中國兩棲動物物種豐富度省級地理分布格局及其與氣候因子的關系
2016-12-09 05:13:55沈夢偉畢孟杰敬琴陳文德陳圣賓
四川動物 2016年1期
沈夢偉, 畢孟杰, 敬琴, 陳文德, 陳圣賓
(1. 成都理工大學旅游與城鄉規劃學院,成都610059;2. 成都理工大學地球科學學院,成都610059; 3. 環境保護部南京環境科學研究所,南京210042)
?
中國兩棲動物物種豐富度省級地理分布格局及其與氣候因子的關系
沈夢偉1, 畢孟杰2, 敬琴1, 陳文德1, 陳圣賓3*
(1. 成都理工大學旅游與城鄉規劃學院,成都610059;2. 成都理工大學地球科學學院,成都610059; 3. 環境保護部南京環境科學研究所,南京210042)
物種豐富度分布格局及其形成機制的研究對于生物多樣性保護具有重要意義。為了解中國兩棲動物物種豐富度分布格局,本文利用中國省級尺度兩棲動物物種分布數據和環境信息,結合GIS和數理統計方法,探討兩棲動物物種豐富度的地理分布格局與環境因子之間的關系。研究結果表明:(1)物種豐富度隨緯度增加呈逐漸遞減趨勢,但缺乏顯著的經度梯度。豐富度最高的地區主要集中在南方,我國北方、西北干旱區和青藏高原北部地區豐富度較低;(2)最優模型由年均溫、最冷月均溫、凈初級生產力、年降水量變化范圍、月均降水量標準差組成,多層次方差分解表明,最冷月均溫的獨立解釋能力(17.6%)高于年均溫(11.5%);(3)方差分解表明,季節性因子的獨立解釋能力(5.6%)低于熱量因子(6.1%),但高于水分因子(4.5%),因此我們認為季節性因子也是限制中國兩棲動物分布的重要因素。
生物地理學;空間分布格局;氣候;宏生態學
物種豐富度的大尺度地理格局是宏生態學和生物地理學的中心議題之一,也是全球及區域物種多樣性保護的重要參考和依據(Brown & Lomolino,2000;Gaston,2000)。物種豐富度在地理尺度上的變化及其與生物和非生物因素的關系,是理解物種地理分布和預測氣候變化對生物多樣性影響的重要基礎(Chown & Gaston,2010;Comontetal.,2012)。
一定時空內物種的組成及數量受生物進化、地質歷史、生態過程和當前環境的共同制約,其中環境因素如何影響物種豐富度的地理格局一直是研究的熱點。以往關于物種豐富度大尺度地理格局的研究涉及各主要生物類群,包括植物(Currie & Paquin,1987;Francis & Currie,2003;Chenetal.,2011)、無脊椎動物(White & Kerr,2006;Chenetal.,2014)、魚類(Zhaoetal.,2006)、兩棲類(Qian,2007;Huetal.,2012)、爬行類(Qianetal.,2007)、鳥類(Dingetal.,2006;Ormeetal.,2006)、哺乳類(Tognelli & Kelt,2004;Tognelli,2005)等,并且圍繞地理格局的成因提出了多個假說,包括面積假說(Rahbek & Graves,2000;Sandersetal.,2007)、能量假說(Wright,1983)、氣候穩定性假說(Klopfer,1959)、生境異質性假說(Kerr & Packer,1997)、歷史假說(Latham & Ricklefs,1993;Qian & Ricklefs,2000)等。這些假說是基于不同的影響因子,來探討大尺度上物種豐富度分布格局形成的機制。雖然各個假說在不同生物類群上進行了驗證,但是對于形成物種豐富度大尺度格局的主導因子,目前仍存在廣泛爭議(Rosenzweig,2001)。
