普通小麥-黑麥草(Loliumperenne L.)易位系鑒定及其后代灌漿與產量性狀分析
2016-12-15 08:23:31王掌軍張文杰萬佳穎白麗娟賈影影
麥類作物學報 2016年7期
關鍵詞:產量
王掌軍,張文杰,萬佳穎,白麗娟,賈影影,
劉 妍1,劉鳳樓1,劉生祥1,張曉崗1
(1.寧夏大學農學院,寧夏銀川 750021; 2.寧夏農林科學院作物研究所,寧夏銀川 750001)
?
普通小麥-黑麥草(LoliumperenneL.)易位系鑒定及其后代灌漿與產量性狀分析
王掌軍1,張文杰2,萬佳穎1,白麗娟1,賈影影1,
劉 妍1,劉鳳樓1,劉生祥1,張曉崗1
(1.寧夏大學農學院,寧夏銀川 750021; 2.寧夏農林科學院作物研究所,寧夏銀川 750001)
為明確外源染色體或片段對小麥籽粒灌漿和產量性狀的影響,以普通小麥寧春4號和黑麥草遠緣雜交后代為材料,借助分子細胞學手段鑒定其BC1代染色體組成,并對其BC1F3代籽粒灌漿和產量性狀進行了分析。結果表明,ND99株系出現兩個頂端小片段易位類型的GISH雜交信號,為真雜種;ND99的BC1F3后代中分離出有芒(ND99A)和無芒(ND99B)兩種類型,并且同一個分子標記3EST-185能夠追蹤這兩種類型。寧春4號和易位系后代(ND99A、ND99B)籽粒的體積、鮮重、干重、水分均表現為先增后降的變化趨勢;寧春4號和ND99A的灌漿速率呈雙峰曲線變化,而ND99B 呈多峰曲線變化。寧春4號的灌漿時間早,快速增長期長,籽粒飽滿指數和粒重最大;ND99A灌漿遲,持續期短,籽粒飽滿指數最低,粒重僅次于寧春4號;ND99B灌漿的峰值出現較早、較大,灌漿持續期短,籽粒飽滿指數居中,粒重最低。易位系后代的穗長、穗粒數和結實小穗數明顯高于寧春4號,可能是影響其灌漿的關鍵因素之一。
普通小麥;黑麥草;易位系;GISH;分子標記;灌漿特性;產量性狀
Ningxia Academy of Agricultural-Forestry Sciences,Yinchuan,Ningxia 750021,China)
由于長期人工選擇和栽培,導致大量小麥優異基因丟失,栽培小麥的遺傳基礎日趨狹窄,遺傳脆弱性逐漸增加[1]。而小麥近緣種蘊含豐富的優異基因資源,如抗病蟲害、抗逆、高蛋白含量等,將其導入普通小麥,可以拓寬遺傳變異范圍[2]。但導入外源遺傳物質后,通常會對某個性狀產生有利影響,而對小麥的其他性狀產生不利影響。因此,在育種實踐中,不僅要注重外源基因是否能夠改良某些性狀,還應重視其對小麥其他性狀的影響,特別是產量性狀,這也是創制的很多攜帶有近緣物種染色體的小麥材料很少能應用于小麥育種實踐中的原因[3-7]。
黑麥草(LoliumperenneL.,2n=2x=14)為禾本科黑麥草屬下的一個種,有高產、營養價值高、抗寒性好、再生速度快等特點[8-9],在寧夏地區種植還表現為不易倒伏、發芽快的特性。已有研究通過遠緣雜交將黑麥草優異基因導入其他物種中,溫常龍等[10]總結了30多年黑麥草與羊茅屬間雜種高產、優質、抗性強的優勢;介紹了通過基因滲入、雄核發育和轉基因手段改良牧草的方法;綜述了黑麥草羊茅屬間雜種基因組原位雜交、分子標記和遺傳圖譜建立、數量性狀位點分析等分子遺傳學的研究進展。王子平等[11]借助花粉管通道法將黑麥草 DNA 導入到水稻保持系中,發現了一份恢復系,并對該恢復系所含恢復基因獨特的遺傳行為進行了分析。