油樟內生溶磷菌的篩選及其生物學特性
2016-12-22 09:47:46馮瑞章周誥均周萬海范軼玲
廣西植物 2016年11期
關鍵詞:植物
馮瑞章, 周誥均, 魏 琴, 周萬海*, 范軼玲, 秦 歡
( 1. 宜賓學院生命科學與食品工程學院 香料植物資源開發與利用四川省高校重點實驗室,四川 宜賓 644000; 2. 四川省高縣月江森林經營所, 四川 宜賓 645152 )
?
油樟內生溶磷菌的篩選及其生物學特性
馮瑞章1, 周誥均2, 魏 琴1, 周萬海1*, 范軼玲1, 秦 歡1
( 1. 宜賓學院生命科學與食品工程學院 香料植物資源開發與利用四川省高校重點實驗室,四川 宜賓 644000; 2. 四川省高縣月江森林經營所, 四川 宜賓 645152 )
篩選具有溶磷能力的植物內生細菌,并探索該類菌的促生和抗逆性能, 有助于擴大溶磷微生物來源、研發微生物肥料、改善土壤磷素營養和提高農業產量。該研究以從油樟組織中分離得到的50株內生細菌為材料,通過溶磷圈法初篩得到24株具有溶磷潛能的菌株,利用鉬藍比色法測定它們的溶磷能力和培養液的pH值,并研究溶磷能力較強菌株產生吲哚乙酸(IAA) 、鐵載體、1-氨基環丙烷-1-羧酸(ACC)脫氨酶、幾丁質酶等促生和抗逆性能。結果表明:24株油樟內生細菌都能從磷酸鈣中釋放出有效磷(溶磷量為51.26~237.08 μg·mL-1),其中,YG60、YG43、YG36、YG25、YG49、YG44株菌的溶磷量較高,均大于150 μg·mL-1。接種油樟內生菌后,培養液的pH值均有一定程度的降低,但菌株溶磷量與培養液pH值間不存在顯著相關性。6株溶磷量大于150 μg·mL-1的菌株大部分具有分泌IAA、產鐵載體、ACC脫氨酶活性和幾丁質酶活性的能力;其中YG43、YG60和YG25分泌IAA的能力較強 (IAA分泌量分別為22.55、18.75和16.41μg·mL-1),YG43和YG60產鐵載體的能力較強(As/Ar小于0.6),YG43、YG60和YG25的ACC脫氨酶活性(分別為0.194、0.224、0.208 U·mg-1)較高,YG43和YG60的幾丁質酶活性(分別為2.968 U和2.502 U)較高。綜合菌株的溶磷、促生和抗逆性能,認為YG43、YG60和YG25 菌株在促進植物生長、提高植物抗性及生物防治方面具有較好的應用前景。
溶磷內生菌, 溶磷能力, IAA, 鐵載體, ACC脫氨酶, 幾丁質酶
磷是植物生長發育所必需營養元素之一,植物缺磷直接影響植物的生長和作物的產量。目前,我國有74%的耕地土壤缺磷。化學磷肥的施加雖然能夠改善植物磷素營養,但由于磷在土壤中的固定導致磷肥當季利用率較低(黃靜等,2010)。在我國南方的紅、黃壤中缺磷現象更為嚴重(王同等,2014)。研究表明接種溶磷微生物能提高植物磷素營養(Illmer & Schinner,1992)。但是,由于受到土著微生物、土壤環境等因素的影響,土壤中溶磷微生物難以成功定殖,導致其應用效果不穩定(Reyes et al,2006;黃靜等,2010)。
內生菌作為植物植株整個微生態系統的重要組成部分,具有固氮、增強植物抗逆性、抗病蟲害及促進植物生長等多種作用(黃靜等,2010);同時大多數植物內生菌屬兼性內生,它們不僅存在于植物體內,而且也常見于根際土壤等環境中 (Lodewyckx et al,2002)。油樟(Cinnamomumlongepaniculatum)為樟科(Lauraceae)樟屬(Cinnamomum)的常綠喬木,也是重要的經濟作物,其根、莖、葉、種子均可作為提取芳香油的原料。調查顯示,油樟本身幾乎不發生植物病害,亦有研究表明油樟油對水稻稻瘟病菌等6種植物病原真菌的菌絲生長、孢子萌發有強烈的抑制作用(魏琴等,2006),這些特性可能與內生菌參與構成油樟植株內部微生態環境有關(游玲等,2009)。因此,以油樟為材料,對其組織內的溶磷細菌進行分離篩選,利用內生細菌的兼性特性,既可使這些微生物定殖于植物體內發揮促生和防病效應,也可使其定殖于植物根際土壤中發揮溶磷功能,為植物生長提供磷素營養,從而促進植物生長。
本研究以50株油樟內生細菌為材料,篩選具溶磷能力的菌株,研究其溶磷能力和生物學特性,為植物內生菌在提高植物磷素營養、促進植物生長等方面的利用提供依據。
1 材料與方法
1.1 材料
供試菌株為分離自油樟(Cinnamomumlongepaniculatum)組織中的50株內生細菌,于宜賓學院香料植物資源開發與利用四川省高校重點實驗室保存。
1.2 方法
1.2.1 油樟內生溶磷菌株篩選
1.2.1.1 內生溶磷菌株初選 將供試菌株活化后點接種至固體PKO培養基上,每株菌重復4次,28 ℃培養 7 d ,觀察能否有溶磷圈生成,測定各菌株的溶磷圈直徑和菌落直徑,分別以D、d表示,根據菌株是否產生溶磷圈初步判定該菌株有無溶磷潛能。
1.2.1.2 油樟內生溶磷菌株搖瓶復選 將50 mL PKO液體培養基置于150 mL三角瓶,121 ℃,1.01 P條件高壓滅菌25 min,備用。將LB斜面培養基上生長24 h的供試菌株制成菌數約為 108cell·mL-1菌懸液,按菌懸液與培養基1∶50比例接種,不接菌作為對照,每株菌重復3次,28 ℃、160 r·min-1搖床培養8 d,4 ℃、10 000 r·min-1離心15 min,取一定量的上清液,測其有效磷含量和pH值,有效磷含量即為菌株的溶磷量。有效磷含量用鉬銻抗比色法測定,菌株溶磷量為扣除不接種對照的值(μg P·mL-1);pH值用酸度計測定。
1.2.2 油樟內生溶磷菌株分泌IAA能力
