基于COⅠ基因分析遼東灣海蜇群體遺傳多樣性
2016-12-19 07:19:51李玉龍王文波
水產(chǎn)科學(xué) 2016年4期
關(guān)鍵詞:研究
李玉龍,王 彬,王文波,董 婧
( 遼寧省海洋水產(chǎn)科學(xué)研究院,遼寧省海洋生物資源與生態(tài)學(xué)重點(diǎn)實(shí)驗(yàn)室,遼寧 大連 116023 )
基于COⅠ基因分析遼東灣海蜇群體遺傳多樣性
李玉龍,王 彬,王文波,董 婧
( 遼寧省海洋水產(chǎn)科學(xué)研究院,遼寧省海洋生物資源與生態(tài)學(xué)重點(diǎn)實(shí)驗(yàn)室,遼寧 大連 116023 )
為研究海蜇群體的遺傳多樣性狀況,采用PCR擴(kuò)增獲得20個(gè)海蜇野生群體樣品的線粒體COⅠ基因部分序列,并結(jié)合GenBank上其他15個(gè)海蜇樣品的同源序列,對(duì)其序列變異和遺傳分化進(jìn)行分析。結(jié)果顯示,35條序列的A、T、C、G堿基含量分別為26.7%、36.4%、18.8%、18.1%;在長(zhǎng)度624 bp的線粒體COⅠ基因片段中共發(fā)現(xiàn)21個(gè)多態(tài)位點(diǎn),定義了17種單倍型,單倍型多態(tài)性為0.91±0.03,核苷酸多態(tài)性為0.0056±0.0032。同其他幾種大型水母相比,海蜇種群的遺傳多樣性處于較高或中等水平。研究結(jié)果表明,不同海蜇種群尤其是相距較遠(yuǎn)的群體在其分布范圍內(nèi)可能存在遺傳分化現(xiàn)象。
海蜇;線粒體DNA;COⅠ基因;遺傳多樣性
海蜇(Rhopilemaesculentum)是經(jīng)濟(jì)價(jià)值最高的一種大型食用水母且兼有藥用功效,屬于海產(chǎn)八珍之一,是我國重要的海洋漁業(yè)資源。海蜇在分類學(xué)上隸屬刺胞動(dòng)物門、缽水母綱、根口水母目、根口水母科、海蜇屬。該物種為暖水性河口種類,適應(yīng)的水溫和鹽度范圍較廣,曾廣泛分布于我國北至鴨綠江口,南至北部灣近岸淺海水域,且在日本西部、朝鮮半島南部和俄羅斯遠(yuǎn)東地區(qū)也有分布[1-4]。在我國從南到北沿海一帶,曾有多個(gè)地理種群,如粵東、閩南、閩東、浙南、杭州灣、海州灣、萊州灣、渤海灣和遼東灣等群體。近十年來,由于人為及自然原因,加上災(zāi)害水母頻發(fā),海蜇自然資源量呈顯著下降趨勢(shì),海蜇的多個(gè)地理種群可能已不復(fù)存在,如黃海北部的海蜇群體自1986年后已經(jīng)逐漸消失,渤海灣、萊州灣、浙江近海、福建近海等海域曾經(jīng)豐富的海蜇資源也已經(jīng)逐漸衰退[1,5]。