作為小麥三級基因庫,20世紀90年代,我國小麥育種家沈天民將黑麥草(母本)和豫麥2號(父本)進行遠緣雜交,培育了高產、優質、多抗的小麥新品種906,此后,再未見以黑麥草為親本通過遠緣雜交培育出小麥新品種或對小麥主要性狀進行遺傳改良方面的報道。鑒于此,本研究以本課題組前期得到的普通小麥寧春4號與黑麥草雜交后代為材料,借助分子細胞遺傳學手段鑒定BC1代外源染色體或片段的來源、身份,并對BC1F3代籽粒灌漿和產量性狀進行分析,以期明確外源染色體或片段對小麥籽粒灌漿和產量性狀的影響,為把近緣種屬的優良基因導入普通小麥、拓寬遺傳變異范圍提供參考信息和種質資源。
1 材料與方法
1.1 材 料
供試材料包括普通小麥寧春4號、黑麥草、寧春4號與黑麥草雜交后回交(寧春4號為輪回親本)BC1代及其F1代回交、自交BC1F3代,均由寧夏大學小麥育種課題組提供。2015年3月7日將寧春4號及其與黑麥草雜交后代BC1F3種植于寧夏大學教學實驗農場,每個材料種植1個小區,每小區20行,行長1.1 m,行寬0.2 m,生育期管理同大田。
1.2 方 法
1.2.1 基因組原位雜交(Genomicinsituhybridization,GISH)分析
根尖細胞染色體制片參照Gill等[12]的方法,并稍作改動。即剪取2個長約1.0~1.5 cm的種子根尖,0 ℃冰水中處理22~24 h,用卡諾氏固定液(95%乙醇∶冰醋酸=3∶1)固定,在45%的醋酸中壓片,相差顯微鏡下鏡檢。
GISH分析參照Jiang和Gill[13]的程序。將45%醋酸壓片-70 ℃冰凍揭片,經70%、95%和100%乙醇梯度脫水后備用。所用探針為經缺口平移法用Fluoresceint-12-dUTP標記的黑麥草基因組DNA。經雜交、洗脫、信號放大及PI復染后,在Olympus BX60型熒光顯微鏡下用450~490 nm激發光波長觀察,用SPOT CCD (SPOT Color Digital Camera)攝取圖像。
1.2.2 分子標記分析
基因組DNA的提取采用SDS法[14]。根據定位于普通小麥中國春7個部分同源群的不同區段的EST序列(http://wheat. pw. usda. gov/ cgi-bin/westsql/map_locus. cgi)或與小麥相應部分同源群具有共線性關系的水稻和短柄草的測序結果設計引物124對,并交由北京奧科生物技術有限責任公司合成。PCR反應體系(10 μL):10× Reaction Buffer 1 μL,MgCl2(25 mmol·L-1)0.8 μL,dNTPs(2.5 mmol·L-1)0.8 μL,上下游引物(10 μmol·L-1)各0.2 μL,模板DNA 25 ng,TaqDNA聚合酶0.5 U,加ddH2O至總體積10 μL。反應條件:94 ℃預變性3 min;94 ℃變性30 s,55 ℃退火45 s,72 ℃延伸1 min,35個循環;72 ℃ 延伸10 min,4 ℃保存。擴增產物采用8%聚丙烯酰胺凝膠電泳檢測,電泳時總電壓為150 V,電泳1.5 h左右,經硝酸銀染色后觀察并照相。
1.2.3 籽粒灌漿和產量性狀分析
2015年在小麥開花盛期準確標記同一時期開花、穗型一致的穗,每個材料標記150穗左右。從開花當天開始,每5 d于上午9 :00前從5個單株上取10個單穗,直至成熟。每次取樣后籽粒人工剝取并放入燒杯,稱鮮重;運用液面差法測量籽粒體積;105 ℃殺青1 h后,75 ℃烘干至恒重測定籽粒干重。籽粒水分=籽粒鮮重-籽粒干重。平均灌漿速率=(前一時間測定的籽粒干重-下一時間測定的籽粒干重)/間隔天數。這兩個指標均用1 000個籽粒計算。