1.2.2.1 定性測定 將篩選出的溶磷效果較好的菌株接種于盛有15 mL King 液體培養基的試管中,28 ℃、160 r·min-1搖床培養8 d,每株菌重復3次。藥品選擇和配置方法參考李振東等(2010)。分別取各菌株培養液50 μL滴置于白色的陶瓷板上,加入50 μL PC 比色液。以在比色液中加等量 50 mg·L-1生長素(IAA),不加菌株培養液作為對照。室溫條件下15 min內觀察顏色變化,顏色變為粉紅色表示可以分泌IAA,顏色越深表示分泌IAA濃度越大,不變色則表示不能分泌IAA。
1.2.2.2 定量測定 采用Glickmann et al (1995)的Salkowski比色法:配置2.5 mg·mL-1的色氨酸溶液過濾除菌,取1 mL加入4 mL已高壓滅菌的LB液體培養基中,使色氨酸濃度達到0.5 mg·mL-1,分別接種各供試菌株,28 ℃、160 r·min-1搖床培養3 d,4 ℃、10 000 r·min-1離心10 min后取上清液1 mL,加入50 μL 的10 mmol·L-1正磷酸,加入2 mL Salkowski’s顯色劑,黑暗條件(25 ℃)顯色30 min,測定530 nm下的吸光值。以蒸餾水為對照調零,配制不同濃度的IAA作標準曲線,計算培養液的IAA濃度,以μg·mL-1表示。
1.2.3 油樟內生溶磷菌株產鐵載體能力的測定
1.2.3.1 定性篩選 將篩選出的溶磷效果較好的菌株分別接種于鉻天青(chromea zural S,CAS) 平板培養基上,28 ℃培養2 d后觀察是否有橙紅色顯色圈出現,并測量顯色圈大小。
1.2.3.2 定量測定 將產生橙紅色顯色圈的菌株分別接種于MKB培養液和富鐵培養液中,28 ℃、160 r·min-1搖床培養24 h,接種1 mL發酵液于MKB和富鐵培養液中,28 ℃、160 r·min-1搖床培養36 h,4 ℃、10 000 r·min-1下離心10 min,即可得到鐵載體上清液和其無鐵載體對照的上清液,備用。取鐵載體上清液于潔凈的試管中,加入相同體積CAS檢測液,充分混勻,于630 nm波長下測定吸光值,以As表示,取雙蒸水作為對照調零;另取相同體積的富鐵培養液(對照液)與CAS檢測液反應,于630 nm波長下測定吸光值,以Ar表示, 計算As與Ar的比值(As/Ar),參照Machuca & Milagres(2003)的標準和方法檢測各菌株產鐵載體的能力。
1.2.4 油樟內生溶磷菌株ACC脫氨酶能力的測定
1.2.4.1 定性測定 參考Penrose & Glick(2003)的方法,分別接種各供試菌株于5 mL的無氮液體培養基中,28 ℃、160 r·min-1搖床培養24 h;接種上述培養液0.1 mL于5 mL DF液體培養基中,28 ℃、160 r·min-1搖床培養24 h;接種0.1 mL培養液于5 mL ADF液體培養基中,28 ℃、160 r·min-1搖床培養24~48 h,以不接種菌株的ADF培養基為對照;將ADF培養基中的生長菌株重復轉接和培養3次,能夠生長的菌株為ACC脫氨酶陽性菌株。
1.2.4.2 定量測定 參照Seleh et al(2001)的方法,每分鐘形成1 mol 丁酮酸的量為1個酶活力單位。采用Bradford法測定蛋白質,標準蛋白為牛血清白蛋白。酶活力的計算參照黃蓋等(2013)的方法,酶活力用U·mg-1表示。
1.2.5 油樟內生溶磷菌株產產幾丁質酶的測定
1.2.5.1 定性測定 分別接種各供試菌株于幾丁質平板培養基上,37 ℃培養3~5 d,觀察菌落周圍是否有透明圈產生,有則表明該株菌有產幾丁質的能力,反之則沒有。
2 結果與分析
2.1 內生溶磷菌株篩選及溶磷能力
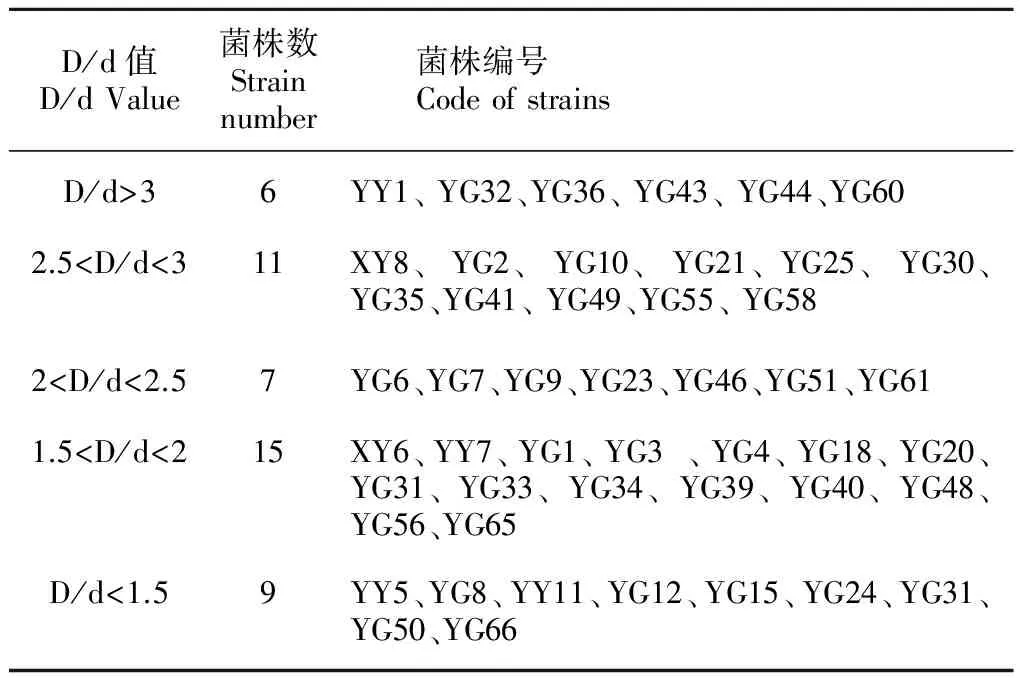
通過溶磷圈法對前期分離純化得到的50株油樟內生細菌進行初篩,發現菌株在PKO固體培養基上產生的透明圈差距較大。如表1所示,培養7 d后,D/d>3的細菌有6株(菌株YG43的D/d值最大,為3.84), 占供試菌株數量的12%, 2.5
表 1 不同 D/d 值菌株分布
Table 1 Distribution of strains with different D/d values
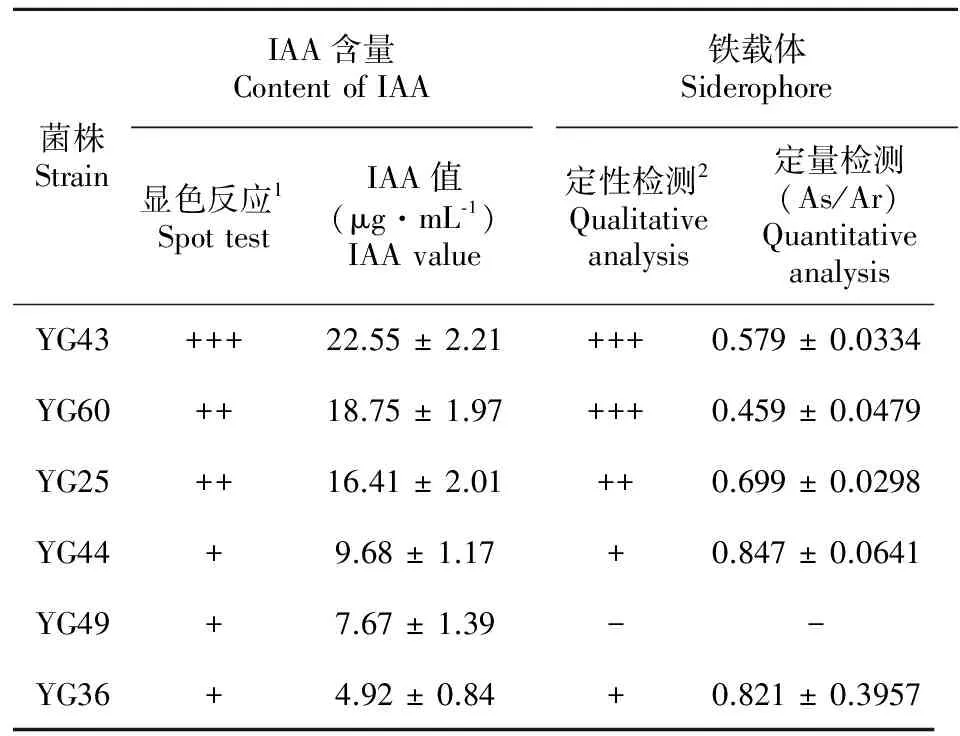
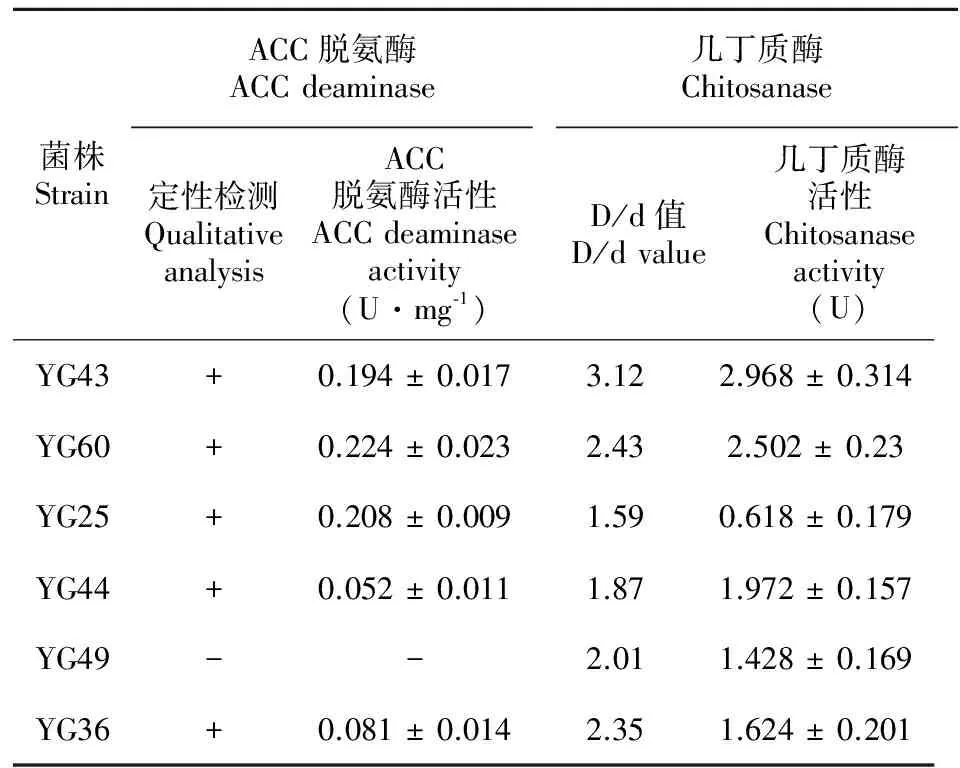
D/d值D/dValue菌株數Strainnumber 菌株編號 CodeofstrainsD/d>36YY1、YG32、YG36、YG43、YG44、YG602.5 將初選得到的24株菌分別液體培養8 d后,測定其上清液可溶性磷含量,由表2可知,菌株對磷酸鈣的溶解能力存在較大差異,供試菌株的溶磷量在51.26~237.08 μg·mL-1之間,其中大于200.00 μg·mL-1的有2株菌,150~200 μg·mL-1的有4株菌,100~150 μg·mL-1的有10株菌,分別占供試菌株的8.33%、16.67%和41.67%。菌株YG60的溶磷量最大,為237.08 μg·mL-1,是溶磷量最小菌株YG33(51.26 μg·mL-1)的4.62倍。此外,從各菌株的溶磷能力定性與定量結果可知,對油樟內生菌株而言,并非定性測定的D/d值越大,定量測定的溶磷量就越大,如YY1和YG60的D/d值均大于3,溶磷量卻分別為237.08 μg·mL-1和61.75 μg·mL-1,前者是后者的3.84倍。因此,固體平板培養只可對供試菌株是否具有溶磷能力作初步定性判斷,而液體培養測定的菌株溶磷能力在理論上更準確。