目前,對(duì)海蜇的研究主要集中在形態(tài)學(xué)分類、生活史、生態(tài)及增殖放流等[1,5-14]方面,對(duì)海蜇的生態(tài)習(xí)性、種群特征,資源管理、苗種培育、池塘養(yǎng)殖、沿岸增殖等領(lǐng)域進(jìn)行了較為系統(tǒng)全面的研究。但迄今為止,有關(guān)海蜇自然群體的遺傳變異及群體間遺傳分化等方面的研究尚不多,所用的分子標(biāo)記主要是核基因分子標(biāo)記如擴(kuò)增片段長(zhǎng)度多態(tài)性和內(nèi)轉(zhuǎn)錄間隔區(qū)序列。高祥剛等[15]采用擴(kuò)增片段長(zhǎng)度多態(tài)性技術(shù)研究了遼寧沿海海蜇和沙蜇(Nemopilemanomurai)野生群體和養(yǎng)殖群體的遺傳多樣性,發(fā)現(xiàn)遼寧沿海的海蜇和沙蜇遺傳多樣性仍處于較高水平。喬洪金等[16]同樣采用擴(kuò)增片段長(zhǎng)度多態(tài)性技術(shù)對(duì)蓬萊、江蘇2個(gè)海蜇捕獲群體以及江蘇2個(gè)海蜇養(yǎng)殖群體共56個(gè)個(gè)體的遺傳多樣性進(jìn)行了分析,結(jié)果同樣表明海蜇遺傳多樣性水平較高,但并未發(fā)現(xiàn)海蜇不同群體間存在遺傳分化現(xiàn)象。孫國華等[17]利用內(nèi)轉(zhuǎn)錄間隔區(qū)序列對(duì)煙臺(tái)萊州灣、江蘇海州灣海蜇捕獲群體和威海養(yǎng)殖群體共24個(gè)個(gè)體進(jìn)行的序列分析亦表明海蜇群體間無明顯的遺傳分化,但遺傳多樣性水平較低。雖然上述研究為海蜇群體的遺傳變異提供了一定的參考,但所用的分子標(biāo)記或者樣品數(shù)量均相對(duì)較少,而要全面了解這一物種的遺傳多樣性背景,還需利用更多的遺傳標(biāo)記方法對(duì)更多的海蜇樣品進(jìn)行研究。
線粒體DNA因具有結(jié)構(gòu)簡(jiǎn)單、無重組、進(jìn)化速率較快且多數(shù)母性遺傳等特點(diǎn),已成為進(jìn)行群體遺傳多樣性分析的理想研究手段,被廣泛應(yīng)用于海洋動(dòng)物群體遺傳學(xué)研究中。線粒體CO Ⅰ基因和核糖體內(nèi)轉(zhuǎn)錄間隔區(qū)基因是研究水母類遺傳多樣性及其遺傳結(jié)構(gòu)常用的分子標(biāo)記,利用兩種分子標(biāo)記對(duì)幾種缽水母類如肺狀根口水母(Rhizostomapulmo)和夜光游水母(Pelagianoctiluca)的研究結(jié)果表明,線粒體CO Ⅰ基因片段比內(nèi)轉(zhuǎn)錄間隔區(qū)序列具有更多的變異位點(diǎn),更適合作為檢測(cè)缽水母類群體遺傳多樣性及遺傳分化的分子標(biāo)記[18-19]。目前,尚未見利用線粒體DNA作為分子標(biāo)記對(duì)海蜇進(jìn)行遺傳多樣性分析的相關(guān)報(bào)道。本研究以遼東灣野生海蜇群體為研究對(duì)象,對(duì)其CO Ⅰ基因片段進(jìn)行比較分析,并與GenBank數(shù)據(jù)庫中不同海域的海蜇同源序列進(jìn)行比較,了解不同海域海蜇群體的遺傳背景及遺傳分化等情況,以期為海蜇遺傳育種及種質(zhì)資源保護(hù)提供科學(xué)依據(jù)。
1 材料與方法