基于環境因子的能量假說、氣候穩定性假說和生境異質性假說經常被認為是制約物種豐富度地理格局的主導因子,并受到更多的關注(Rosenzweig,2001;王志恒等,2009)。能量假說認為,物種豐富度主要受能量控制,能量越高則物種豐富度越高(Evansetal.,2005)。根據能量的不同形式,能量假說包括環境能量假說、生態學代謝假說、生產力假說、水分-能量動態假說以及寒冷忍耐假說(王志恒等,2009;陳勝東等,2011)。氣候穩定性假說認為,穩定的氣候環境能促進物種的特化,并使其生態位區域狹窄,因此環境能容納更多的物種;相反在波動的氣候環境下,物種需要更廣泛的生理機能才能生存下來,因此物種豐富度較低(Stevens,1989)。生境異質性假說認為,生境異質性高的地區能夠提供更多的生態位,也更有利于物種共存,因此物種豐富度也隨之增加(Kerr & Packer,1997;Büchietal.,2009)。
兩棲動物起源于距今3.5億年前的總鰭魚類,隸屬脊索動物門Chordata脊椎動物亞門Vertebrata兩棲綱Amphibia。現存兩棲動物分為蚓螈目Gymnophiona、有尾目Caudata和無尾目Anura。全球共有兩棲動物約73科7426種(Amphibian species of the world,2015)。根據《中國兩棲動物彩色圖鑒》,記錄我國兩棲動物共370種(亞種),隸屬11科64屬(費梁等,2010)。由于兩棲動物同時可以在水中和陸地生活,這種特殊的生物學和生態學特征使其經常成為生態監測的重要類群(Stuartetal.,2004),因而對于研究環境污染和氣候變化有重要作用。并且研究大尺度兩棲動物物種豐富度的格局及其氣候制約因素對于生物多樣性的保護和生態系統管理具有重要意義。
本文在省級尺度上探討中國兩棲動物物種豐富度的地理格局及其與氣候因子,特別是溫度(熱量)的關系。由于兩棲動物屬于冷血動物,其生命活動所需的能量大部分來源于外部環境,因此,我們由此提出第1個假設:相比年水分因子,熱量因子對兩棲動物物種豐富度的空間分異影響更大;根據寒冷忍耐假說,物種難以在寒冷的冬季生存,因此,提出第2個假設:相比年均溫,最冷月均溫對物種分布的影響更大;根據氣候穩定性假說,本文提出第3個假設,相比溫度和水分因子,季節性因子同樣對兩棲動物的地理分布有重要影響。
1 材料與方法
1.1 研究概況
本研究區域范圍包括中國34個省、自治區、直轄市以及香港和澳門2個特別行政區。由于海陸熱力性質差異顯著,除青藏高原和西北內陸地區之外,中國大部分地區受來自太平洋的東南季風和來自印度洋的西南季風影響(馮建孟,徐成東,2009)。中國南北氣候差異主要表現在溫度方面,而水分差異在東西方向更為明顯。從南到北,中國的地帶性植被帶依次為熱帶雨林季雨林帶、亞熱帶常綠闊葉林帶、暖溫帶落葉闊葉林帶、溫帶針闊混交林帶和寒溫帶針葉林帶;從東向西,植被帶依次為森林、森林草原植被帶、草原植被帶和荒漠半荒漠植被帶(方精云,2001)。
1.2 數據來源
本文以費梁等(2010)編寫的《中國兩棲動物彩色圖鑒》為藍本,對照《中國兩棲動物志》、中國動物數據庫和近年來發表的相關文獻。總共錄入3目11科64屬394種兩棲動物,包括亞種。以“1”或“0”記錄某種兩棲動物在某省份的分布與否,建立物種分布數據庫,進而計算各省物種豐富度。由于部分直轄市和特別行政區面積較小,將北京市和天津市并入河北省;將上海市并入浙江省;將香港和澳門特別行政區并入廣東省(圖1)。不同區域研究人員的數量和科研水平可能影響對兩棲動物豐富度的估計,但由于相關數據的限制,本研究并未對此進行評估。

圖1 中國省級行政區尺度兩棲動物物種豐富度
Fig. 1 The geographic pattern of amphibian species richness at provincial scale in China
注: 北京市和天津市并入河北省, 將上海市并入浙江省, 重慶市并入四川省, 香港和澳門特別行政區并入廣東省。
Notes: Hebei province was merged with Beijing and Tianjin, Zhejiang province was merged with Shanghai, Sichuan province was merged with Chongqing, and Guangdong province was merged with Hong Kong and Macau.