此外,各菌株在PKO培養基上生長8 d 后,其培養液的pH值均較接種前明顯下降,各菌株培養液的pH值介于4.5~5.5,下降最大的為YG9,pH值下降為4.4。 表 2 菌株的溶磷量和培養液的pH值 Table 2 Available phosphorus and pH value of phosphate-solubilizing endophytic bacteria 菌株Strain溶磷量Availablephosphorus(μg·mL?1)pH值pHvalue菌株Strain溶磷量Availablephosphorus(μg·mL?1)pH值pHvalueYG60237.08±7.454.9YG38125.63±6.935.0YG43211.53±12.394.6YG30113.80±9.255.3YG36180.68±13.175.2YG23108.63±8.155.1YG25166.85±11.215.5YG6104.28±6.125.4YG49151.28±7.414.6YG996.23±4.314.4YG44150.20±5.234.6YG6191.60±10.755.3YG2145.90±8.494.8YG1883.50±5.824.7YG32143.30±10.275.1YY769.36±8.585.2XY8139.30±8.405.4YG467.11±6.407.0YG46137.00±9.295.2YY161.75±3.235.8YG35135.45±10.575.1YG5755.15±5.116.0YG7126.74±7.824.9YG3351.26±4.345.3 為進一步發掘油樟溶磷內生菌的功能,選擇溶磷量大于150 μg·mL-1的6株菌(分別為YG60、YG43、YG36、YG25、YG49、YG44)研究其生物學特性。 2.2 內生菌分泌IAA性能分析 油樟內生細菌分泌IAA能力的定性顯色反應結果表明(表3)。6株供試菌株不但具有較高的溶解難溶性磷的能力,同時均具有分泌生長素的功能,YG43的顯色反應為深紅色, YG60、YG25和YG44菌株呈粉紅色, YG49和YG36呈淺粉紅色。定量測定結果表明各菌株分泌IAA量在4.92~22.55 μg·mL-1之間,其中分泌IAA量超過20 μg·mL-1的有1株(YG43,22.55 μg·mL-1), 超過10 μg·mL-1的有2株(YG60、 YG25,IAA分泌量分別為18.75 μg·mL-1和16.41 μg·mL-1),其余3株的IAA分泌量均低于10 μg·mL-1,定量測定與定性測定結果相對一致。 表 3 菌株分泌 IAA和產鐵載體的性能 Table 3 IAA and siderophore producing ability of phosphate solubilizing endophytic bacteria 菌株StrainIAA含量ContentofIAA顯色反應1SpottestIAA值(μg·mL?1)IAAvalue鐵載體Siderophore定性檢測2Qualitativeanalysis定量檢測(As/Ar)QuantitativeanalysisYG43+++22.55±2.21+++0.579±0.0334YG60++18.75±1.97+++0.459±0.0479YG25++16.41±2.01++0.699±0.0298YG44+9.68±1.17+0.847±0.0641YG49+7.67±1.39--YG36+4.92±0.84+0.821±0.3957 注: 1. +++、++、 + 分別表示深粉紅色、粉紅色和淺粉色; 2. +++、++、+、-分別表示橙黃色暈圈較大、中等、較小和沒有。 Note: 1. +++、++、 + - + . means deep pink, pink and light pink respectively; 2. +++、++、+、- means larger, moderate, small and without orange-red hole respectively. 通過“拍立得”的推廣活動,我們可以得出這樣一個結論:在耳邊說一千遍的道理,都不如讓顧客親眼看到更有影響力。 2.3 內生菌產鐵載體能力分析 將6株油樟內生細菌接種于CAS平板上培養2 d后,檢測到除YG49之外的其它5菌株(YG43、YG60、YG25、YG44、YG36)的菌落周圍均出現橙黃色暈圈,說明這些菌株在CAS培養基上產生了鐵載體,其中YG43、YG60和YG25菌落周圍形成的橙紅色暈圈較大,表明這3株菌能夠產生較多的鐵載體。定量檢測鐵載體的方法以As與Ar的比值(As/Ar,Ar為對照吸光值)大小比較不同菌株產生鐵載體的能力,As/Ar值越小,說明菌株產鐵載體的能力越強。由表3可知,YG43和YG60的As/Ar值小于0.6,分別為0.579和0.458,說明這2株菌產生鐵載體的能力較強;YG25的As/Ar值為0.694,說明產鐵載體的能力中等;YG44和YG36的As/Ar值均大于0.8,說明這2株菌能產生鐵載體,但產鐵載體能力較弱。 2.4 內生菌產脫氨酶(ACC)能力分析 將溶磷能力較高的6株油樟內生細菌接種于SMA固體培養基后的ACC脫氨酶活性定性篩選實驗結果表明(表4),經5次傳代后YG49不能正常生長,其它5株菌(YG60、YG43、YG36、YG25、YG44)均能夠在SMA固體培養基上正常生長,說明除YG49外,其它菌株都具有ACC脫氨酶活性。定量檢測結果顯示(表4),各菌株的ACC脫氨酶活性大小順序為YG60>YG25>YG43> YG36> YG44,酶活大小介于0.052~0.224 U·mg-1;YG49無產ACC脫氨酶能力。 