成熟期在1.1 m雙行區內,調查30個有效穗的總小穗數、總粒數和結實小穗數,并稱量每穗粒重,然后計算每穗平均粒數、粒重、不孕小穗數和結實小穗數。不孕小穗數=總小穗數-結實小穗數。籽粒飽滿指數=成熟籽粒曬干體積/最大鮮籽粒體積。經濟系數=經濟產量/生物產量,其中,生物產量為收獲后未脫粒前的植株(中途未取樣的植株)重量,經濟產量為脫粒后的籽粒重量。
1.2.4 數據處理
采用Microsoft Excel 2010處理數據,并作圖。
2 結果與分析
2.1 小麥-黑麥草遠緣雜交后代的GISH分析和分子標記鑒定結果
在普通小麥寧春4號與黑麥草雜交后收獲到的11粒F1代種子中,大多數干癟、發芽力很低,或者發芽不結實,僅有1粒種子長出后代并結實,用寧春4號回交,BC1代經GISH鑒定,所有個體的體細胞染色體數目均為2n=42,但多數沒有檢測到雜交信號,唯有編號為ND99個體的染色體頂端有GISH信號(圖1),說明該個體中小麥與黑麥草發生了易位,且屬于自發易位類型。
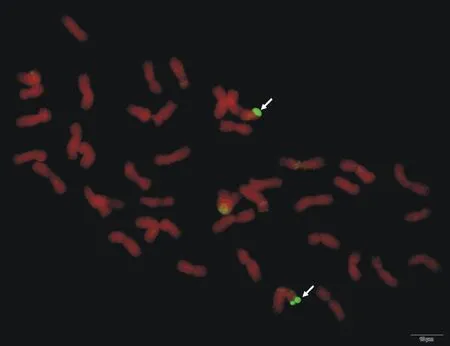
圖中箭頭所指綠色熒光信號為易位的黑麥草染色體;圖示標尺為10 μm。
Chromosome fragments visualized with green fluorescence were fromLoliumperenne; Scale bar was 10 μm.
圖1 ND99根尖細胞染色體的GISH分析
Fig.1 GISH analysis of ND99 root tip cell chromosome
以寧春4號和黑麥草的DNA為模板,利用已開發的124對 EST標記引物進行擴增,篩選到13對在寧春4號和黑麥草間有多態性的標記引物(表1),多態性比率為10.48%。利用這13對具有多態性的引物對寧春4號、黑麥草、ND99及ND99回交、自交后代中分離出的有芒和無芒株系(ND99A和ND99B)進行分析發現,定位于小麥染色體3AL、3BL、3DL上的標記3EST-185在黑麥草、ND99及ND99A和ND99B各15個隨機取樣單株中均擴增出相同的特異性條帶,而寧春4號中并沒有擴增到分子量相同的條帶(圖2)。進一步說明株系ND99為小麥與黑麥草之間發生了易位,而且標記3EST-185可以跟蹤小麥背景中黑麥草染色體片段。
2.2 小麥-黑麥草易位系后代籽粒灌漿特性分析
2.2.1 籽粒形態指標的動態變化
寧夏灌區小麥基本上5月中下旬開花,5月底、6月初開始灌漿,7月中上旬小麥成熟,灌漿結束。本試驗從花后第1天6月1日到7月15日,選取生育期一致的株系,每5d測量1次灌漿期籽粒體積、鮮重、干重和水分等指標。
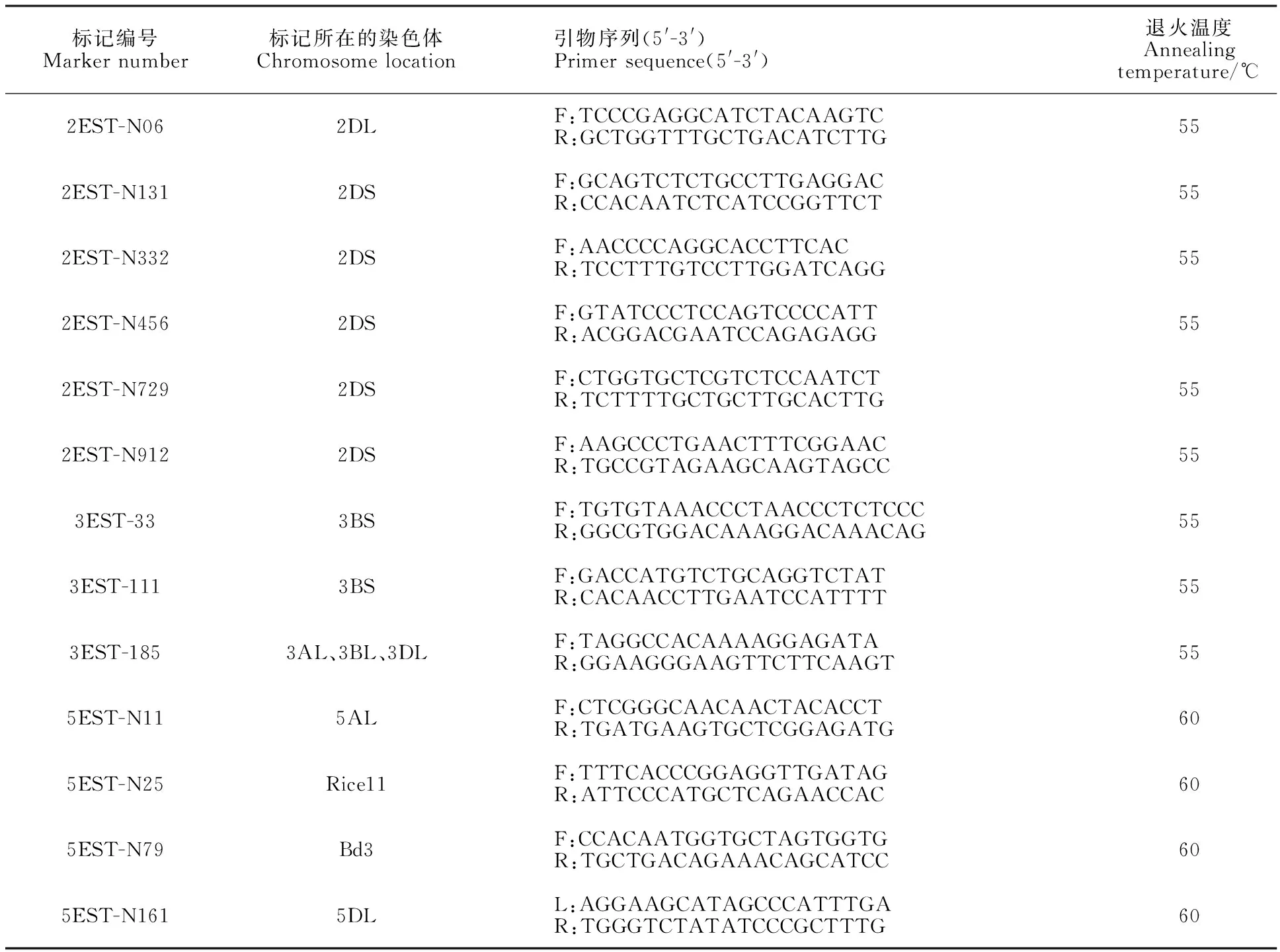
表1 本研究篩選到的黑麥草多態性引物
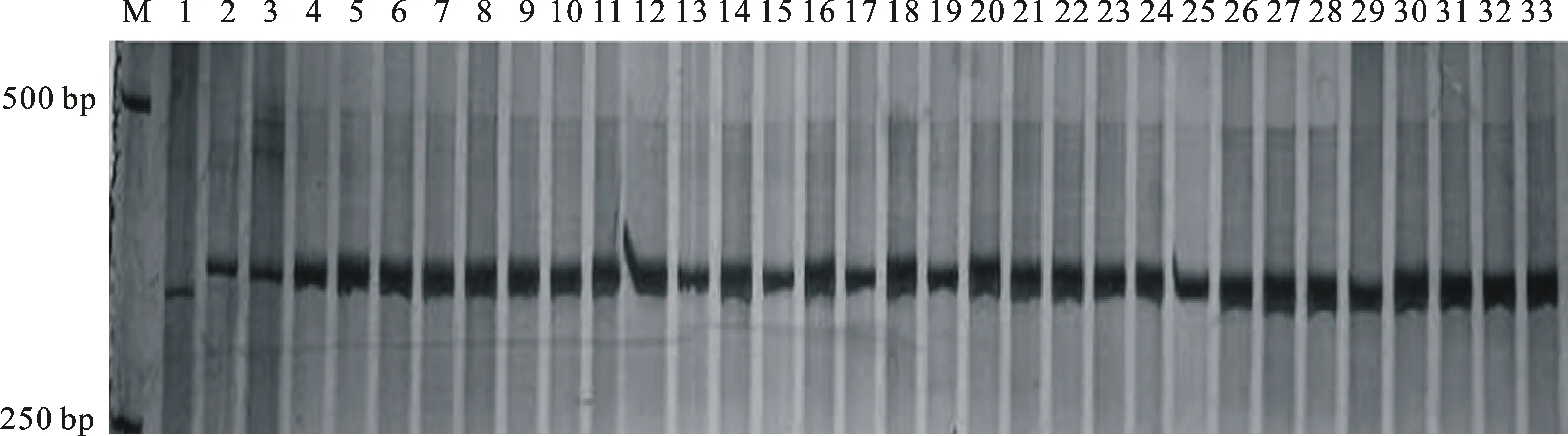
M:DL2000;1:寧春4號;2:黑麥草;3:ND99;4~18:ND99A的15個不同單株;19~33:ND99B的15個不同單株。
M:DL2000; 1:Ningchun 4; 2:Loliumperenne; 3:ND99; 4-18:Fifteen individual plants of ND99A; 19-33:Fifteen individual plants of ND99B.