本研究共采集了代表熱量、降水和季節性的12個氣候因子(表1):熱量因子(energy availability)包括年均溫(mean annual temperature,TEM)、最冷月均溫(mean temperature of the coldest month,TEMmin)、最暖月均溫(mean temperature of the warmest month,TEMmax)和凈初級生產力(net primary productivity,NPP);水分因子(water availability)包括年均降水量(annual precipitation,PREC)、夏季降水量(annual precipitation in summer,PRECsum)、水分虧缺指數(water deficit,WD;即潛在蒸散量與實際蒸散量的差值)和濕潤指數(moisture index,MI;即蒸散量/實際蒸散量);季節性因子(seasonality)包括年溫度變化范圍(annual temperature range,TEMvar;即TEMmax和TEMmin的差值)、月均溫標準差(the standard deviation of mean monthly temperature,TEMsd)、年降水量變化范圍(annual precipitation range,PRECvar,即最大和最小月均降水量的差值)、月均降水量標準差(the standard deviation of mean monthly precipitation,PRECsd)。NPP數據來源于http://atlas.sage.wisc.edu/,其他氣候數據來源于WorldClim(http://www.worldclim.org),利用Arcgis 9.3將中國矢量地圖切割成3844個0.5°像元,提取每個像元中心點的經緯度,運用DIVA-GIS軟件,獲取每個中心點的氣候數據。各省內各像元氣候因子的均值代表其平均氣候條件。

表1 中國省級行政區尺度兩棲動物物種豐富度與每個氣候因子的線性回歸關系
1.3 統計方法
為了降低空間自相關,本文采用Moran’sI檢驗數據在不同空間距離上的空間自相關。取值范圍在-1到1之間,正值表示正相關,負值表示負相關,0則表示空間格局是隨機的。
本文首先采用基于普通最小二乘法(ordinary least squares,OLS)的一元線性回歸模型考察各氣候因子對兩棲動物豐富度的解釋能力。然后根據Akaike信息量準則(Akaike information criterion,AIC)篩選出最優模型(AIC值最小的1個因子)及其所包含的變量(Legendre & Legendre,1998)。
為了解決自變量共線的問題,本文采用了方差分解(variation partitioning)(Borcardetal.,1992)和多層次方差分解(hierarchical partitioning)(Mac Nally,2000)。這2種方法不能給出擬合預測的方程式,但能夠幫助理解可能的因果關系和各自變量的解釋能力。運用方差分解和多層次方差分解把全部能夠解釋的方差分解成各個因子共同作用部分和獨立作用部分,獨立作用部分的大小表明某一個(類)因子的相對重要性(Mac Nally,2000)。
為了滿足數據的正態性,將各省物種數進行對數轉換。以上統計在SAM(spatial analysis in macroecology)和R (R Development Core Team, 2013)中完成。
2 結果
2.1 物種豐富度與經緯度的關系
本文共統計3目11科64屬394種兩棲動物,包括亞種。其中物種最多的目為無尾目,有326種,占總物種的82.7%,其次為有尾目67種,占17%,而蚓螈目僅1種,占0.3%;物種最多的科為蛙科Ranidae,有131種,占總物種的33.2%,其次為角蟾科Megophryidae 85種,占21.6%和樹蛙科Rhacophoridaae 55種,占14%;物種最多的屬為角蟾屬Megophrys,有31種,占總物種的7.9%,其次為樹蛙屬Rhacophorus26種(6.6%)和臭蛙屬Odorrana25種(6.3%)。