2.5 內生菌產幾丁質酶能力 將6株油樟內生溶磷細菌接種于產幾丁質酶的固體平板上培養7 d,發現均有明顯的透明圈產生,說明供試菌株皆有一定的產幾丁質酶的能力,但各菌株的透明圈差異較大,透明圈直徑(D)與菌落直徑(d)的比值(D/d)介于1.59~3.12。通過液體培養定量測定供試菌株的幾丁質酶活性的結果表明(表4),各菌株的幾丁質酶活性大小順序為YG60>YG25>YG43>YG36>YG44>YG49,酶活大小介于0.618~2.968 U·mg-1。 表 4 菌株ACC脫氨酶活性和幾丁質酶活性 Table 4 ACC deaminase activity and chitosanase activity of phosphate solubilizing endophytic bacteria 菌株StrainACC脫氨酶ACCdeaminase定性檢測QualitativeanalysisACC脫氨酶活性ACCdeaminaseactivity(U·mg?1)幾丁質酶ChitosanaseD/d值D/dvalue幾丁質酶活性Chitosanaseactivity(U)YG43+0.194±0.0173.122.968±0.314YG60+0.224±0.0232.432.502±0.23YG25+0.208±0.0091.590.618±0.179YG44+0.052±0.0111.871.972±0.157YG49--2.011.428±0.169YG36+0.081±0.0142.351.624±0.201 注: +,陽性; -,陰性。 Note: +, positive; -, negative. 植物內生細菌因其對宿主的多種有益的生物學作用,逐漸成為國內外學者研究的熱點。作為我國特有的天然產芳香油植物,可經高溫蒸餾從油樟(Cinnamomumlongepaniculatum)葉片中提取幾十種不同沸點的化學物質,這些物質精加工后的產物均是國防、醫藥、輕工等方面重要的稀有原料。具有溶磷能力的油樟內生促生細菌的篩選及促生、抗逆等生物學特性的研究,對溶磷微生物資源收集,植物內生細菌與宿主相互作用機制的理解及土壤磷素營養改善途徑的開發都具有重要意義(黃靜等,2010)。 本研究從油樟內生細菌中初篩到24株具有溶磷潛能的菌株,液體培養結果顯示,溶磷量大于200.00 μg·mL-1的菌株有2株,150~200 μg·mL-1的菌株有4株,黃靜等(2010)從玉米葉中分離的內生細菌溶磷量高達537.6 μg·mL-1,姚玉玲等(2014)從矮生嵩草中分離的內生細菌溶磷量為60.52 μg·mL-1,說明油樟內生細菌對無機磷的溶解能力較強。微生物在溶磷過程中會分泌如甲酸、草酸、蘋果酸等在內的各類有機酸,這些酸類物質既可降低培養基質的pH 又能與鈣、鐵、鋁等離子形成螯合物從而使難溶性磷酸鹽溶解(Rashid et al,2004)。本研究中24株菌在PKO培養基上生長8 d 后,其培養液的pH值較接種前均有明顯下降,但菌株溶磷量與培養液pH值之間無顯著線性關系(P>0.05)。表明內生溶磷細菌在培養期間,介質的酸度有一定降低,但培養液中的有機酸含量及種類、磷酸酶活性及產生多糖等的動態變化,均會對菌株的溶磷能力產生影響(李顯剛等,2012)。 植物內生菌可通過自身合成或者促進植物合成如生長素(IAA)、赤霉素(GA)、細胞分裂素(CTKs) 、脫落酸 (ABA)和乙烯等植物生長激素促進植物體的生長發育,其中生長素(IAA)是最活躍的成分(楊波等,2013)。一定濃度的IAA既可直接促進植物生長,又能通過增加細胞的體積和質量、改變細胞的內環境達到促生的目的(李顯剛等,2012)。本研究發現,6株溶磷能力較強的油樟內生細菌均具有分泌生長素的功能,且3株分泌IAA的能力較強(YG43、YG60和YG25),這一結果對油樟內生細菌作為促生菌促進植物生長開辟了新的途徑。 鐵載體的產生是植物內生細菌促進宿主生長發育的另一條重要途徑,植物內生微生物或者植物根際通過分泌產生多種對鐵具有高親和力的鐵載體,鐵載體可將Fe3+還原為植物體可以高效吸收和利用的Fe2+,溶解和結合土壤中的鐵元素供植物細胞利用;同時微生物產生的鐵載體通過與植物根際的病原微生物爭奪有限的鐵營養,抑制病原微生物的生長和繁殖,從而起到生物防治作用(Buyer et al,1993)。目前還無可作為各類微生物鐵載體的標品用于其標準曲線的測定,因此,國際上通用As/Ar 值作為微生物鐵載體定量檢測的指標(何苗等,2011)。本研究中,5株油樟內生細菌能夠產生鐵載體,且YG43和YG60的As/Ar值小于0.6,產鐵載體的能力較強。因此,產鐵載體可能是油樟內生細菌的促生機理之一。 ACC(1-aminocyclopropane-1-carboxylate,1-氨基環丙烷-1-羧酸)脫氨酶是許多植物促生細菌(Plant growth promoting bacteria,PGPB)共有的一個特征性酶,具有該種酶活性的內生細菌和植物根際對促進植物生長發育、提高植物抗高低溫、鹽、旱、澇、及重金屬等各種脅迫起到重要作用(Iniguez et al,2004)。黃蓋等(2013)從苜蓿根際分離的ACC 30菌株的酶活力為0.217 U·mg-1,該菌株能夠促進苜蓿根的伸長。本研究中除YG49外,其它5株菌均具有ACC脫氨酶活性,且YG49、YG60和YG25的酶活力在0.2 U·mg-1左右。因此,油樟內生細菌具有極大的促生、抗逆潛能。 