圖2 標記3EST-185在寧春4號、黑麥草及其二者遠緣雜交后代中的擴增結果
Fig.2 Amplication of marker 3EST-185 in Ningchun 4,LoliumperenneL. and the two distant hybrid offsprings
就籽粒體積而言,寧春4號在花后1~5 d增長較ND99A和ND99B快,花后5~13 d快速增長,此后,增長較為緩慢,花后33 d達到最大(56.391 mL),花后33~41 d迅速降低,花后41~45 d又出現緩慢增長的趨勢;ND99A在花后29~37 d,增長較寧春4號迅速,同在花后33 d達到最大(59.352 mL);ND99B在花后17~21 d和25~29 d增長最為迅速,花后29 d達到最大值(68.105 mL),花后29~45 d體積一直處于變小趨勢(圖3A)。
就籽粒鮮重而言,寧春4號、ND99A和ND99B三個材料在花后1~5 d,表現為穩中有升,之后,各自進入快速增長期;寧春4號和ND99A在花后33 d達到最大值(68.921 g和69.464 g),而ND99B花后29 d達到最大值(78.204 g),隨后,鮮重下降,下降幅度大小順序為:ND99A>ND99B>寧春4號(圖3B)。
就籽粒干重而言,花后1~9 d,寧春4號、ND99A和ND99B均處于平穩增長期,增幅不大,此后迅速增加,寧春4號在花后9~29 d從6.973 g增加到38.062 g,表現為持續快速增加,持續時間最長,花后41 d達到最大值(47.267 g),隨后下降,花后45 d降到43.787 g;ND99A在花后13 d進入持續快速增長期,花后37 d達到最大值(41.303 g);而ND99B分別在花后9~13、17~21、25~29 d有3個間斷的快速增長期,在花后37 d達到最大值(44.822 g),從花后37 d開始,ND99A干重較ND99B大幅度下降(圖3C)。
就籽粒水分而言,花后1~17 d,寧春4號遠高于ND99A和ND99B,花后17 d達到最大值(30.939 g),之后寧春4號的明顯低于其他兩個材料;而ND99A和ND99B的最大值(33.442 g和43.063 g)分別出現在花后25 d和21 d,并且最大值均高于寧春4號;達到最大值后,均以不同幅度下降,寧春4號和ND99A花后41 d下降到最小值(4.715 g和3.747 g),而ND99B花后45 d下降到最小值(5.863 g)(圖3D)。
2.2.2 籽粒灌漿速率的動態變化
