
低溫弱光對不同品種甜辣椒幼苗抗氧化酶活性與質膜透性的影響
2016-12-17 21:21:09孟雅寧嚴立斌范妍芹
山東農業科學 2016年11期
孟雅寧+嚴立斌+范妍芹

摘要:以10個甜(辣)椒品種為參試材料,以茄門(不耐低溫弱光對照,CK1)和湘研1號(耐低溫弱光對照,CK2) 為對照材料,在低溫弱光條件下,研究了幼苗葉片抗氧化酶活性與質膜透性的變化。結果表明:參試材料中8個品種耐低溫弱光性較強,其順序為:AB91-XB>冀研16號、 JF8G-2-1-2-5-1-11-4>冀研28號、 AB91-W222-49176> HPJ-2-2-1-2-1、 BYT-4-1-3-6-8、 AB91-LH-51217;耐低溫弱光較強的品種SOD、POD和CAT活性相對較高,而MDA含量和質膜透性相對較低且變化幅度較小。
關鍵詞:低溫弱光;甜(辣)椒幼苗;抗氧化酶活性;質膜透性
中圖分類號:S641.301 文獻標識號:A 文章編號:1001-4942(2016)11-0038-05
Abstract Using ten sweet (hot) pepper varieties as materials with Qiemen (not resistant to weak temperature and low light, CK1) and Xiangyan 1 (resistant to low temperature and weak light, CK2) as controls, the changes of anti-oxidative enzyme activities and plasma membrane permeability of seedling leaves were studied under low temperature and weak light. The results showed that eight varieties had strong tolerance to low temperature and weak light with the order of AB91-XB>Jiyan 16, JF8G-2-1-2-5-1-11-4>Jiyan 28, AB91-W222-49176>HPJ-2-2-1-2-1, BYT-4-1-3-6-8, AB91-LH-51217. And the varieties with strong tolerance to weak light and low temperature had relatively higher SOD, POD and CAT activities. But their MDA activity and plasma membrane permeability were relatively lower with little changes.
Keywords Low temperature and weak light; Sweet (hot) pepper seedling; Anti-oxidative enzyme activity; Plasma membrane permeability
甜(辣)椒(Capsicum annuum L.) 為喜溫性蔬菜,對溫度、光照反應敏感,是我國栽培面積最大的蔬菜作物之一,亦是保護地蔬菜生產的主要果菜之一。但我國北方保護地栽培期間,正值低溫、弱光環境,使甜(辣)椒正常生長發育受到抑制,給生產造成嚴重損失。
甜(辣)椒低溫弱光條件下的保護酶活性與膜脂過氧化產物含量的變化,可以為保護地栽培品種的選擇提供理論依據。許多研究[1-3]表明,植物的耐低溫弱光性是植物對周圍環境適應性的一種生理反應,與細胞的結構、生理活性及其酶防御系統的活性有著密切關系。