由圖1可知,物種豐富度熱點主要集中在南方,而北方、西北干旱區和青藏高原的北部(青海)等省物種豐富度較低(圖1)。物種豐富度最高的為云南省,有131種,其次為四川、廣西、貴州、湖南,物種數均在70種以上。物種豐富度最低的為寧夏,僅有6種,其次為新疆、青海、內蒙古,其物種數均低于10種。兩棲動物物種豐富度沒有明顯的經度梯度(圖2:a)(R2=0.0085,P>0.05),而在緯度梯度上空間變異明顯,即隨緯度增加,物種豐富度顯著降低(圖2:b)(R2=0.6286,P<0.001)。
2.2 物種豐富度與氣候的關系
簡單線性回歸表明,物種豐富度除了與WD、TEMvar和TEMsd呈負相關,與其余氣候因子均呈正相關(表1)。在熱量因子中,NPP(R2=0.639,P<0.001)對物種豐富度的影響最大,這也是所有因子中對物種豐富度影響最大的一個,其次為TEMmin(R2=0.522,P<0.01)和TEM(R2=0.365,P<0.05)。

圖2 中國省級行政區尺度兩棲動物物種豐富度與經度(a)和緯度(b)的關系
在水分因素中,PREC對物種豐富度的影響最大(R2=0.487,P<0.001),其次為MI(R2=0.409,P<0.05)和PRECsum(R2=0.474,P<0.001)。在季節性因子中,TEMvar(R2=0.564,P<0.001)對物種豐富度的影響最大,其次為TEMsd(R2=0.557,P<0.001)和PRECsd(R2=0.420,P<0.001)。
根據AIC共篩選出4095種模型,其中最優模型由TEM、TEMmin、NPP、PRECvar和PRECsd這5項組成,AIC值為0.795,能夠解釋豐富度空間變異的77.7%(表2)。將最優模型中因子進行多層次方差分解(圖3),結果表明,NPP的獨立解釋能力和總解釋能力最強,分別能夠解釋模型方差的26.3%和63.9%;其次為TEMmin、PRECsd、TEM和PRECvar,分別能夠解釋方差的17.6%和52.2%、12.3%和42.0%、11.5%和36.5%以及10.0%和34.3%。模型的殘差沒有明顯的空間屬性,這可以通過Moran’sI在不同距離上的顯著性和殘差與經緯度間的關系看出(圖4)。豐富度Moran’sI指數在較短的距離

表2 兩棲動物物種豐富度-氣候最優模型參數估計

圖3 兩棲動物物種豐富度主要影響因子的獨立作用部分、交互作用部分

圖4 不同空間距離上物種豐富度(實線)和模型殘差(虛線)的Moran’s I
上呈正相關,在較長的距離上呈負相關;模型殘差Moran’sI指數在較短的距離上呈負相關,之后又呈正相關,而在較長的距離上呈負相關并趨近于0,這表明所構建的模型能夠很好地解釋中國兩棲動物物種豐富度格局,模型的殘差主要是一些隨機變化和取樣誤差。
方差分解結果顯示,熱量因子中,TEM、TEMmin、TEMmax和NPP這4項能夠解釋全部方差的76.6%;水分因素中,PREC、PRECsum、WD和MI這4項能夠解釋全部方差的51.7%;季節性因子中,TEMvar、TEMsd、PRECvar和PRECsd這4項能夠解釋全部方差的62.7%。熱量因子與季節性因子的交互作用最大為59.0%,其次是熱量因子與水分因子之間的交互作用為49.1%,而水分因子和季節性因子之間的交互作用為37.6%。三者之間的交互作用為37.6%。熱量因子的獨立作用最大,為6.1%,其次為季節性因子,為5.6%,水分因子的獨立解釋作用僅為4.5%(圖5)。
3 討論
最優模型和方差分解的結果均表明,相比于水分,熱量因子對兩棲動物豐富度分布格局的影響更大,這一點也與前人的研究結果類似(Allenetal.,2002;Hawkinsetal.,2003;馮建孟,徐成東,2009),同時也驗證了本文的第1個假設。最優模型共包含5個變量,其中有3個與熱量相關的因子,并且這3個因子能夠解釋物種豐富度分布格局的76.6%。從方差分解的結果來看,熱量因子的總解釋率和獨立解釋率分別為76.6%和6.1%,高于水分因子的51.7%和4.5%。熱量影響著動物的各個方面,如體型、行為和進化進程等。兩棲類動物在溫度較高的地區可以擁有更多的時間覓食、繁殖,更快的生長速度和更高的存活率(Gotthardtetal.,2000;Chown & Gaston,2010)。Allen(2002)認為,植物物種分布格局主要受水分和熱量共同作用的影響,而動物主要受熱量的影響,水分的作用相對小一些。在動物之中,恒溫動物的體溫不隨外界溫度的變化而變化,因此,其豐富度分布格局受到生產力的影響。變溫動物體溫受外界環境的影響較大,因此,在溫度較高的區域,其新陳代謝更快,進而縮短世代時間,最終提高物種形成的速率和物種多樣性。