1.1 樣品采集
因野生海蜇樣品數(shù)量較少難以采集,本研究?jī)H對(duì)20個(gè)海蜇樣品進(jìn)行分析,其中19個(gè)取自2012年6—8月進(jìn)行的遼東灣海蜇跟蹤監(jiān)測(cè)調(diào)查中所獲得的樣品(取樣站位見圖1黑色圓點(diǎn)所示),每個(gè)站位隨機(jī)取樣(1~5個(gè)樣品不等),傘徑40~440 mm,剩余1個(gè)樣品為2013年取自韓國江華島附近海域。樣品采集后取傘部及肩板組織于95%酒精中保存?zhèn)溆谩?/p>
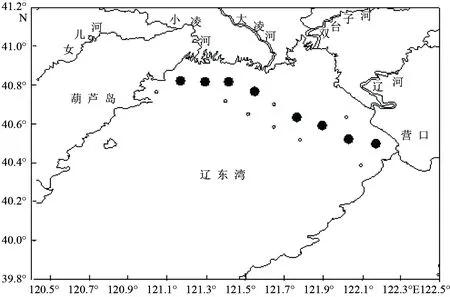
圖1 遼東灣海蜇樣品取樣站位
1.2 DNA提取、擴(kuò)增及測(cè)序
采用CTAB法提取基因組DNA,PCR擴(kuò)增在Eppendorf AG型PCR儀上進(jìn)行,擴(kuò)增引物序列均為:FFDL 5′-TTTCAACTAACCAYAAAGAYATWGG-3′ 和FRDL2 5′-TANACTTCWGGRTGNCCRAAGAATCA-3′[20]。25 μL反應(yīng)體積,包括:0.2 mmol/L每種dNTPs,0.2 μmol/L每種引物,1 μLDNA模板,1 UTaq,2.0 mmol/L MgCl2,2.5 μL 10×緩沖液,滅菌超純水補(bǔ)足剩余體系。反應(yīng)程序:95 ℃預(yù)變性3 min后,95 ℃變性30 s,51 ℃退火30 s,72 ℃延伸45 s,運(yùn)行35個(gè)循環(huán),最后72 ℃延伸5 min。擴(kuò)增產(chǎn)物純化后進(jìn)行雙向測(cè)序(上海英濰捷基有限公司)。
1.3 數(shù)據(jù)分析
將測(cè)定的COⅠ序列進(jìn)行拼接和人工校對(duì),并通過將COⅠ基因序列翻譯成氨基酸序列來證實(shí)其準(zhǔn)確性和排除假基因干擾。從GenBank數(shù)據(jù)庫中下載了15條海蜇樣品的同源序列與本研究所獲得的序列進(jìn)行分析,根據(jù)GenBank數(shù)據(jù)庫中信息按樣品的地理來源將海蜇劃分為3個(gè)群體,采自日本海的3個(gè)樣品HF930514-15、KF840718歸于日本海群體(JP),遼東灣19個(gè)海蜇樣品及2個(gè)采自渤海的樣品EU373722-23屬于渤海群體(BH),采自韓國的1個(gè)樣品與采自黃海的6個(gè)樣品EU3737224、JX845347、HF536571-74歸為黃海群體(HH),另有4條序列HF930546、HF930550、HF937282、HF937287為采自于中國的海蜇樣品,未標(biāo)明采集地,最后分子變異分析時(shí)將其歸為中國組群進(jìn)行計(jì)算(表1)。利用DnaSP v5[21]軟件確定單倍型。單倍型數(shù)目、多態(tài)位點(diǎn)、單倍型多態(tài)度、核苷酸多態(tài)度等分子多樣性指數(shù)由Arlequin 3.01[22]軟件計(jì)算獲得。通過MEGA 3.0[23]計(jì)算堿基含量及單倍型間的遺傳距離。使用Arlequin 3.01軟件中的分子變異分析[24]來評(píng)估群體間遺傳變異,其顯著性通過1000次重復(fù)抽樣來檢驗(yàn),群體間的遺傳距離采用Kimura2-parameter模型計(jì)算。