本試驗以10個甜(辣)椒品種為試材,通過測定低溫弱光處理下其葉片相關指標的相對變化,分析低溫弱光脅迫對其抗氧化酶活性與質膜透性的影響,比較不同品種耐低溫弱光的差異,為甜(辣)椒穩產高產及抗逆生理育種提供理論依據。
1 材料與方法
1.1 試驗材料
參試甜(辣)椒材料共10個,分別為HPJ-2-2-1-2-1、BYT-4-1-3-6-8、AB91-XB、冀研28號、AB91-LH-51217、冀研16號、JF8G-2-1-2-5-1-11-4、JF1F2-2-1-7-10、JF8S-1-1-5-4-1-5-2、AB91-W222-49176,依次用M1~M10表示;對照材料2個,分別為茄門(不耐低溫弱光對照,CK1)和湘研1號(耐低溫弱光對照,CK2)。上述材料均為河北省農林科學院經濟作物研究所茄果室選育的不育系、自交系和雜交種。
1.2 試驗方法
將甜(辣)椒種子經浸種、消毒后播于72孔的育苗盤內,育苗基質為常規花卉營養土(由草炭和蛭石混合制成)。育苗環境為保護地自然環境,依照常規生產方法管理,保證幼苗長勢良好。當幼苗長至4葉1心時移入人工氣候箱,培養條件[4]為光強4 000~5 000 lx,光照時間8 h/d;溫度15℃(晝)/5℃(夜),溫度誤差±0.5℃,進行0、7、14、21 d的低溫弱光處理。處理后,選取上數第3-4片功能葉測定指標,重復3次。
1.3 測定項目及方法
1.3.1 SOD活性測定 采用NBT法[5]測定。冰浴研磨提取,粗酶液與反應液充分混勻后,在4 000 lx 熒光燈下顯色20 min后比色,測定560 nm處的吸光度,SOD活性單位以抑制NBT光化還原50%作為一個酶活單位表示。
1.3.2 POD活性測定 采用愈創木酚法[5]測定。冰浴研磨提取,粗酶液與反應液充分混勻后,34℃水浴保溫3 min,470 nm下連續記錄4 min內的吸光度值。
1.3.3 CAT活性的測定 采用紫外吸收法[5]測定。冰浴研磨提取,粗酶液與反應液充分混勻后,加入300 μL H2O2后在240 nm下連續記錄4 min內的吸光度值。
1.3.4 丙二醛(MDA)含量的測定 采用TBA法[6]測定。冰浴研磨提取,加入2 mL 0.6%的TBA,混合后沸水浴30 min,冷卻后3 000 r/min離心10 min,用紫外可見分光光度計(UV-762)在450、532、600 nm處比色測定。
1.3.5 質膜透性的測定 采用電導法[7],以相對電解質滲出率表示,利用DDS-12A型數字電導率儀測定。
2 結果與分析
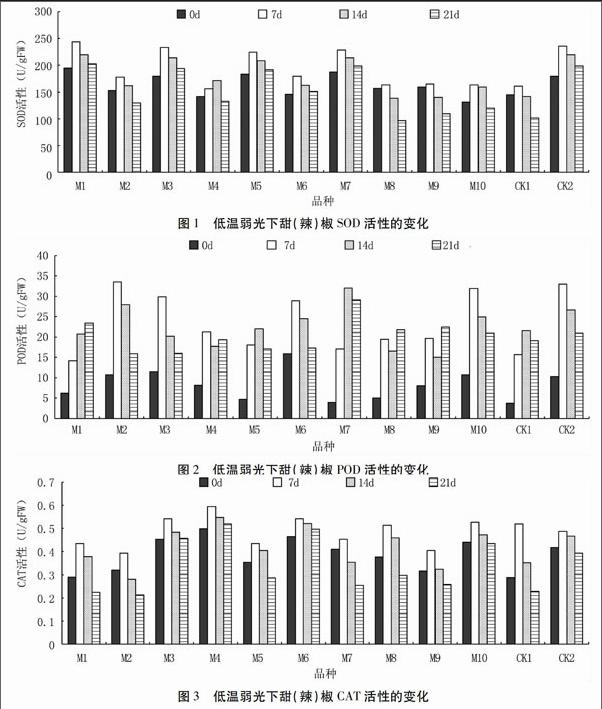
2.1 低溫弱光脅迫對甜(辣)椒葉片SOD活性的影響
由圖1可見,經低溫弱光處理后,甜(辣)椒葉片中的SOD活性均呈單峰型變化趨勢。處理7 d時,所有品種的SOD活性均高于處理前;處理14 d后SOD活性逐漸降低(除M4),其中M8、M9低于處理前;處理21 d時,5個品種(M1、M3、M5、M6、M7)的SOD活性與CK2相似,均下降幅度較小,且均高于處理前,初步認定為耐低溫弱光品種;而其它5個品種與CK1相似,處理21 d后SOD值低于處理前。說明甜(辣)椒受到低溫脅迫時,SOD積極作用以清除活性氧和自由基;但當溫度過低、脅迫時間過長時,SOD活性會受到抑制,從而加速細胞膜的過氧化作用。
2.2 低溫弱光脅迫對甜(辣)椒葉片POD活性的影響
由圖2可見,低溫弱光處理明顯提高了甜(辣)椒葉片POD活性。10個參試品種中,M1的POD活性持續增加,處理21 d時達最高;M2、M3、M5、M6、M7、M10的POD活性呈單峰型變化趨勢;M4、M8、M9的POD活性出現兩個高峰;M2、M3、M6、M7、M10的POD活性與CK2相似,POD活性相對較高且呈單峰變化趨勢,初步認定為耐低溫弱光品種。
2.3 低溫弱光脅迫對甜(辣)椒葉片CAT活性的影響