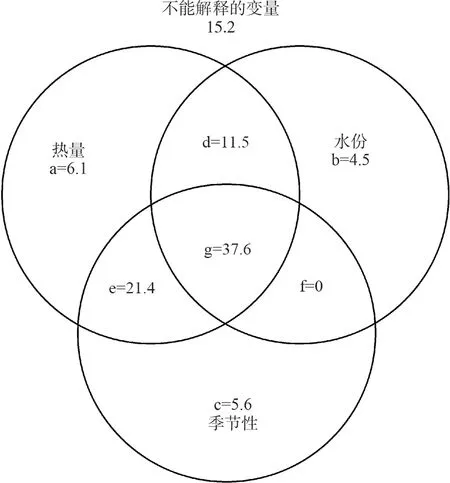
圖5 影響兩棲動物物種豐富度的各因子方差分解結果圖示
a、b和c為熱量、水分和季節性因子的獨立解釋作用;d、e、f和g表示它們之間的交互作用。
Variation of amphibian richness explained by three sets of variables: energy, water, and seasonality, and the unexplained variation; a, b, and c are unique effects of energy, water, and seasonality, respectively; while d, e, f and g indicates their joint effects.
簡單線性回歸表明TEMmin(R2=0.522,P<0.001)的解釋能力高于TEM(R2=0.365,P<0.001)。并且多層次方差結果顯示,TEMmin的獨立解釋作用為17.6%,總解釋作用為52.2%,而TEM的獨立解釋為11.5%,總解釋作用為36.5%。與TEM相比,TEMmin對兩棲動物物種豐富度分布格局的影響更大。寒冷忍耐假說認為,很多物種由于不能忍受冬季的低溫而無法生存。因此,隨著冬季溫度的降低,物種多樣性也隨之減少(Hawkinsetal.,2003),這里的冬季溫度通常是指最冷月平均溫度,一些生態學家也將這一假說稱為“低溫限制假說”,許多研究結果也證明了這一假說。如Sakai和Weiser(1973)研究發現,北美洲部分樹木的分布區主要受冬季低溫的控制;Fang和Yoda(1991)指出中國的常綠闊葉林不能分布到平均極端最低溫低于-3 ℃到-2 ℃的地區。另一種說法是,絕大部分物種的祖先是從熱帶地區或是一些濕熱地區進化而來的,它們并不具備抵御寒冷的機制,因此,大部分物種分布受到低溫的控制(Ricklefs,2007)。TEMmin的解釋能力要大于TEM,這可能與研究區空間面較大,氣候空間分異強烈,而TEM只能反映研究單元內總體的氣候狀況,包含的信息量較少,不能全面反映研究單元內氣候的空間分異和季節性分析有關(馮建孟,2008)。
最優模型由TEM、TEMmin、NPP、PRECvar和PRECsd這5項組成,其中PRECvar和PRECsd為季節性因素。多層次方差分解表明,PRECvar的獨立解釋能力和總解釋能力分別為10.0%和34.3%,PRECsd的獨立解釋能力和總解釋能力分別為12.3%和42.0%,略高于TEM的解釋能力。方差分解結果表明,季節性因子的總解釋能力(62.7%)略低于熱量因子(76.6%),但高于水分因子(51.7%)。同樣,其獨立解釋作用(5.6%)也低于熱量因子(6.1%),但高于水分因子(4.5%)。以上都說明季節性因子對于我國兩棲動物分布格局有著重要的影響,也驗證了本文的第3個假設。季節性因子一定程度上代表了氣候的穩定性。氣候穩定性假說認為,穩定的氣候環境能促進物種的特化,并使生態位區域狹窄,因此能容納更多的物種;相反在波動的氣候環境下,物種需要更廣泛的生理機能才能生存下來,因此物種豐富度較低(Stevens,1989)。根據Rapport法則,溫度和降水年較差和波動性由南向北有逐漸增大的趨勢,這也說明了為什么我國南方地區物種豐富度相比北方地區要高(馮建孟,2008)。然而,季節性因子能否作為一個重要的預測因子,目前仍存在廣泛的爭論。對哺乳動物和鳥類的研究發現,季節性因子對其分布有著重要的影響(Andrews & O’Brien,2000;Badgley & Fox,2000;Tello & Stevens,2010),而在昆蟲類群卻得到了相反的結論(Kerr & Packer,1999;Schuldt & Assmann,2009),這可能由于研究的類群不同造成的,如冷血動物和恒溫動物,或者由于它們的取樣粒度不同造成的。
陳勝東, 徐海根, 曹銘昌, 等. 2011. 物種豐富度格局研究進展[J]. 生態與農村環境學報, 27(3): 1-9.