利用Mega 3.0軟件[23]構(gòu)建海蜇群體COⅠ單倍型的鄰接系統(tǒng)樹,系統(tǒng)樹的可靠性采用1000次重復(fù)抽樣進(jìn)行評(píng)估。此外,為探討海蜇單倍型的譜系結(jié)構(gòu),采用中介網(wǎng)絡(luò)法[25]構(gòu)建單倍型網(wǎng)絡(luò)關(guān)系圖。
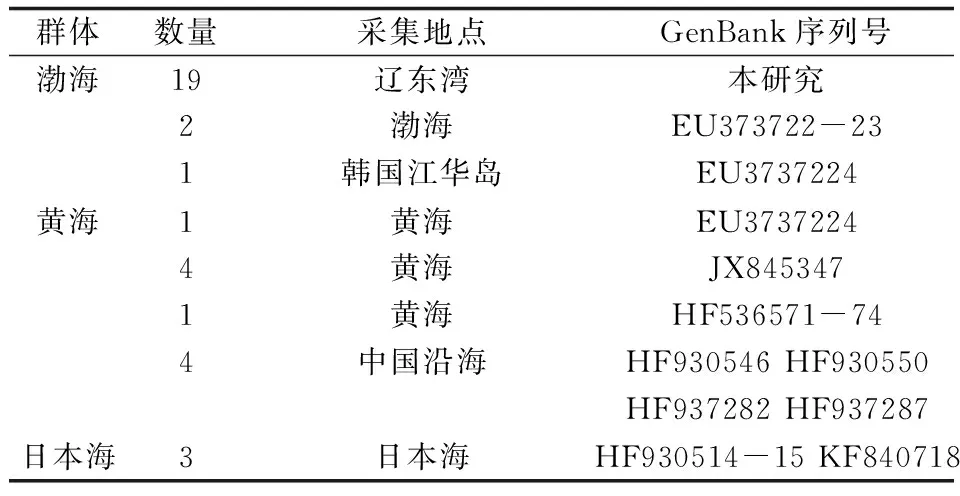
表1 試驗(yàn)用海蜇樣品的采集數(shù)量、地點(diǎn)及引用序列登錄號(hào)
2 結(jié)果與分析
2.1 堿基組成及序列變異
經(jīng)比對(duì)去掉兩端引物本研究分析的海蜇CO Ⅰ基因同源序列長(zhǎng)度為624 bp,其A、T、G、C 4種堿基的平均含量分別為26.7%、36.4%、18.8%、18.1%,A+T(63.1%)含量明顯高于G+C(36.9%)含量。624個(gè)位點(diǎn)中共檢測(cè)到21個(gè)多態(tài)位點(diǎn),包括9個(gè)簡(jiǎn)約信息位點(diǎn)和12個(gè)單態(tài)核苷酸變異位點(diǎn)。大部分突變(86%)發(fā)生在密碼子第3位上,有3個(gè)突變發(fā)生在密碼子第1位,所有突變位點(diǎn)均未導(dǎo)致氨基酸的改變。21個(gè)變異位點(diǎn)共定義了17種單倍型,定義為單倍型1~17,單倍型間遺傳距離為0.2%~1.5%,平均遺傳距離0.7%。單倍型6、9、10、11、14是海蜇群體的共有單倍型,其中單倍型6、9、10為主體單倍型,其所占頻率分別為17%、23%、14%。除此之外,其他單倍型均為各群體特有單倍型。單倍型頻率及其在3個(gè)群體中的分布見表2。
計(jì)算了海蜇不同群體及總?cè)后w的單倍型多樣性指數(shù)和核苷酸多樣性指數(shù),其中因日本海海蜇樣品數(shù)量太少(n=3),未計(jì)算該群體的多樣性指數(shù),同時(shí)根據(jù)已有資料比較了海蜇與其他幾種大型水母的相關(guān)遺傳多樣性參數(shù)(表3),結(jié)果表明,海蜇群體的COⅠ變異水平較高,除夜光游水母外,與沙蜇或肺狀根口水母相比,海蜇的遺傳多樣性水平最高。
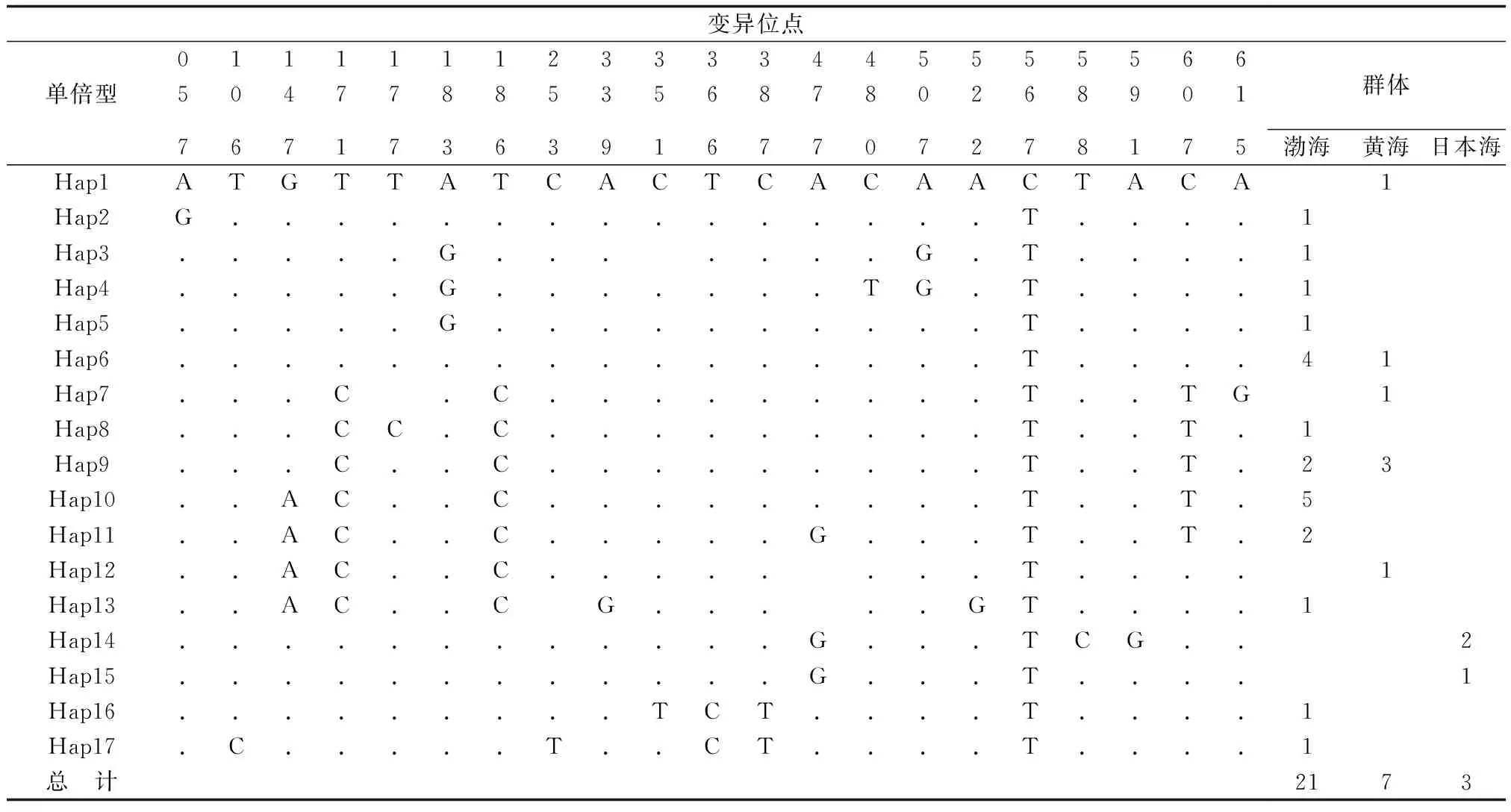
表2 海蜇變異位點(diǎn)及單倍型在各群體中的分布
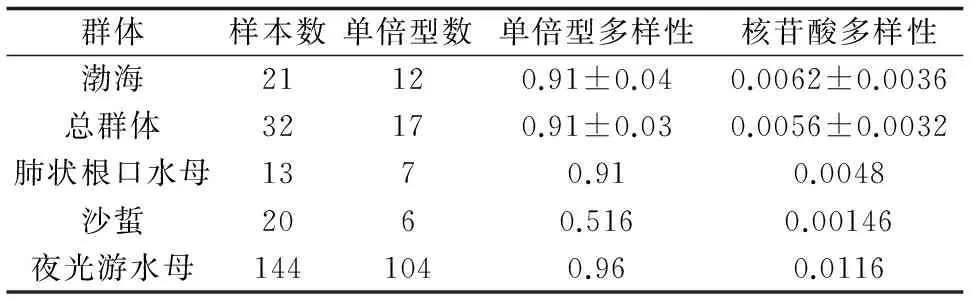
表3 海蜇與其他3種大型水母COⅠ基因遺傳多樣性參數(shù)比較
2.2 群體間遺傳分化
