羊草形態及生物量分配可塑性對土壤供磷能力的響應研究
2017-01-03 10:49:40郭豐輝侯向陽丁勇李西良武自念王珍胡靜李新樂馬文靜趙勁博
草業學報 2016年12期
郭豐輝,侯向陽*,丁勇,李西良 ,武自念,王珍,胡靜,李新樂 ,馬文靜,趙勁博
(1.中國農業科學院草原研究所,國家牧草改良中心,內蒙古 呼和浩特 010010;2.中國林業科學研究院沙漠林業實驗中心,內蒙古 磴口 015200)
?
羊草形態及生物量分配可塑性對土壤供磷能力的響應研究
郭豐輝1,侯向陽1*,丁勇1,李西良1,武自念1,王珍1,胡靜1,李新樂2,馬文靜1,趙勁博1
(1.中國農業科學院草原研究所,國家牧草改良中心,內蒙古 呼和浩特 010010;2.中國林業科學研究院沙漠林業實驗中心,內蒙古 磴口 015200)
植物形態及生物量分配變化是天然草原退化過程中的主要表現之一,通過研究植物形態及生物量分配對環境的響應規律有助于揭示草原退化及植被演替的內在機理。本試驗研究了羊草形態、生物量累積及分配對土壤供磷能力的可塑性響應。試驗采用有效磷含量低于3 mg P2O5/kg的土壤設置0,30,90,180 mg P2O5/kg 4個磷肥添加梯度,形成4種具有不同供磷能力的土壤,用于對羊草植株的培養,并于后期進行各項指標的測定。結果表明,1)土壤供磷能力的減弱明顯減小了羊草的株高、各構件生物量,表現為高磷添加量顯著高于低磷添加量,株高、葉生物量、莖生物量、根生物量、總生物量的降低幅度分別達到了14%,50%,51%,79%,63%。 2)羊草個體形態及生物量分配對土壤供磷能力的響應并不完全一致,生物量累積對供磷量的敏感度最大,生物量分配次之,形態的敏感度最差,可塑性指數分別為0.58,0.31,0.19;單個性狀指標以根干重最為敏感,可塑性指數達到了0.79。3)相關分析表明,羊草地上干重與葉寬、節間數、株高達到了極顯著水平,此3個形態指標對羊草地上生物量的貢獻率分別為30%,27%,26%,是形態指標中解釋羊草地上生物量產生差異的主要因子。本試驗在課題組前期研究的基礎上,進一步從個體水平上探索羊草對土壤供磷能力的響應,與課題組前期試驗結果相結合,可有助于探索天然草原退化和植被演替的內在原因。
羊草;土壤供磷能力;表型可塑性;可塑性指數
表型可塑性與植物對環境的適應能力、外來物種的入侵能力、物種的進化及生態系統中植被的演替息息相關[1-4]。植物在不同生存環境下的可塑性響應可以提高其入侵能力、拓寬其生存范圍。通過研究植物對環境因子(包括養分、水分、光照、溫度、二氧化碳等非生物因子及生物因子)的可塑性響應有助于揭示自然界中物種入侵、植被演替等現象的內在機理[5-8]。近些年有關植物表型可塑性的研究主要集中于植物對環境因子的可塑性響應與外來物種入侵能力、生態幅、物種進化的關系等方面[9-12]。也有學者研究了植被演替過程中植物的個體行為,試圖通過植物的個體行為來探討植被演替的機理[13-14],有關植物個體行為對環境的響應及其在生態系統中所起到的作用越來越受到人們的關注。
天然草原是陸地生態系統的重要組成部分,其功能的多樣性對于維持全球的生態平衡具有重要作用[15]。近幾十年來,由于人類的干擾及氣候環境的變化等因素,天然草原退化嚴重,表現為生產力降低、生態功能減弱等[16]。植物可以通過可塑性變化來適應其生長環境的改變,不同物種可塑性能力存在差異[17-18],這從一定程度上導致了植物不同的適應能力,進而影響到了天然草原植被的演替[13, 18-19]。鑒于植物表型可塑性在物種生存、群落演替中的重要作用,過去許多學者針對天然草原植物對放牧、土壤養分、土壤水分及種群密度等環境因素的表型可塑性做了大量研究,試圖通過對草原退化過程中植物的可塑性響應來揭示草原退化的原因。李西良等[3]、安慧[20]、李金花等[21]都對放牧干擾下植物的形態及生物量分配可塑性做了研究,發現放牧可以改變植物的高度、莖葉及生物量分配等性狀,而植物也可以通過形態可塑性的響應來適應放牧干擾;沙米(Agriophyllumsquarrosum)對土壤養分、水分和種群密度3個環境因子均有表型可塑性響應,但以對土壤養分的響應幅度最大[22]。總之,表型可塑性是草原生態系統植物個體的一種普遍行為,其對維持天然草原的組成結構與功能具有重要作用。