幾丁質酶水解真菌細胞壁幾丁質產生的中間產物幾丁寡糖可以作為植物功能調節劑,刺激植物生長,誘導植物產生防御反應,從而提高植物的抗病性(郭玉蓮,2005)。此外,幾丁質酶與β-1,3-葡聚糖酶、其它PRP等防衛蛋白存在協同作用,可以增大作用范圍,從而提高抑菌效果。本研究中的6株油樟內生細菌皆有較高的產幾丁質酶的能力,酶活大小介于0.618~2.968 U·mg-1,這些內生細菌產生的幾丁質酶對植物病蟲卵的入侵和感染起到重要的防治作用(Barboza-Corona et al,2009),這也可能是油樟生長中幾乎不發生植物病、蟲害的原因之一。研究表明,真菌和細菌的幾丁質酶活性最高,細菌對真菌的抑制作用上最強(陳三鳳和李季倫,1994),因此,本研究可豐富幾丁質酶產生菌及生防菌資源。 通過溶磷圈法和液體培養法從50株油樟內生細菌中篩選到6株(YG60、YG43、YG36、YG25、YG49、YG44)溶磷能力較強的菌株,綜合各菌株的溶磷、促生和抗逆性能,認為YG43、YG60和YG25 菌株在促進植物生長、提高植物抗性及生物防治方面具有較好的應用前景。 BARBOZA-CORONA JE,ORTIZ-RODRIGUEZ T,FUENTE-SALCIDO N,et al, 2009. Hyperproduction of chitinase influences crystal toxin synthesis and sporulation of [J]. Anton Leeuwenhoek,96(1):31-42. BUYER JS,KRATZKE MG,SIKORA LJ, 1993. A method for detection of pseudobactin,the siderophore produced by a plant-growthpromoting Pseudomonas strain, in the barley rhizosphere [J]. Appl Environ Microbiol,59(3):677-681. CHEN SF,LI JL, 1994. Purification and properties of Chitinase fromFlavobacteriumsp. [J]. Acta Microbiol Sin, 34(1):14-19. [陳三鳳,李季倫, 1994. 黃桿菌(Flavobacteriumsp)幾丁質酶的純化和性質 [J]. 微生物學報, 34(1):14-19.] GLICKMANN E, DESSAUX Y, 1995. A critical examination of the specificity of the salkowski reagent for indolic compounds produced by phytopathogenic bacteria [J]. Appl Environ Microbiol, 619(2): 793-796. GUO YL, 2005. Chitinase and its effects on plant disease prevention [J]. Chin Agric Sci Bull,21(1):283-285. [郭玉蓮, 2005. 微生物幾丁質酶及其在植物病害防治中的作用 [J]. 中國農學通報,21(1):283-285.] HE M,HUANG Y,WANG J,et al, 2011. Isolation,characterization and mutation breeding of siderophore-producing bacterium MX-26 of peach rhizosphere [J]. Acta Phytophyl Sin,38(5):432-436. [何苗,黃云,王靖,等, 2011. 桃樹根際鐵載體產生菌MX-26的分離鑒定及誘變選育 [J]. 植物保護學報,38(5):432-436.]HUANG G,GAO H,WANG C,et al, 2013. ACC 30 strain with ACC deaminase activity:its isolation,identification and growth-promoting effect [J]. Microbiol Chin,40(5):812-821. [黃蓋,高焓,王琛,等, 2013. ACC脫氨酶活性菌株ACC 30的分離、鑒定及其促生作用 [J]. 微生物學通報,40(5):812-821.] HUANG J,SHENG XF,HE LY, 2010. Biodiversity of phosphate-dissolving and plant growthpromoting endophytic bacteria of two crops [J]. Acta Microbiol Sin,50(6) :710-716. [黃靜,盛下放,何琳燕, 2010. 具溶磷能力的植物內生促生細菌的分離篩選及其生物多樣性 [J]. 