由圖3可見,低溫弱光處理21 d內,所有參試甜(辣)椒品種的葉片CAT活性均呈單峰型變化趨勢。10個樣品CAT活性均在處理7 d時達到最高值,均高于處理前。處理21 d時,除M3、M4、M6外,其它品種的CAT活性均低于處理前;M3、M4、M6、M10與CK2相似,處理期間CAT活性較高且波動幅度較小,初步認定這4個品種為耐低溫弱光品種;而其它6個品種CAT活性較低且波動幅度較大,與CK1相似。
2.4 低溫弱光脅迫對甜(辣)椒葉片MDA含量的影響
由圖4可見,低溫弱光處理的甜(辣)椒葉片MDA含量均呈單峰型變化趨勢,且均有不同程度的升高。10個參試品種處理7 d時的MDA含量均高于處理前;除M4、M7、M9、M10外,其它品種MDA含量處理14 d較處理7 d時低。處理21 d時,除M3外,其它品種的MDA含量均高于處理前;M2、M5、M8、M9與CK1相似,MDA含量較高且變化幅度較大,而M1、M3、M4、M6、M7、M10與CK2相似,MDA含量相對較低且波動幅度較小,初步認定這6個品種為耐低溫弱光品種。
2.5 低溫弱光脅迫對甜(辣)椒葉片膜透性的影響
由圖5可見,低溫弱光處理的甜(辣)椒膜透性均呈單峰型變化趨勢,且處理后均比處理前有不同程度的升高。在10個參試品種中,M1、M6、M8、M9、M10波動幅度較大,與CK1相似,而M2、M3、M4、M5、M7與CK2相似,膜透性相對較低并且波動幅度較小,初步認定這5個品種為耐低溫弱光品種。
3 討論與結論
研究表明[8-13],當植物受到逆境脅迫時,抗氧化酶的平衡系統就會被打破,引起生理紊亂,嚴重時甚至導致植物死亡。POD、CAT和SOD是膜保護系統重要的3種酶,其中,SOD 是生物體內最重要的活性氧清除酶之一,能有效清除自由基,提高膜的穩定性[14,15];而POD和CAT可以清除體內的H2O2,維持體內的活性氧代謝平衡,保護膜結構,從而使植物能在一定程度上忍耐、減緩或抵抗逆境脅迫[16]。當植株體內活性氧的產生與清除平衡遭到破壞,活性氧大量積累,使得膜脂過氧化加劇,引起膜脂過氧化產物(MDA)增加,造成膜系統損傷[17,18]。
相關分析表明,SOD、POD、CAT活性及MDA含量和細胞膜透性是能較好地反映植物抗逆能力的指標[19,20]。前人研究結果[21]表明,上述指標的變化在種間及不同處理條件下存在差異。本試驗中,甜(辣)椒幼苗在低溫弱光脅迫下啟動了自身的應激反應,引起了一系列的抗氧化酶活性的變化:SOD、CAT活性及MDA含量和膜透性呈單峰型變化趨勢,處理21 d后SOD、CAT活性或高或低于處理前水平,而MDA含量和膜透性幾乎均高于處理前水平;不同品種的POD活性變化規律不同,但處理21 d后均高于處理前。這說明低溫弱光對甜(辣)椒幼苗葉片膜透性及抗氧化酶活性均產生了影響,表明植株為避免遭受低溫弱光逆境的傷害會做出適應性反應。
本研究通過低溫弱光脅迫下幼苗葉片5個生理特性指標的變化,對參試甜(辣)椒品種的耐低溫弱光性進行了評價,根據SOD活性鑒定出M1、M3、M5、M6、M7為耐低溫弱光品種;根據POD活性鑒定出M2、M3、M6、M7 、M10為耐低溫弱光品種;根據CAT活性鑒定出M3、M4、M6、M10為耐低溫弱光品種;根據MDA含量鑒定出M1、M3、M4、M6、M7、M10為耐低溫弱光品種;根據葉片膜透性鑒定出M2、M3、M4、M5、M7為耐低溫弱光品種。根據這5個生理指標鑒定耐低溫弱光品種的次數排序,耐低溫弱光性的順序為:M3>M6、M7>M4、M10>M1、M2、M5,初步認定這8個品種為耐低溫弱光材料。但植物生理指標眾多,本研究僅測定了其中的幾個指標,其它指標隨逆境的變化如何還有待進一步研究。此外,本試驗結果僅是在苗期人工模擬自然環境條件下得出的,所設溫度和光照與自然環境存在差異,至于各品種在大田日光溫室、保護地栽培中的具體表現如何,尚有待進一步研究與驗證。
參 考 文 獻:
[1] 張渝潔. 低溫弱光對不同品種青菜生理生化指標的影響[J].江蘇農業科學,2009,5(2):179-180.