中國北方草原在退化過程中,土壤有效磷含量呈降低的趨勢,致使其成為該生態系統植物生長的限制因子[23]。同時,近些年來由于環境的污染,氮沉降量加大[24-25],這種氮輸入的增加一方面可以緩解天然草原植物的氮限制[26],另一方面也打破了植物對養分的需求平衡,而使得植物更易受到其他養分如磷元素的限制[27-28]。而當土壤有效磷含量成為限制因子時,植物一方面可以通過生理可塑性來適應磷的限制,如增加根系酸性磷酸酶、質子及有機酸的分泌量來提高根基土壤有效磷的濃度[29];另一方面,植物也可以發生形態可塑性,通過增加比葉面積、葉片大小來響應土壤供磷能力的變化[30]。已有研究表明羊草(Leymuschinensis)在草原退化過程中會出現植株變矮、莖稈變短、葉片變短等小型化響應[3,13,18-19],其對過度放牧的可塑性響應越來越受到人們的重視。
之前有關羊草對環境的可塑性研究大都是在群落水平下針對多個環境因子對羊草產生的綜合影響,不能確定過度放牧下羊草對每個環境因子的可塑性響應。課題組前期研究表明,羊草莖質量、總質量、莖高、株高、葉面積等為對放牧響應的敏感性狀,而葉片數、莖粗、葉寬等較為穩定,為惰性性狀;莖長、株高、葉面積、葉長等性狀的投影重要性指標大于1,對地上生物量變化的解釋率為68.6%,是導致長期放牧下羊草個體生物量降低的主要因子。本試驗在課題組前期關于放牧干擾下羊草的可塑性響應的基礎上,通過室內控制試驗來研究羊草對土壤供磷能力的可塑性響應,試圖揭示1)羊草對土壤供磷能力的可塑性響應規律;2)土壤供磷能力改變羊草個體地上生物量的驅動因子;3)結合課題組前期關于放牧對羊草形態可塑性的研究結果,嘗試初步探討土壤供磷能力在天然草原退化演替中的可能作用。
1 材料與方法
1.1 試驗材料
試驗于2015年5月初至7月初在農業部沙爾沁牧草資源重點野外科學觀測試驗站溫室進行。為了有效研究土壤中磷元素對天然草原植物所起的作用,避免其他營養成分偏離正常水平而對試驗造成影響,試驗所用土壤采自于中國農業科學院廊坊試驗基地,該土壤有效磷含量長期嚴重缺乏(該土壤有效磷含量為3.00 mg P2O5/kg,據徐明崗等[31]研究當土壤有效磷含量低于5 mg P2O5/kg時即為虧缺狀態),而其他養分水平相對正常[32]。土壤的基本理化性質為:全磷0.56 g/kg、全氮0.66 g/kg、有效磷3.00 mg P2O5/kg、堿解氮51.36 mg/kg、有機質1.11%、pH 7.50。將所有土壤充分混勻、過篩、風干,然后裝入直徑為18 cm的花盆,每盆2.25 kg風干土。為了保證羊草種子基因的一致性,減小處理內的變異性,羊草種子選用變異性較小的吉生4號,由試驗站羊草資源圃提供。
1.2 試驗方法
磷處理設置0,30,90,180 mg P2O5/kg四個梯度,每個處理6個重復,磷肥選用過磷酸鈣。將裝好土的花盆按照其施磷梯度計算好施磷量(每盆含風干土2.25 kg),然后將每盆所添加的磷肥充分與土壤混勻,最后給每盆澆同等量的水(第一次澆透為止,之后保持土壤最大持水量的45%左右)。
挑選大小一致的飽滿羊草種子進行種植,每盆均勻地撒下25粒種子,敷一層薄土。待出苗15 d左右間苗,每盆留長勢基本相同的幼苗12株。在羊草生長期間,定期對每盆澆同等量的水(根據植物不同生長時期,澆水量略有改變,幼苗期為促進其根系的生長澆水量少于其他時期),同時拔除花盆里所出現的少量雜草。
1.3 樣品采集與數據測定
生長到60 d時收獲,此時羊草處于生長旺盛時期的末期,生長速度明顯減慢。收獲時將花盆打破,輕輕地抖掉根系上面的土壤,然后將每株的根系分開,并洗凈。首先對植物的形態指標進行測定,從每盆的12株中隨機選取3株進行測定,這樣每個處理需對18個單株進行測定。利用游標卡尺測量羊草的葉寬及節間粗,對每片葉及每一節間進行測定后取平均值;利用鋼尺對植物的株高、節間長、葉長進行測定。待所有形態指標測量結束后,對每株進行根、莖、葉的分離,并置于60 ℃烘箱烘干,然后稱重。