微生物學報,50(6) :710-716.] ILLMER P, SCHINNER F, 1992. Solubilization of inorganic phosphates by microorganisms isolated from forest soil [J]. Soil Biol Biochem,24(4):389-395. INIGUEZ AL,DONG Y,TRIPLETT EW, 2004. Nitrogen fixation in wheat provided byKlebsiellapneumoniae342 [J]. Mol Plant Microb Inter,17(10):1078-1085. LI XG,WANG XL,YAO T,et al, 2012. Characters of solubilizing phosphate,secreting IAA and organic acids of phosphorus-solubilizing bacteria from rhizosphere ofLotuscorniculatus[J]. Chin J Soil Sci,43(6):1385-1390. [李顯剛,王小利,姚拓,等, 2012. 溶磷菌的溶磷、分泌IAA及有機酸特性研究 [J]. 土壤通報,43(6):1385-1390.] LI ZD,CHEN XR,LI P,et al, 2010. Identification ofPolygonumviviparumendophytic bacteria Z5 and determination of the capacity to secrete IAA and antagonistic capacity towards pathogenic fungi [J]. Acta Pratac Sin,19 (2):61-68. [李振東,陳秀蓉,李鵬,等, 2010. 珠芽蓼內生菌Z5產IAA和抑菌能力測定及其鑒定 [J]. 草業學報,19 (2):61-68.] LODEWYCKX C,VANGRONSVELD J,PORTEOUS F,et al, 2002. Endophytic bacteria and their potential applications [J]. Crc Crit Rev Plant Sci,21:583-606. MACHUCA A,MILAGRES AMF, 2003. Use of CAS-agar plate modified to study the effect of different variables on the siderophore production by Aspergillus [J]. Lett Appl Microbiol,(36):177-181. PENROSE DM,GLICK BR, 2003. Methods for isolating and characterizing ACC deaminase-containing plant growth-promoting rhizobacteria [J]. Physiol Plant,118: 10-15. RASHID M,KHALIL S,AYUB N, 2004. Organic acids production and phosphate solubilization by phosphate solubilizing microorganisms (PSM) underinvitroconditions [J]. Pakistan J Biol Sci,7(2):187-196. REYES I,VALERY A,VALDUZ Z, 2006. Phosphate-solubilizing microorganisms isolated from rhizospheric and bulk soils of colonizer plants at an abandoned rock phosphate mine [J]. Plant Soil, 287:69-75. SELEH SS,GLICK BR, 2001. Involvement ofgacSandrpoSin enhancement of the plant growth-promoting capabilitiesEnterobactercloacaeCAL2 and UW4 [J]. Can J Microbiol,47(8):698-705. WANG T,KONG LY,JIAO JG,et al, 2014. Screeing of phosphate-solubilizing bacteria in Red soil and their acting mechanisms [J]. Aata Pedol Sin,51(2):373-380. [王同,孔令雅,焦加國,等, 2014. 紅壤溶磷菌的篩選及溶磷機制 [J]. 土壤學報,51(2):373-380.] WEI Q,LI Q,LUO Y,et al, 2006. Antifungal activity of leaf essential oil fromCinnamomumlongepaniculatum(Gamble) N. Chao [J]. Chin J Oil Crop Sci,28(1):63-66. [魏琴,李群,羅揚,等, 2006. 油樟油對植物病原真菌活性的抑制作用 [J]. 中國油料作物學報,28(1):63-66.] YANG B,CHEN Y,LI X,et al, 2013. Research progress on endophyte-promoted plant nitrogen assimilation and metabolism [J]. Acta Ecol Sin, 33(9):2656-2664. [楊波,陳晏,李霞,等, 2013. 植物內生菌促進宿主氮吸收與代謝研究進展 [J]. 