由圖4可知,在花后1~5 d,寧春4號千粒重每天增加0.4 g左右,而ND99A和ND99B平均每天增加分別為0.13和0.16 g;花后9~13 d,ND99B灌漿速率最快(1.61 g·d-1),寧春4號次之(1.18 g·d-1),ND99A最低(0.65 g·d-1);花后29~33 d,ND99A首次超過其他兩個材料;花后17~21 d,寧春4號、ND99A和ND99B同時迎來第一個灌漿高峰期,花后25~29 d,3個材料迎來第二個灌漿高峰期,灌漿速率分別為1.75、2.02和2.63 g·d-1,此后,所有材料的灌漿速率呈下降趨勢,花后37~41 d,寧春4號千粒重每天增加0.8g,而ND99A和ND99B的千粒重呈負增長;花后41~45 d,3個材料的千粒重增長均呈負增長。
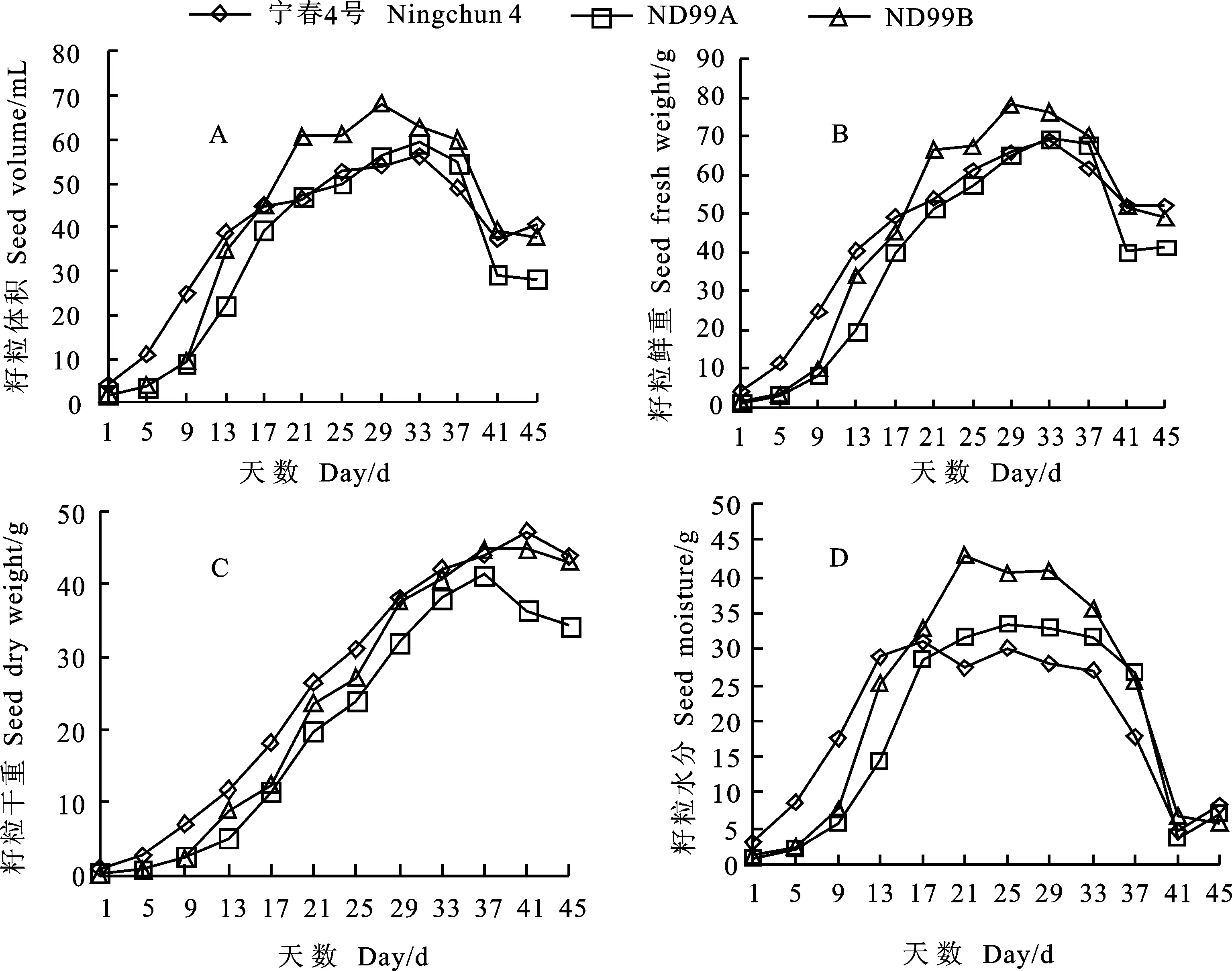
圖3 親本和小麥-黑麥草易位系后代灌漿期籽粒(1 000粒)不同形態指標的動態變化
2.3 小麥-黑麥草易位系后代產量性狀分析
產量性狀分析結果(表2)表明,ND99A和ND99B的穗長分別高出寧春4號32.33%和22.21%,穗粒數明顯高于寧春4號,分別高出86.81%和88.70%,結實小穗數也分別高出42.86%和28.57%,因而易位系后代因為穗長、穗粒數較多和結實小穗數較高,形成較大庫容,進而影響灌漿,導致其穗粒重和籽粒飽滿指數較低;不孕小穗數為ND99A>寧春4號>ND99B;寧春4號的生物產量居中,而經濟產量最高,所以,其經濟系數為最高。總體上,易位系后代有效地提高了穗長、穗粒數、結實小穗數,但是籽粒飽滿指數較低,穗粒重偏低,在生物產量相當的情況下,經濟產量不高,導致其經濟系數偏低。
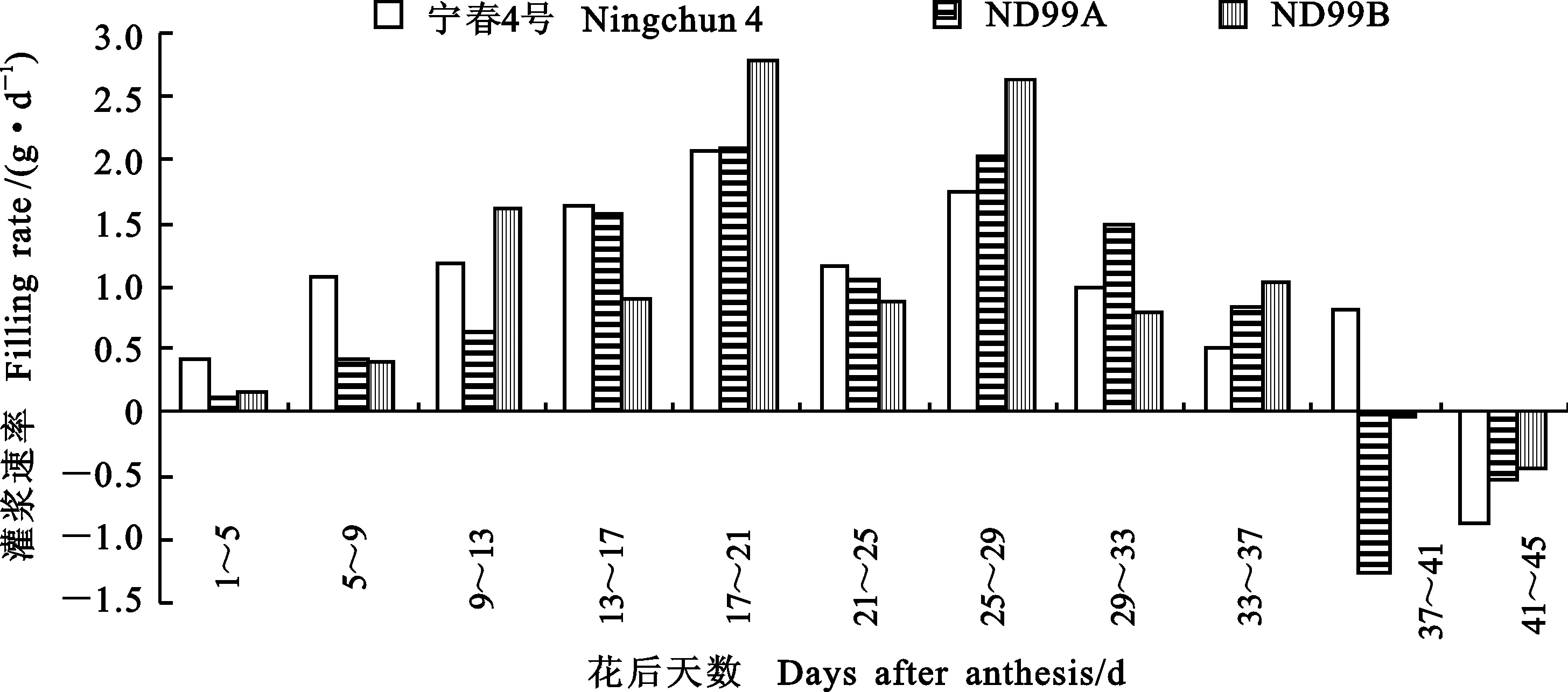
圖4 親本和小麥-黑麥草易位系后代籽粒灌漿速率的動態變化
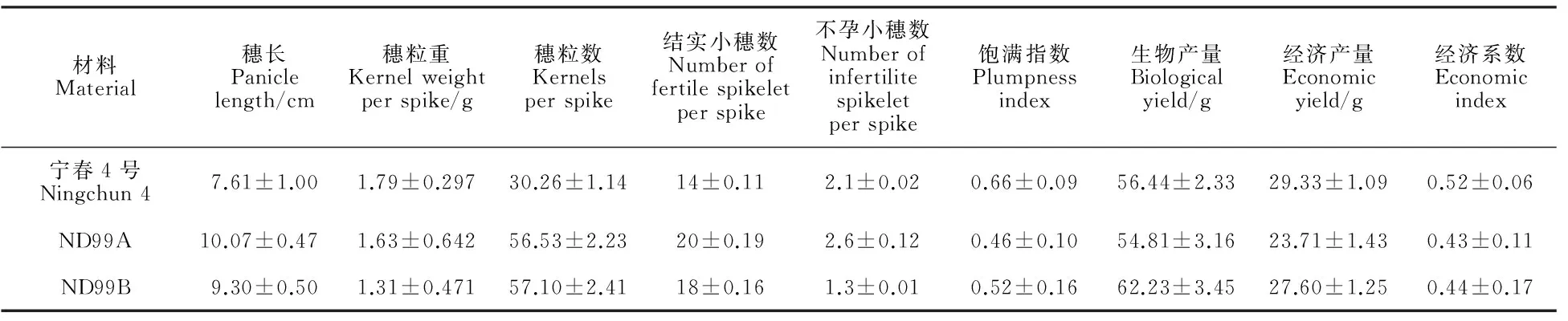
表2 親本和小麥-黑麥草易位系后代產量性狀測定結果
3 討 論
黑麥草作為小麥遺傳改良的三級基因庫中的一個物種[15],因親緣關系較遠,與小麥雜交難度很大[16],而經一些科技工作者的不懈努力,已將外源物種一些優異基因導入小麥背景,在生長上發揮著巨大的作用[17-20];同時,利用GISH[13]和DNA分子標記技術[21-22],有效地鑒定小麥背景中外源染色體的來源、身份。本研究成功利用GISH鑒定了小麥-黑麥草易位系,并篩選到一個黑麥草的特異分子標記,可以看出,因外源染色體(片段)的導入,對小麥灌漿和產量性狀有明顯影響。該研究結果對于黑麥草基因組學研究和挖掘利用這類資源具有現實意義。