基于COⅠ基因?qū)r?個(gè)群體的分子變異分析表明,80.71%的差異屬于群體內(nèi)差異,群體間差異為19.29%,遺傳分化指數(shù)為0.1929且差異顯著(P=0.005),說明海蜇群體間有較明顯的遺傳分化(表4)。進(jìn)一步將海蜇分為兩組,中國沿海海蜇樣品為一組,日本海海蜇樣品為另一組,計(jì)算結(jié)果表明遺傳分化指數(shù)為0.418且差異極顯著(P=0)。
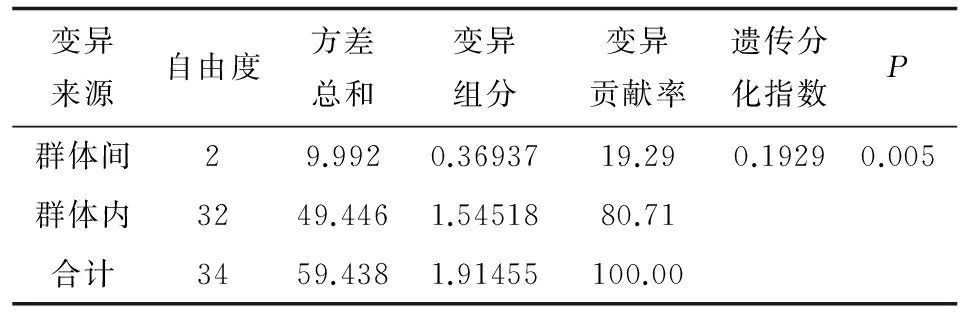
表4 海蜇群體的AMOVA分析
2.3 分子系統(tǒng)分析
利用Mega 3.0軟件構(gòu)建了海蜇的單倍型鄰接關(guān)系樹(圖2)。由圖2可見,鄰接關(guān)系樹分成2個(gè)明顯的進(jìn)化枝,其中來自日本海的2個(gè)單倍型首先聚為一支,再與來自渤海的7個(gè)單倍型以及韓國的1個(gè)單倍型聚為類型Ⅰ;而來自黃海的3個(gè)單倍型和渤海剩余的單倍型則聚為類型Ⅱ。
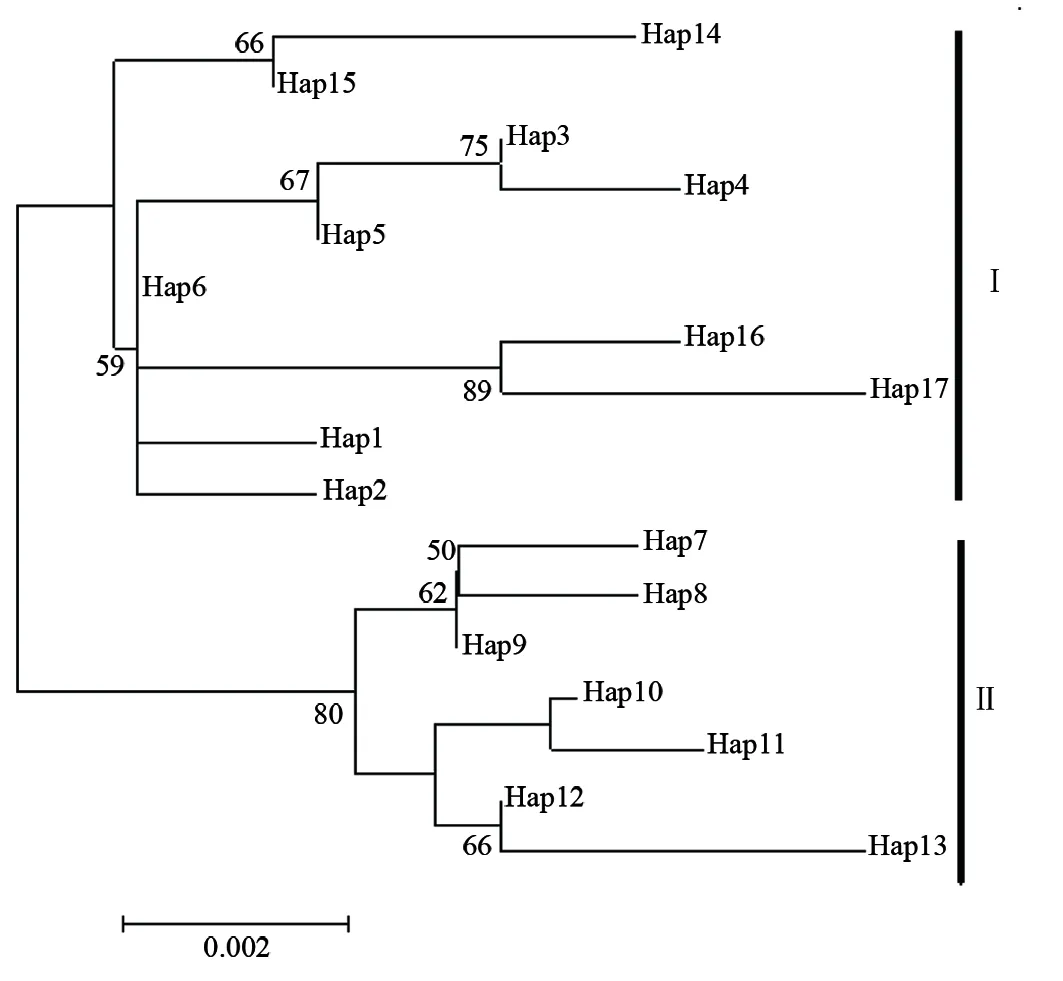
圖2 海蜇單倍型鄰接關(guān)系樹
采用中介網(wǎng)絡(luò)法構(gòu)建了海蜇單倍型的網(wǎng)絡(luò)關(guān)系圖(圖3)。與單倍型鄰接關(guān)系樹的拓?fù)浣Y(jié)構(gòu)一致,單倍型網(wǎng)絡(luò)關(guān)系圖顯示出2個(gè)明顯的單倍型譜系分支,主體單倍型6和9、10分別位于2個(gè)分支Ⅰ和Ⅱ的中心,其他單倍型通過一步或多步突變分別與主體單倍型相連。
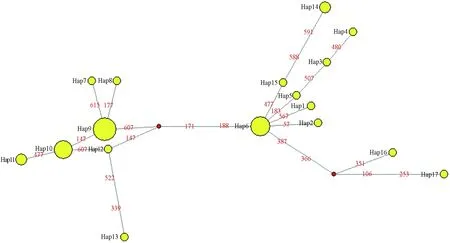
圖3 海蜇單倍型的中介網(wǎng)絡(luò)圓圈面積表示單倍型的頻率,數(shù)字表示突變位點(diǎn)
3 討 論
3.1 海蜇群體COⅠ基因遺傳多樣性分析
海蜇曾是我國近海重要的漁業(yè)資源,全國的海蜇漁業(yè)最高產(chǎn)量曾高達(dá)581 930 t[5],但由于過度捕撈及海洋生態(tài)環(huán)境破壞等不利因素影響,近年來海蜇資源量呈顯著下降趨勢(shì)。近幾年,遼寧、山東、江蘇等省份均在其傳統(tǒng)的海蜇漁場(chǎng)開展了海蜇生產(chǎn)性放流工作,取得一些效果,增加了海蜇產(chǎn)量。但是,有研究表明,人工增殖放流在增加資源量的同時(shí)會(huì)污染原物種自然群體的遺傳基因,破壞遺傳結(jié)構(gòu),造成種群遺傳多樣性降低。本文基于線粒體CO Ⅰ基因部分片段的研究結(jié)果顯示,與其他幾種大型水母相比較,海蜇群體的遺傳多樣性處于中等或較高水平,這與用擴(kuò)增片段長(zhǎng)度多態(tài)性方法[15-16]對(duì)海蜇群體進(jìn)行研究得出的研究結(jié)果相一致,而與孫國華等[17]利用內(nèi)轉(zhuǎn)錄間隔區(qū)序列對(duì)煙臺(tái)萊州灣、江蘇海州灣群體和威海養(yǎng)殖群體得出的結(jié)果不同。