方精云. 2001. 也論我國東部植被帶的劃分[J]. 植物學報, 43(5): 522-533.
費梁, 葉昌媛, 江建平. 2010. 中國兩棲動物彩色圖鑒[M]. 成都: 四川科學技術出版社.
馮建孟, 徐成東. 2009. 中國種子植物物種豐富度的大尺度分布格局及其與地理因子的關系[J]. 生態環境學報, 18(1): 249-254.
馮建孟. 2008. 中國種子植物物種多樣性的大尺度分布格局及其氣候解釋[J]. 生物多樣性, 16(5): 470-476.
王志恒, 唐志堯, 方精云. 2009. 物種多樣性地理格局的能量假說[J]. 生物多樣性, 17(6): 613-624.
Allen AP, Brown JH, Gillooly JF. 2002. Global biodiversity, biochemical kinetics, and the energetic-equivalence rule[J]. Science, 297(5586): 1545-1548.
Andrews P, O’Brien EM. 2000. Climate, vegetation, and predictable gradients in mammal species richness in southern Africa[J]. Journal of Zoology, 251(2): 205-231.
Badgley C, Fox DL. 2000. Ecological biogeography of North American mammals: species density and ecological structure in relation to environmental gradients[J]. Journal of Biogeography, 27(6): 1437-1467.
Borcard D, Legendre P, Drapeau P. 1992. Partialling out the spatial component of ecological variation[J]. Ecology, 73(3): 1045-1055.
Brown JH, Lomolino MV. 2000. Concluding remarks: historical perspective and the future of island biogeography theory[J]. Global Ecology and Biogeography, 9(1): 87-92.
Büchi L, Christin P, Hirzel AH. 2009. The influence of environmental spatial structure on the life-history traits and diversity of species in a metacommunity[J]. Ecological Modelling, 220(21): 2857-2864.
Chen S, Mao L, Zhang J,etal. 2014. Environmental determinants of geographic butterfly richness pattern in eastern China[J]. Biodiversity and Conservation, 23(6): 1453-1467.
Chen SB, Jiang GM, OuYang ZY,etal. 2011. Relative importance of water, energy, and heterogeneity in determining regional pteridophyte and seed plant richness in China[J]. Journal of Systematics and Evolution, 49(2): 95-107.
Chown SL, Gaston KJ. 2010. Body size variation in insects: a macroecological perspective[J]. Biological Reviews, 85(1): 139-169.