籽粒灌漿是產量形成的一個關鍵階段,而小麥灌漿進程受基因型、生態因素(溫度、干旱、地區性等)、生長調節物質、耕作栽培措施等綜合因素的影響[23-28]。本試驗中寧春4號和兩個易位系后代(ND99A和ND99B)灌漿期籽粒的體積、鮮重、干重、水分均表現為先增后降的變化趨勢。易位系后代的籽粒體積、鮮重、水分最大值均高于寧春4號,而最大籽粒干重卻明顯低于寧春4號。同時,寧春4號和ND99A的灌漿速率呈雙峰曲線變化,而ND99B呈多峰曲線變化。其中,(1)籽粒體積增大是灌漿的一個重要標志,但是體積過大,后期籽粒能否飽滿是一個問題。由于易位系后代的最大籽粒體積較大,可能影響到籽粒的飽滿指數,造成籽粒皺縮加深,飽滿程度下降。(2) 由于寧夏灌漿初期氣溫較低,影響到光合強度和物質運輸,同一麥穗不同位置的小穗其發育通常不同步,中部小穗開花比兩端小穗早;中部的籽粒為強勢籽粒,生長迅速,進入灌漿快速期早,而兩端的弱勢籽粒灌漿開始生長緩慢,歷經較長時間才進入灌漿快速期;中部成熟籽粒的粒重高于兩端相應的成熟籽粒。易位系后代的穗長、穗粒數、結實小穗數均高于寧春4號,因為易位系后代屬于強、弱勢粒異步灌漿型[29],所以,粒重較低、不穩定,進而影響其產量。因此,在改良性狀的同時還應加強栽培措施,促其穩產、高產。(3) 籽粒干重是小麥產量最直接的體現,灌漿速率為每天增加的籽粒干重。灌漿速率主要受遺傳控制[30],與粒重顯著正相關[31]。寧春4號籽粒干重比易位系后代持續快速增長時間長,而且干重達到最大值的時間較晚,而易位系后代的干物質積累量提前進入負增長,這主要受到當地溫度等環境因子的調控,綜合采用育種改良和栽培學措施,使其灌漿速度和灌漿時間盡可能協調。(4) 水是植物體完成生命活動所必須的,小麥籽粒不同時期含水量的多少對小麥灌漿影響很大,干物質運輸需要水分,所以,灌漿前期干物質積累與含水量增加呈正相關,這有利于干物質的在小麥體內的積累與運輸;而灌漿后期,易位系后代籽粒水分較高,種子曬干隨著水分的大量蒸發,籽粒飽滿指數較低,造成籽粒皺縮加深,飽滿程度下降;而寧春4號籽粒水分較低,這正是其鮮重不高的情況下籽粒干重較高,飽滿指數和穗粒重較高的原因所在。值得一提的是,易位系在穗長、穗粒數、結實小穗數等性狀方面得到了改良,明顯高于對照寧春4號,這可能是影響其灌漿的關鍵因素之一。今后工作中,對易位系后代的株型(比如矮稈株系選育)、合理的栽培密度、籽粒灌漿同步性等方面進行深入研究,以提高源庫平衡程度,充分發揮其穗長粒多的優勢,通過提高小麥單產保證總產;同時,加強外源基因對小麥品質、抗生物和非生物逆境等方面的潛能,以保證小麥可持續化生產。
[1]JIANG J M,GILL B S.Different species specific chromosome translocations inTriticumtimopheeviiandT.turgidumsupportthe diphyletic origin of polyploid wheats [J].ChromosomeResearch,1994,2(1): 59-64.