基于內(nèi)轉(zhuǎn)錄間隔區(qū)序列對(duì)海蜇群體進(jìn)行的分析表明海蜇群體的遺傳多樣性水平較低,這一方面可能是由于內(nèi)轉(zhuǎn)錄間隔區(qū)序列變異較少所致,另一方面其試驗(yàn)樣品取自放流海蜇的捕撈群體,也會(huì)導(dǎo)致所研究群體的遺傳多樣性水平較低。這種現(xiàn)象在本研究中也有體現(xiàn),海蜇渤海群體的遺傳多樣性參數(shù)明顯高于黃海群體,其中渤海的海蜇樣品大多為采自遼東灣的野生海蜇群體,而黃海的海蜇樣品大多為捕撈的放流海蜇樣品,因此遺傳多樣性水平較低。
3.2 海蜇群體的遺傳分化
分子變異分析的結(jié)果表明,在本研究中渤海海蜇群體和GenBank數(shù)據(jù)庫中樣品來自黃海的海蜇群體和日本海的海蜇群體間存在群體遺傳結(jié)構(gòu),其遺傳分化指數(shù)為0.1929且差異檢驗(yàn)的結(jié)果顯著(P=0.005),進(jìn)一步將海蜇群體分成中國組群和日本族群兩組后計(jì)算的遺傳分化指數(shù)為0.418,且差異極顯著(P=0),這表明相距較遠(yuǎn)的海蜇地理種群間具有明顯的遺傳分化,這與利用擴(kuò)增片段長(zhǎng)度多態(tài)性技術(shù)[16]和利用內(nèi)轉(zhuǎn)錄間隔區(qū)序列[17]分析黃、渤海海蜇群體發(fā)現(xiàn)群體間無明顯遺傳分化的結(jié)果不一致。系統(tǒng)樹與單倍型最小跨度樹的結(jié)果亦表明,來自渤海的海蜇群體和黃海、日本海的海蜇群體間具有2個(gè)明顯的單倍型譜系分支,其中日本海的單倍型全部屬于類型Ⅰ,黃海的單倍型大部分屬于類型Ⅱ,這說明海蜇不同地理群體間尤其是相距較遠(yuǎn)的群體間可能存在遺傳分化。海蜇是河口性近海種類,具有水螅體世代、水母體世代交替出現(xiàn)的世代交替生活史,海蜇水母體世代生命周期一般為3—5個(gè)月,為季節(jié)性的浮游生物,而其水螅體世代生命周期至少為7—9個(gè)月,如果條件適宜甚至可以存活幾年時(shí)間,占據(jù)其生活史大部分的階段[1],水螅體階段主要營(yíng)固著生活,這種獨(dú)特的生活習(xí)性決定了其移動(dòng)能力十分有限,主要是隨海流的作用隨波逐流。因此,相距較遠(yuǎn)的地理群體間由于長(zhǎng)期的隔離分化會(huì)產(chǎn)生明顯的群體遺傳結(jié)構(gòu),這種現(xiàn)象也存在于馬賽克水母(Catostylusmosaicus)及海月水母(Aureliaaurita)的不同地理群體間[26]。
此外,本文的研究結(jié)果與利用擴(kuò)增片段長(zhǎng)度多態(tài)性技術(shù)和內(nèi)轉(zhuǎn)錄間隔區(qū)序列對(duì)海蜇群體進(jìn)行研究結(jié)果不同的原因,一方面可能是由于COⅠ屬于線粒體基因標(biāo)記,而擴(kuò)增片段長(zhǎng)度多態(tài)性和內(nèi)轉(zhuǎn)錄間隔區(qū)序列屬于核基因標(biāo)記,兩者間的遺傳方式、進(jìn)化速率等不同導(dǎo)致研究結(jié)果出現(xiàn)差異;另一方面擴(kuò)增片段長(zhǎng)度多態(tài)性[16-17]所分析的海蜇樣品主要為黃海采集的海蜇放流捕撈群體及養(yǎng)殖群體,樣品不同也可能是導(dǎo)致研究結(jié)果不同的重要原因[19]。