生態學報, 33(9):2656-2664.] YAO YL,WANG Y,WANG YQ,et al, 2014. Identification of endophytic bacteria fromkobresiahumilisand determination of phoaphate-solubilizing, IAA secretion and antagonistic abilities [J]. Acta Agrest Sin,22(6):1252-1257. [姚玉玲,王穎,王玉琴,等, 2014. 矮生嵩草內生細菌溶磷抑菌和產IAA能力的測定及鑒定 [J]. 草地學報,22(6):1252-1257.] YI T,LIAO YX,FENG YJ, 2008. Plant-endophyte interaction: growth-promoting effect of endophytes and their biofilm formation [J]. Microbiol China,35(11):1774-1780. [易婷,繆煜軒,馮永君, 2008. 內生菌與植物的相互作用:促生與生物薄膜的形成 [J]. 微生物學通報,35(11):1774-1780.] YOU L,WANG T,LI L,et al, 2009. Analyses on volatile organic compound of 78 endophytic fungi isolated fromCinnamomumlongepaniculatum( Gamble) N. Chao ex H. W. Li [J]. J NW A & F Univ, 37(9):193-198. [游玲,王濤,李蘭,等, 2009. 78 株油樟內生真菌發酵產物的揮發性組分分析 [J]. 西北農林科技大學學報, 37(9):193-198.] Screening and characterization of phosphate dissolving endophytic bacteria fromCinnamomumlongepaniculatum FENG Rui-Zhang1, ZHOU Gao-Jun2, WEI Qin1,ZHOU Wan-Hai1*, FAN Yi-Ling1, QIN Huan1 ( 1.KeyLabofAromaticPlantResourcesExploitationandUtilizationinSichuanHigherEducation,CollegeofLifeSciences&FoodEngineering,YibinUniversity, Yibin 644000, Sichuan, China; 2.GaoCountyYuejiangForestManagementStation, Yibin 645152, Sichuan, China ) Screening endophytic strains with phosphate-dissolving and exploring the characterization of growth promoting and resistance were conducive to broad the phosphate dissolving microbial resources, to develop microbial fertilizers, to improve soil phosphorus nutritious and to increase agricultural yield. We isolated 50 endophytic bacteria from interior tissues ofCinnamomumlongepaniculatum, and 24 phosphate dissolving endophytic bacteria strains were screened using phosphate solubilizing zone (PKO) inorganic culture medium, subsequently, the capacity of dissolving phosphorus of 24 strains were determined by phosphor-molybdate blue color methods, and the characteristics benefited to plant-microbe interaction-indoleacetic acid (IAA), siderophore, 1-aminocyclopropane-1-carboxylic acid deaminase (ACC) and chitosanase activities were also evaluated. The results showed that 24 strains could release phosphate from tricalcium phosphate with the range of dissolving phosphate from 51.26 μg·mL-1to 237.0 μg·mL-1, and the maximum phosphate content in the solution was obtained with strain YG60. Among 24 strains, 6 strains (YG60, YG43, YG36, YG25, YG49, YG44) showed the higher phosphate solubilization capacity (237.08, 