Comont RF, Roy HE, Lewis OT,etal. 2012. Using biological traits to explain ladybird distribution patterns[J]. Journal of Biogeography, 39(10): 1772-1781.
Currie DJ, Paquin V. 1987. Large-scale biogeographical patterns of species richness of trees[J]. Nature, 329(6137): 326-327.
Ding TS, Yuan HW, Geng S,etal. 2006. Macro-scale bird species richness patterns of the East Asian mainland and islands: energy, area and isolation[J]. Journal of Biogeography, 33(4): 683-693.
Evans KL, Greenwood JJ, Gaston KJ. 2005. Dissecting the species-energy relationship[J]. Proceedings of the Royal Society of London B: Biological Sciences, 272(1577): 2155-2163.
Fang J, Yoda K. 1991. Climate and vegetation in China V. Effect of climatic factors on the upper limit of distribution of evergreen broadleaf forest[J]. Ecological Research, 6(1): 113-125.
Francis AP, Currie DJ. 2003. A globally consistent richness-climate relationship for angiosperms[J]. The American Naturalist, 161(4): 523-536.
Gaston KJ. 2000. Global patterns in biodiversity[J]. Nature, 405(6783): 220-227.
Gotthardt M, Trommsdorff M, Nevitt MF,etal. 2000. Interactions of the low density lipoprotein receptor gene family with cytosolic adaptor and scaffold proteins suggest diverse biological functions in cellular communication and signal transduction[J]. Journal of Biological Chemistry, 275(33): 25616-25624.
Hawkins BA, Field R, Cornell HV,etal. 2003. Energy, water, and broad-scale geographic patterns of species richness[J]. Ecology, 84(12): 3105-3117.
Hu JH, Li C, Xie F,etal. 2012. Endemic amphibians and their distribution in China[J]. Asian Herpetological Research, 3(2): 163-171.
Kerr JT, Packer L. 1997. Habitat heterogeneity as a determinant of mammal species richness in high-energy regions[J]. Nature, 385(6613): 252-254.
Kerr JT, Packer L. 1999. The environmental basis of North American species richness patterns amongEpicauta(Coleoptera: Meloidae)[J]. Biodiversity & Conservation, 8(5): 617-628.
Klopfer PH. 1959. Environmental determinants of faunal diversity[J]. American Naturalist, 93(837): 337-342.
Latham RE, Ricklefs RE. 1993. Global patterns of tree species richness in moist forests: energy-diversity theory does not account for variation in species richness[J]. Oikos, 67(2): 325-333.
Legendre P, Legendre L. 1998. Numerical ecology 2nd edition[M]. Amsterdam, NL: Elsevier Science.
Mac Nally R. 2000. Regression and model-building in conservation biology, biogeography and ecology: the distinction between- and reconciliation of- ‘predictive’and ‘explanatory’models[J]. Biodiversity & Conservation, 9(5): 655-671.
Orme CDL, Davies RG, Olson VA,etal. 2006. Global patterns of geographic range size in birds[J]. PLoS Biology, 4(7): e208.
Qian H, Ricklefs RE. 2000. Large-scale processes and the Asian bias in species diversity of temperate plants[J]. Nature, 407(6801): 180-182.
Qian H, Wang X, Wang S,etal. 2007. Environmental determinants of amphibian and reptile species richness in China[J]. Ecography, 30(4): 471-482.
Qian H. 2007. Relationships between plant and animal species richness at a regional scale in China[J]. Conservation Biology, 21(4): 937-944.
Rahbek C, Graves GR. 2000. Detection of macro-ecological patterns in south American hummingbirds is affected by spatial scale[J]. Proceedings of the Royal Society of London B: Biological Sciences, 267(1459): 2259-2265.
Ricklefs RE. 2007. History and diversity: explorations at the intersection of ecology and evolution[J]. The American Naturalist, 170(S2): S56-S70.