綜上所述,本文利用COⅠ基因部分片段對(duì)遼東灣野生海蜇群體進(jìn)行的分析表明遼東灣海蜇自然群體的遺傳多樣性尚處于中等或較高水平,分布于不同海域的海蜇群體間存在顯著的遺傳分化,這與利用擴(kuò)增片段長(zhǎng)度多態(tài)性技術(shù)和內(nèi)轉(zhuǎn)錄間隔區(qū)序列對(duì)海蜇群體進(jìn)行研究的結(jié)果不一致,樣品不同是導(dǎo)致研究結(jié)果不一致的重要原因,因此還需要利用不同的分子標(biāo)記對(duì)更多的海蜇樣品進(jìn)行分析,以期對(duì)海蜇這一重要經(jīng)濟(jì)水母的種群遺傳結(jié)構(gòu)及分子系統(tǒng)進(jìn)化關(guān)系有更加全面準(zhǔn)確的認(rèn)識(shí)。
[1] 董婧,姜連新,孫明,等.渤海與黃海北部大型水母生物學(xué)研究[M].北京:海洋出版社,2013.
[2] 內(nèi)田亨.日本動(dòng)物分類:缽水母綱[M].東京:三省堂,1936.
[3] Kramp P L. Synopsis of the medusae of the world[J]. J Mar Biol Ass UK, 1961, 40(1):467-469 .
[4] 洪惠馨,張士美,王景池.海蜇[M].北京:科學(xué)出版社,1978.
[5] 姜連新,葉昌臣,譚克非,等.海蜇的研究[M].北京:海洋出版社,2007.
[6] 丁耕蕪,陳介康.海蜇的生活史[J].水產(chǎn)學(xué)報(bào),1981,5(2):93-102.
[7] 陳介康,丁耕蕪.溫度對(duì)海蜇橫裂生殖的影響[J].動(dòng)物學(xué)報(bào),1983,29(3):195-206.
[8] 陳介康,丁耕蕪,劉春洋.營(yíng)養(yǎng)條件對(duì)海蜇橫裂生殖的影響[J].水產(chǎn)學(xué)報(bào),1985,9(4):321-329.
[9] 陳介康.海蜇的培育與利用[M].北京:海洋出版社,1985.
[10] 郭平.營(yíng)養(yǎng)條件對(duì)海蜇螅狀體形成足囊及足囊萌發(fā)的影響[J].水產(chǎn)學(xué)報(bào), 1990, 14(3):206-211.
[11] 趙斌,張秀梅,陳四清,等.環(huán)境因子對(duì)海蜇早期幼體發(fā)育影響的生態(tài)學(xué)研究進(jìn)展[J].海洋水產(chǎn)研究,2006, 27(1):87-92.
[12] 梁維波, 姜連新,于深禮.遼寧近海漁場(chǎng)海蜇增殖放流的回顧與發(fā)展[J].水產(chǎn)科學(xué),2007,26(7):423-424.
