects of early-postmortem glycolytic rate on beef tenderness. Meat Science, 1987, 21(4): 241-248.
[14] MARSH B B, RINGKOB T P, RUSSELL R L, SWARTZ D R, PAGEL L A. Mechanisms and strategies for improving meat tenderness//Proceedings of the 41st Annual Reciprocal Meat Conference, 1989: 113-121.
[15] MADDOCK K R, HUFF-LONERGAN E, ROWE L J, LONERGAN S M. Effect of pH and ionic strength on calpastatin inhibition of μand m-calpain. Journal of Animal Science, 2005, 83(6): 1370-1376.
[16] HWANG I H, THOMPSON J M. The interaction between pH and temperature decline early postmortem on the calpain system and objective tenderness in electrically stimulated beef longissimus dorsi muscle. Meat Science, 2001, 58(2): 167-174.
[17] RHEE M S, RYU Y C, KIM B C. Postmortem metabolic rate and calpain system activities on beef Longissimus tenderness classifications. Bioscience Biotechnology and Biochemistry, 2006, 70(5): 1166-1172.
[18] WHIPPLE G, KOOHMARAIE M, DIKEMAN M E, CROUSE J D. Effects of high temperature conditioning on enzymatic-activity and tenderness of bos-indicus longissimus muscle. Journal of Animal Science, 1990, 68(11): 3654-3662.
[19] CLAEYS E, DE SMET S, DEMEYER D, GEERS R, BUYS N. Effect of rate of pH decline on muscle enzyme activities in two pig lines. Meat Science, 2001, 57(3): 257-263.
[20] POMPONIO L, LAMETSCH R, KARLSSON A H, COSTA L N, GROSSI A, ERTBJERG P. Evidence for post-mortem m-calpain autolysis in porcine muscle. Meat Science, 2008, 80(3): 761-764.
[21] YUAN H B K, LUC G, ROSENVOLD K. Pre rigor processing, ageing and freezing on tenderness and colour stability of lamb loins. Meat Science, 2013, 95(2): 412-418.
[22] CRUZEN S M, PEDRO V R P, LONERGAN S M, HUFFLONERGAN E. Postmortem proteolysis in three muscles from growing and mature beef cattle. Meat Science, 2014, 96(2): 854-861.
[23] GEESINK G H, KUCHAY S, CHISHTI A H, KOOHMARAIE M. μ-Calpain is essential for postmortem proteolysis of muscle proteins. Journal of Animal Science, 2006, 84(10): 2834-2840.
[24] KOOHMARAIE M. The role of Ca2+-dependent proteases (calpains) in postmortem proteolysis and meat tenderness. Biochemistry, 1992, 74(3): 239-245.
[25] KOOHMARAIE M, GEESINK G H. Contribution of postmortem muscle biochemistry to the delivery of consistent meat quality with particular focus on the calpain system. Meat Science, 2006, 74(1): 34-43.
[26] O’HALLORAN G R, TROY D R, BUCKLEY D J, REVILLE W J. The role of endogenous proteases in the tenderization of fast glycolysing muscle. Meat Science, 1997, 47(3): 187-200.
(責(zé)任編輯 趙伶俐)
Analysis and Comparison of Calpain in Mutton with Different Levels of Tenderness
DU Man-ting, LI Xin, LI Zheng, GAO Xing, ZHANG Cai-xia, ZHANG De-quan
(Institute of Food Science and Technology, Chinese Academy of Agricultural Sciences/Key Laboratory of Agro-Products Processing, Ministry of Agriculture, Beijing 100193)
【Objective】Calpain plays an important role in postmortem tenderization through the proteolysis ability. The activity of calpains, especially μ-calpain, is closely related to the tenderness of muscles. The objective of this study was to investigate the activation process and the autolysis rate of μ-calpain in mutton during postmortem aging by analyzing and comparing the differences of calpain in mutton with different levels of tenderness, thereby provide a theoretical basis for regulating tenderness through regulating calpain activity. 【Method】 The longissimus dori of WuZhuMuQin Fat Tailed sheep were divided into tender group and tough group based on myofibrillar fragmentation index, then the pH value, the existent state of μ-calpain large subunit and the content of native μ-/m-calpain during postmortem were analyzed. 【Result】The variation trend of pH at postmortem in tender/tough group was identical, all the pH values in two groups at the same postmortem time showed no difference. The 80 kDa subunit of μ-calpain degraded gradually after slaughter. The content of μ-calpain 80 kDa subunit of tender group was significantly higher than tough group at 30 min postmortem. The intact 80 kDa subunit of μ-calpain could not be detected after 48 h in tender group, while it could be detected until 5 d postmortem in tough group. The content of native μ-calpain was decreased gradually, and this content oftender group was significantly higher than tough group at 30 min, 2 h postmortem, however, at 12 h, 5 d, and 7 d postmortem the tough group was significantly higher than the tender group. The autolysis and degradation of μ-calpain in tender group was apparent at 6 h postmortem.【Conclusion】Compared with tough group, tender group had a higher initial μ-calpain content, but the autolysis and degradation rate of μ-calpain in tender group was much faster, leading higher autolysis and degradation degree in tender group. The results indicated that meat tenderness during the postmortem was directly affected by μ-calpain expression quantity and autolysis. In addition, changes of μ-calpain during 24 h postmortem, especially 12 h, had the most impact on mutton tenderness.
mutton; muscle; tenderness; μ-calpain; postmortem aging
2016-01-22;接受日期:2016-04-07
國(guó)家自然科學(xué)基金面上項(xiàng)目(31471604)、國(guó)家農(nóng)業(yè)科技創(chuàng)新工程
聯(lián)系方式:杜曼婷,Tel:010-62816474;E-mail:mantingdu@126.com。通信作者張德權(quán),Tel:010-62818740;E-mail:dequan-zhang0118@126.com