[2]SEARS E R.Transfer of Alien Genetic Material to Wheat [M].In:Evans L T,Peacock W J,eds.Wheat Science-Today and Tomorrow.Cambridge:Cambridge University Press,1981:75-89.
[3]陳佩度,周 波,齊莉莉,等.用分子原位雜交(GISH)鑒定小麥-簇毛麥雙倍體、附加系、代換系和易位系 [J].遺傳學報,1995,22(5):380-386.
CHEN P D,ZHOU B,QI L L,etal.Identification of wheat-Haynaldiaviilosaamphiploid,addition,substitution and translocation lines byinsituhybridization using biotin-labelled genomic DNA as a probe [J].ActaGeneticaSinica,1995,22(5):380-386.
[4]KISHII M,YAMADA T,SASAKUMA T,etal.Production of wheat-Leymusracemosuschromosome addition lines [J].TheoreticalandAppliedGenetics,2004,109:255-260.
[5]OLIVER R E,CAI X,XU S S,etal.Wheat alien species derivatives:a novel source of resistance to Fusarium head blight in wheat [J].CropScience,2005,45:1353-1360.
[6]SCHNEIDER A,MOLNR I,MOLNR-LNG M.Utilisation ofAegilops(goatgrass) species to widen the genetic diversity of cultivated wheat [J].Euphytica,2008,163:1-19.
[7]KANG H Y,ZHONG M Y,XIE Q,etal.Production and cytogenetics of trigeneric hybrid involvingTriticum,PsathyrostachysandSecale[J].GeneticResourcesandCropEvolution,2012,59:445-453.
[8]于玉紅,王 亮,黃曉露,等.6個多年生黑麥草品種的交播性狀比較 [J].草業科學,2009,26(8):172-176.
YU Y H,WANG L,HUANG X X,etal.Turf performance of six perennial ryegrass varieties used for overseeding [J].PrataculturalScience,2009,26(8):172-176.
[9]李杰勤,王麗華,詹秋文,等.2個黑麥草品種SPAD值和葉綠素及粗蛋白含量的相關性研究 [J].草業科學,2010,27(10):39-42.
LI J Q,WANG L H,ZHAN Q W,etal.The correlation study of SPAD value,chlorophyll content and crude protein content in two varieties ofLoliumperenne[J].PrataculturalScience,2010,27(10):39-42.
[10]溫常龍,趙 冰,杜建材.黑麥草與羊茅屬間雜種研究進展 [J].中國農業科學,2010,43(7):1346-1354.
WEN C L,ZHAO B,DU J C.Advances in research of festulolium intergeneric hybrids [J].ScientiaAgriculturaSinica,2010,43(7):1346-1354.
[11]王子平,陽標仁.黑麥草恢復基因在水稻中的遺傳行為研究 [J].湖南農業科學,2013(13):7-9.
WANG Z P,YANG B R.Genetic action of ryegrass restoring gene in rice [J].HunanAgriculturalSciences,2011(13):7-9.[12]GILL B S,FRIEBE B,ENDO T R.Standard karyotype and nomenclature system for description of chromosome bands and structural aberration in wheat [J].Genome,1991,34:830-834.
[13]JIANG J M,GILL B S.Sequential chromosome banding andinsituhybridization analysis [J].Genome,1993,36:792-795.
[14]YANG Z J,LI G R,CHANG Z J,etal.Charaeterization of a partial amphiploid betweenTriticumaestivume cv.Chinese Spring andThinopyrumintermidumssp.Trichophorum[J].Euphytica,2006,149:11-17.
[15]DONG F G,SONG J,NAESS S K,etal.Development and applications of a set of chromosome-specific cytogenetic DNA markers in potato [J].TheoreticalandAppliedGenetics,2000,101:1001-1007.
[16]MULLAN D J,MIRZAGHADERI G,WALKER E,etal.Development of wheat-Lophopyrumelongatumrecombinant lines for enhanced sodium 'exclusion' during salinity stress [J].TheoreticalandAppliedGenetics,2009,119:1313-1323.
[17]郭光艷,李瑞芬,張敬原,等.小麥微衛星引物對多枝賴草基因組DNA擴增的研究 [J].華北農學報,2004(1):1-4.
GUO G Y,LI R F,ZHANG J Y,etal.Amplifying products fromLeymusmulticaulisusing wheat microsatellite prime pairs [J].ActaAgriculturaeBoreali-Sinica,2004(1):1-4.
[18]QI L L,PUMPHREY M O,FRIEBE B,etal.A novel Robertsonian translocation event leads to transfer of a stem rust resistance gene( Sr52) effective against race Ug99 fromDasypyrumvillosuminto bread wheat [J].TheoreticalandAppliedGenetics,2011,123:159-167.
[19]KLINDWORTH D L,NIU Z X,CHAO S M,etal.Introgression and characterization of a goatgrass gene for a high level of resistance to Ug99 stem rust in tetraploid wheat [J].GenesGenomesGenetics,2012,2:665-673.
[20]CHEN P D,YOU C F,HU Y,etal.Radition-induced translocations with reducedHaynaldiavillosachromatin at the Pm21 locus for powdery mildew resistance in wheat [J].MolecularBreeding,2013,31:477-484.
[21]ZHANG W,ZHANG R Q,FENG Y G,etal.Distribution of highly repeated DNA sequences inHaynaldiavillosumand its application in the identification of alien chromatin [J].ChineseScienceBulletin,2013,58:890-897.
[22]ZHAO R H,WANG H Y,JIA Q,etal.Development of EST-PCR markers for the chromosome 4V ofHaynaldiavillosaand their application in identification of 4V chromosome structural aberrants [J].JournalofIntegrativeAgriculture,2014,13(2):282-289.
[23]呂厚波,馮安榮,董 瑞,等.不同基因型小麥灌漿和抗倒伏性能與籽粒產量的關系 [J].西北農業學報,2015,24(6):27-33.
Lü H B,FENG A R,DONG R,etal.Grain-filling characteristics and stem lodging resistance and their relationships with grain yields of different genotypic wheat [J].ActaAgriculturaeBoreali-occidentalisSinica,2015,24(6):27-33.