211.53, 180.68, 166.85, 151.28, 150.20 μg·mL-1respectively). Furthermore, 24 strains could decrease the pH of the medium with the pH value range from 4.5 to 5.5, and the relationship between pH and phosphorus solubizition was also discussed; however, there were no close relationship between them. The further experimental results showed that most of the 6 strains with higher phosphate dissolving capacity had the ability to produce IAA and siderophores, and had ACC deaminase and chitosanase activity. Among them, YG43, YG60 and YG25 possessed higher IAA producing capacity (22.55, 18.75 and 16.41 μg·mL-1respectively); YG43 and YG60 possessed higher siderophores of As/Ar<0.6 (0.459, 0.579 respectively ); YG43, YG60 and YG25 had strong ACC deaminase activity (0.194, 0.224, 0.208 U·mg-1),YG43 and YG60 had strong chitosanase activity (2.968 U,2.502 U). In terms of all the properties of dissolving phosphate, secreting IAA, siderophores, ACC deaminase and chitosanase, strains YG43, YG60 and YG25 isolated from interior tissues ofC.longepaniculatumhave abundant biological characteristics related to plant growth promotion, stress homeostasis regulation and biocontrol activity. They are possible to be further developed as excellent strains for application. phosphate-dissolving endophytic bacteria, capacity of dissolving phosphorus, IAA, siderophores, ACC deaminase, chitosanase 10.11931/guihaia.gxzw201506007 2015-09-10 2015-12-09 宜賓市科技創新專項(2012ZNY006);四川省大學生創新創業訓練計劃項目 (20151064b17); 四川省高校科研創新團隊項目(14TD0031);四川省青年科技創新研究團隊培育計劃項目(2011JTD0035) [Supported by Program for Science and Technology Bureau of Yibin City (2012ZNY006); Training Programs of Innovation and Entrepreneurship Undergraduates in Sichuan Province (201510641017); Scientific Research Fund of Sichuan Provincial Education Department (14TD0031); Sichuan Youth Science and Technology Innovation Teams (2011JTD0035)]。 馮瑞章(1978-),女,甘肅武威人,博士,副教授,研究方向為微生物資源開發利用,(E-mail)ruizhangfeng@126.com。 *通訊作者: 周萬海,博士,副教授,主要從事農業資源環境研究,(E-mail)wanhaizhou@126.com。 Q939.96 A 1000-3142(2016)11-1396-07 馮瑞章, 周誥均, 魏琴, 等. 油樟內生溶磷菌的篩選及其生物學特性 [J]. 廣西植物, 2016, 36(11):1396-1402 FENG RZ, ZHOU GJ, WEI Q, et al. Screening and characterization of phosphate dissolving endophytic bacteria fromCinnamomumlongepaniculatum[J]. Guihaia, 2016, 36(11):1396-1402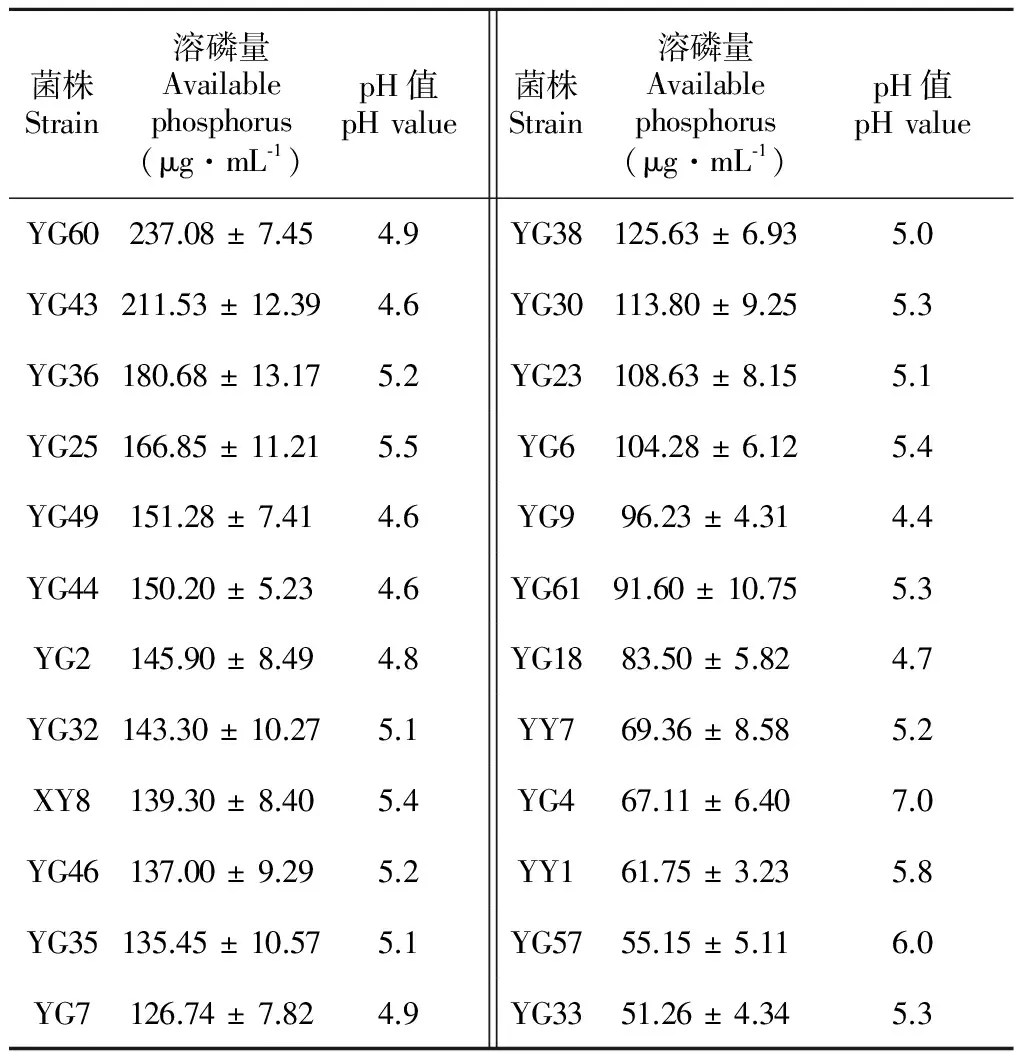
3 討論與結論
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