Rosenzweig ML. 2001. The four questions: what does the introduction of exotic species do to diversity?[J]. Evolutionary Ecology Research, 3(3): 361-367.
Sakai A, Weiser CJ. 1973. Freezing resistance of trees in North America with reference to tree regions[J]. Ecology, 54(1): 118-126.
Sanders NJ, Lessard JP, Fitzpatrick MC,etal. 2007. Temperature, but not productivity or geometry, predicts elevational diversity gradients in ants across spatial grains[J]. Global Ecology and Biogeography, 16(5): 640-649.
Schuldt A, Assmann T. 2009. Environmental and historical effects on richness and endemism patterns of carabid beetles in the western Palaearctic[J]. Ecography, 32(5): 705-714.
Stevens GC. 1989. The latitudinal gradient in geographical range: how so many species coexist in the tropics[J]. American Naturalist, 133(2): 240-256.
Stuart SN, Chanson JS, Cox NA,etal. 2004. Status and trends of amphibian declines and extinctions worldwide[J]. Science, 306(5702): 1783-1786.
Tello JS, Stevens RD. 2010. Multiple environmental determinants of regional species richness and effects of geographic range size[J]. Ecography, 33(4): 796-808.
Tognelli MF, Kelt DA. 2004. Analysis of determinants of mammalian species richness in south America using spatial autoregressive models[J]. Ecography, 27(4): 427-436.
Tognelli MF. 2005. Assessing the utility of indicator groups for the conservation of south American terrestrial mammals[J]. Biological Conservation, 121(3): 409-417.
White P, Kerr JT. 2006. Contrasting spatial and temporal global change impacts on butterfly species richness during the 20th century[J]. Ecography, 29(6): 908-918.
Wright DH. 1983. Species-energy theory: an extension of species-area theory[J]. Oikos, 17(3): 496-506.
Zhao S, Fang J, Peng C,etal. 2006. Patterns of fish species richness in China’s lakes[J]. Global Ecology and Biogeography, 15(4): 386-394.
Relationships between Geographic Amphibian Species Richness Provincial Pattern and Environmental Factors in China
SHEN Mengwei1, BI Mengjie2, JING Qin1, CHEN Wende1, CHEN Shengbin3*
(1. College of Tourism and Urban-Rural Planning, Chengdu University of Technology, Chengdu 610059, China;2. College of Earth Sciences, Chengdu University of Technology, Chengdu 610059, China;3. Nanjing Institute of Environmental Sciences, Ministry of Environmental Protection, Nanjing 210042, China)
Understanding the spatial patterns of species richness is a hot topic in macroecology because of its significance to biodiversity conservation. In this paper, the distribution patterns of amphibian species richness in China and its relationship with environmental factors were investigated based on the amphibian distribution data and environmental variables at provincial scale combined with GIS and statistical methods. The results showed that: (1) species richness decreased significantly with latitude rather than longitude. Amphibian richness was the highest in south and was low in north and northwest China, north Qinghai-Tibet Plateau; (2) The best model was based on the Akaike information criterion including mean annual temperature (TEM), mean temperature of the coldest month (TEMmin), net primary productivity, annual precipitation range and the standard deviation of mean monthly precipitation, moreover, the TEMmin independently explained in amphibian richness (17.6%) was higher than TEM (11.5%) as determined by hierarchical partitioning; (3) The independent effect of seasonality was lower (5.6%) than that of energy (6.1%), but much higher than that of water (4.5%) as reflected by variance partitioning. Therefore, we concluded that seasonality was one of the most important factors limiting amphibian distribution in China.
biogeography; spatial distribution pattern; climate; macroecology
10.11984/j.issn.1000-7083.20150239
2015-07-23 接受日期:2015-10-29
Q14
A
1000-7083(2016)01-0009-08
*通信作者 Corresponding author, E-mail:chainpin@126.com
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
英語世界(2023年10期)2023-11-17 09:18:18
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
光學精密工程(2016年6期)2016-11-07 09:07:19
核科學與工程(2015年4期)2015-09-26 11:59:03