[24]孔治有,唐李軍,張玉榮,等.Wx基因對灌漿期小麥葉片POD活性的影響 [J].湖北農業科學,2015,54(18):4401-4404.
KONG Z Y,TANG L J,ZHANG Y R,etal.Effect ofWxgenes on POD activity of wheat leaves at filling stage [J].HubeiAgriculturalSciences,2015,54(18):4401-4404.
[25]文廷剛,陳昱利,杜小鳳,等.不同植物生長調節劑對小麥籽粒灌漿特性及粒重的影響 [J].麥類作物學報,2014,34(1):84-90.
WEN T G,CHEN Y L,DU X F,etal.Effects of different plant growth regulators on the grain filling characteristics and grain weight in wheat [J].JournalofTriticeaeCrops,2014,34(1):84-90.
[26]張卓亞,王曉琳,許曉明,等.腐植酸對小麥揚花期水分利用效率及灌漿進程的影響 [J].江蘇農業學報,2015,31(4):725-731.
ZHANG Z Y,WANG X L,XU X M,etal.Effect of humic acid on water use efficiency and grouting process of wheat at flowering [J].JiangsuJournalofAgriculturalScience,2015,31(4):725-731.
[27]梁海燕,劉迪迪,張 娜,等.干旱脅迫對不同基因型小麥籽粒灌漿及內源激素的影響 [J].西北農業學報,2015,24(5):41-47.
LIANG H Y,LIU D D,ZHANG N,etal.Effect of water stress on grain filling and endogenous hormone changes in wheat grain of different genotypes [J].ActaAgriculturaeBoreali-occidentalisSinica,2015,24(5):41-47.
[28]張英華,楊佑明,曹 蓮,等.灌漿期高溫對小麥旗葉與非葉器官光合和抗氧化酶活性的影響 [J].作物學報,2015,41(1):136-144.
ZHANG Y H,YANG Y M,CAO L,etal.Effect of high temperature on photosynthetic capability and antioxidant enzyme activity of flag leaf and non-leaf organs in wheat [J].ActaAgronomicaSinica,2015,41(1):136-144.
[29]任紅松,朱家輝,艾比布拉,等.小麥籽粒灌漿特性分析 [J].西北農林科技大學學報(自然科學版),2006,34(3):55-60.
REN H S,ZHU J H,AIBI B L,etal.Analysis on grain filling charaeteristics of wheat variety [J].JournalofNorthwestA&FUniversity(NaturalScienceEdition),2006,34(3):55-60.
[30]WIEGAND C I,CUELLAR J A.Duration of grain filling and kernel weight of wheat as effeeted by temperature [J].CropScience,1981,21(l): 95-101.
[31]馮素偉,胡鐵柱,李 淦,等.不同小麥品種籽粒灌漿特性分析 [J].麥類作物學報,2009,29(4):643-646.
FENG S W,HU T Z,LI G,etal.Analysis on grain filling characteristics of different wheat varieties [J].JournalofTriticeaeCrops,2009,29(4):643-646.
Identification of Translocation Line ofTriticumaestivum-LoliumperenneL. and Analysis on Grain Filling and Yield Traits of Its Offsprings
WANG Zhangjun1,ZHANG Wenjie2,WAN Jiaying1,BAI Lijuan1,JIA Yingying1,LIU Yan1,LIU Fenglou1,LIU Shengxiang1,ZHANG Xiaogang1
(1.Agricultural College of Ningxia University,Yinchuan,Ningxia 750021,China; 2.Institute of Crop Sciences,
In order to explore the effect of exogenous chromosome or fragment on grain filling and yield of wheat,the offsprings of “Ningchun 4 ×Loliumperenne” distant hybridization were used as materials in this study. The chromosome composition of BC1hybrid offsprings was identified based on genomicinsituhybridization(GISH) usingLoliumperenneL. DNA as probe and molecular markers,and characteristics of grain filling and yield traits in BC1F3offsprings were analyzed. The results showed that,No. ND99 had two GISH signals of small fragment translocation types in top,and was true hybrid; The BC1F3offsprings of ND99 were divided into awned type(ND99A) and awnless type(ND99B),and a molecular marker 3EST-185 could identify the two types. The changes of grain volume,fresh weight,dry weight,and water at filling stage in variety Ningchun 4 and translocation offsprings(ND99A,ND99B) showed a trend of first increasing and then decreasing;The filling rate of Ningchun 4 and ND99A showed a double-peak curve,but ND99B showed many peaks.The filling stage of Ningchun 4 was early in time,with long rapid growth period and biggest plumpness index of grain and kernel weight per spike. The filling time of ND99A showed later,with short extending period and lowest plumpness index,and kernel weight per spike was lower than that of Ningchun 4. The filling peak of ND99B appeared earlier and bigger,with short extending period and lowest kernel weight per spike. In the yield traits,panicle length,kernel number and fertile spikelet number per spike of translocation lines were apparently higher than those of Ningchun 4,which could be one of the key factors affecting grain filling.
Triticumaestivum;LoliumperenneL.; Translocation line; GISH; Molecular marker; Filling characteristics; Yield trait
時間:2016-07-07
2016-01-31
2016-04-13
寧夏回族自治區農業育種專項(2013NYYZ0202)
E-mail:wangzj-gs@126.com
S512.1;S330
A
1009-1041(2016)07-0841-08
網絡出版地址:http://www.cnki.net/kcms/detail/61.1359.S.20160707.1529.006.html
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(2021年10期)2021-12-05 16:31:48
今日農業(2021年14期)2021-11-25 23:57:29
今日農業(2021年13期)2021-08-14 01:37:56
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中國果業信息(2019年10期)2019-11-13 01:21:34
中國化肥信息(2019年2期)2019-01-18 15:24:35
中國化肥信息(2019年1期)2019-01-17 21:31:12
中國化肥信息(2019年4期)2019-01-17 18:47:06