[2] 郁繼華,張國斌,馮致,等. 低溫弱光對辣椒幼苗抗氧化酶活性與質膜透性的影響[J].西北植物學報,2005,25(12):2478-2483.
[3] 劉皓.干旱脅迫下杠柳種子萌發期保護酶活性及丙二醛含量變化[J].農產品加工,2014(8):53-56.
[4] 毛愛軍,耿三省.低溫對甜椒生長發育的影響及甜椒耐低溫篩選方法的研究[J].中國辣椒,2001(1):17-21.
[5] 徐偉慧.辣椒耐低溫性鑒定方法的研究[D].蘭州:甘肅農業大學,2006.
[6] 李合生.植物生理生化實驗原理和技術[M].北京:高等教育出版社,2000.
[7] 西北農業大學植物生理生化教研室.植物生理學實驗指導[M].西安:陜西科學技術出版社,1987:149-151.
[8] 李愛民,張永泰,熊飛,等.低溫弱光對西瓜幼苗生理特性的影響[J].揚州大學學報(農業與生命科學版),2012, 33(2):78-82.
[9] 周兵,閆小紅,蔣平,等.青葙根氯仿相提取物對多種植物的生物活性及抑菌作用[J].華中農業大學學報,2010,29(2):143-147.
[10]侯立剛,陳溫福,馬巍,等. 低溫脅迫下不同磷營養對水稻葉片質膜透性及抗氧化酶活性的影響[J].華北農學報,2012,27(1):118-123.
[11]鄧化冰,王天順,肖應輝,等.低溫對開花期水稻穎花保護酶活性和過氧化物積累的影響[J].華北農學報,2010,25(增刊):62-67.
[12]柴文臣,馬蓉麗,邢國明,等.辣椒耐低溫研究進展[J].山西農業科學,2009,37(1):85-87.
[13]王玉玲,康潔.低溫脅迫對冬小麥苗期和拔節期生理生化特性的影響[J].河南農業科學,2004(5):3-6.
[14]張圣平,顧興芳,王燁,等.低溫脅迫對以野生黃瓜(棘瓜)為砧木的黃瓜嫁接苗生理生化指標的影響[J].西北植物學報,2005,25(7):1428-1432.
[15]王文,陸振德,羅慶熙,等.外源NO對苯丙烯酸脅迫下黃瓜幼苗生長及活性氧代謝的影響[J]. 中國農業科學,2010,43(17):3677-3683.
[16]Zhou S J,Zhu Y Q,Chen L P,et al. Studies on growth and physiologic characteristics of different cucumber cultivars under low temperature and light intense[J].Journal of Yangzhou University:Agricultural and Life Science Edition,2007,28(2):91-94.
[17]Reddy A R,Chaitanya K V,Vivekanandan M. Drought induced responses of photosynthesis and antioxidant metabolism in higher plants [J]. Journal of Plant Physiology,2004,161(11):1189-1202.
[18]葛體達,隋方功,白莉萍,等.水分脅迫下夏玉米根葉保護酶活性變化及其對膜脂過氧化作用的影響[J].中國農業科學,2005,38(5):922-928.
[19]Zhang E P, Zhang S H, Si L T. Influence of NaCl stress on membrane-lipid peroxidation of cotyledon in cucumber seedlings[J]. Journal of Shenyang Agricultural University,2001,32(6):446-448.
[20]張治平,於丙軍,汪良駒,等.低溫下ALA對番茄光合色素和抗氧化酶活性的影響[J]. 江蘇農業學報,2014,30(1):222-224.
[21]于龍鳳,安福全,李富恒.低溫弱光對西葫蘆氣孔特性及膜脂過氧化的影響[J].天津農業科學, 2013,19(2):9-11.