[13] 游奎,馬彩華,王紹軍,等.短期低溫條件對(duì)海蜇橫裂生殖的影響[J].水生生物學(xué)報(bào),2010,34(6):1223-1227.
[14] 王永順,黃嗚夏.海蜇增養(yǎng)殖技術(shù)[M].北京:金盾出版社,2004.
[15] 高祥剛,曹潔,董婧,等.遼寧沿海海蜇與沙海蜇遺傳多樣性的AFLP分析[J].水產(chǎn)科學(xué),2009, 28(2):75-78.
[16] Qiao H J, Liu X Q, Zhang X J, et al. Genetic diversity of populations and clones ofRhopilemaesculentumin China based on AFLP analysis[J]. Chinese Journal of Oceanology and Limnology, 2013, 31(2):391-397.
[17] 孫國華,劉相全,楊建敏,等.海蜇養(yǎng)殖群體及自然捕獲群體ITS序列遺傳分析[J].海洋科學(xué),2010, 34(10):90-95.
[18] Stopar K, Ramsak A, Trontelj P, et al. Lack of genetic structure in the jellyfishPelagianoctiluca(Cnidaria:Scyphozoa:Semaeostomeae) across European seas [J]. Mol Phylogenet Evol, 2010, 57(1):417-428.
[19] Andreja R, Katja S, Alenka M. Comparative phylogeography of meroplanktonic species,Aureliaspp. andRhizostomapulmo(Cnidaria:Scyphozoa) in European seas[J]. Hydrobiologia,2012,690(1):69-80.
[20] Armani A, Tinacci L, Giusti A, et al. What is inside the jar forensically informative nucleotide sequencing (FINS) of a short mitochondrial COⅠ gene fragment reveals a high percentage of mislabeling in jellyfish food products[J]. Food Research International, 2013, 54(2):1383-1393.
[21] Librado P, Rozas J. DnaSP v5: a software for comprehensive analysis of DNA polymorphism data[J]. Bioinformatics, 2009, 25(11):1451-1452.
[22] Excoffier L, Laval G, Schneider S. Arlequin ver 3.01: an integrated software package for population genetics data analysis[J]. Evolutionary Bioinformatics Online, 2005(1):47-50.
[23] Kumar S, Tamura K, Nei M. MEGA 3: integrated software for molecular evolutionary genetics analysis and sequence alignment [J]. Briefings in Bioinformatics, 2004, 5(2):150-163.
[24] Excoffier L, Smouse P E, Quattro J M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: application to human mitochondrial DNA restriction data [J]. Genetics,1992,131(2):406-425.
[25] Bandelt H, Forster P, Rohl A. Median joining networks for inferring intraspecific phylogenics [J]. Molecular Biology Evolution, 1999, 16(1):37-48.
[26] Dawson M N, Gupta A S, England M H. Coupled biophysical global ocean model and molecular genetic analyses identify multiple introductions of cryptogenic species[J]. Proceedings of the National Academy of Sciences USA, 2005, 102(34):11968-11973.
GeneticDiversityAnalysisofJellyfishRhopilemaesculentuminLiaodongBayBasedontheCOⅠSequence
LI Yulong, WANG Bin, WANG Wenbo, DONG Jing
( Key Laboratory of Marine Biological Resources and Ecology, Liaoning Province, Liaoning Ocean and Fisheries Science Research Institute, Dalian 116023,China )
The 624 bp fragments of mitochondrial cytochrome oxidase Ⅰ(COⅠ) gene were sequenced and analyzed in 20 individuals jellyfishRhopilemaesculentumcollected from Liaodong Bay to investigate the genetic variation and diversity. In addition, the homologous sequences were analyzed in other 15 individuals sampled from the Bohai and Yellow Seas and Sea of Japan. The average contents were found to be 26.7% in A, 36.4% in T, 18.8% in C and 18.1% in G for COⅠ sequence. A total of 21 polymorphic nucleotide sites were detected, which defined 17 haplotypes. Compared with several other giant jellyfish species, the variation level of the jellyfish was moderate to high withHof 0.91 ± 0.03 and π of 0.0056±0.0032. With the aid of the homologous sequences retrieved from GenBank, both phylogenetic and molecular variance analysis supported significant differentiation among jellyfish populations in Sea of Japan and the Bohai Sea and Yellow Sea populations.
Rhopilemaesculentum; mtDNA; COⅠ gene; genetic diversity
S917
A
1003-1111(2016)04-0404-06
10.16378/j.cnki.1003-1111.2016.04.016
2015-5-12;
2016-03-01.
國家自然科學(xué)基金資助項(xiàng)目(31400406);國家海洋公益性行業(yè)科研專項(xiàng)項(xiàng)目(201405010).
李玉龍(1981—),男,助理研究員;研究方向:海洋生物分子生物學(xué)及漁業(yè)資源增殖放流.E-mail: liyudragon@126.com. 通訊作者:董婧(1966—),女,研究員;研究方向:水母生態(tài)學(xué)及漁業(yè)資源.E-mail: 1024470248 @qq.com.
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國語大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學(xué)報(bào)(2017年2期)2017-07-05 08:13:02
國際商務(wù)財(cái)會(huì)(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