1.4 數據處理
本研究中的性狀指標包括形態性狀:株高(plant height, PH)、節間數(internode number, SN)、節間長(internode length, SL)、節間粗(internode thick, SD)、葉長(leaf length, LL)、葉寬(leaf width, LW)、節間長/節間粗(internode length/internode thick, SLD)、葉長/葉寬(leaf length/leaf width, LLW);生物量累積性狀:葉干重(leaf mass, LM)、莖干重(stem mass, SM)、根干重(root mass, RM)、地上干重(aboveground biomass mass, ABM)、總干重(total mass, TM);生物量分配性狀:葉生物量比(leaf mass distribution, LMD)、莖生物量比(stem mass distribution, SMD)、根生物量比(root mass distribution, RMD),LMD、SMD、RMD分別為LM/TM、ABM/TM、RM/TM。

利用Excel 2010進行數據整理及簡單的數據分析,利用SPSS 19.0進行方差分析和相關分析,利用Sigmaplot 12.0作圖。方差分析法分析羊草各個性狀在不同處理間的差異顯著性,進行Duncun多重比較,數據以平均值±標準誤差表示;相關分析利用Pearson相關法,分析部分性狀之間的相關性,并在0.05水平上進行顯著性檢驗。
2 結果與分析
2.1 羊草對土壤供磷能力的形態可塑性響應
隨著土壤供磷能力的增強,羊草株高增高、節間數增多、葉片寬度增大而長度沒有變化,其中株高及節間數在不同處理間達到了顯著水平(P<0.05),葉片寬度在最低土壤供磷水平條件下顯著小于最高土壤供磷水平(P<0.05),葉片長度在每個處理間差異不顯著;羊草的節間長度、節間粗及節間長/節間粗、葉長/葉寬對土壤供磷能力產生了一定程度的響應,其中節間長、節間長/節間粗隨著供磷能力的增強呈增大趨勢,而節間粗、葉長/葉寬呈減小趨勢,這4個性狀指標在部分處理間也達到了顯著水平(P<0.05)(圖1)。

圖1 羊草形態可塑性(平均值±標準誤差)Fig.1 The morphological plasticity of L. chinensis (mean±SE) P0、P1、P2、P3分別代表磷添加量0、30、90、180 mg P2O5/kg。不同小寫字母表示差異顯著(P<0.05)。PH:株高;SN:節間數;SL:節間長;SD:節間粗;LL:葉長;LW:葉寬;SLD:節間長/節間粗;LLW:葉長/葉寬;下同。P0、P1、P2、P3 are the phosphorus concent 0, 30, 90, 180 mg P2O5/kg. Different small letters indicate significant differences (P<0.05). PH: Plant height; SN: Internode number; SL: Internode length; SD: Internode thick; LL: Leaf length; LW: Leaf width; SLD: Internode length/Internode thick; LLW: Leaf length/ Leaf width; The same below.
2.2 羊草生物量累積及分配對土壤供磷能力的可塑性響應
在生物量累積方面,隨著土壤供磷能力的增強,羊草各構件生物量呈增高趨勢。當土壤供磷能力最強時,葉干重、莖干重、根干重、地上干重和總干重均顯著高于其他處理(P<0.05);最低供磷能力的各生物量累積指標均顯著低于其他處理。隨著土壤供磷能力的改變,生物量分配規律明顯,在最低供磷能力下,羊草的葉生物量比及莖生物量比均顯著小于其他處理,地下生物量比顯著高于其他處理,各磷添加的處理之間差異不顯著(P<0.05)(圖2)。
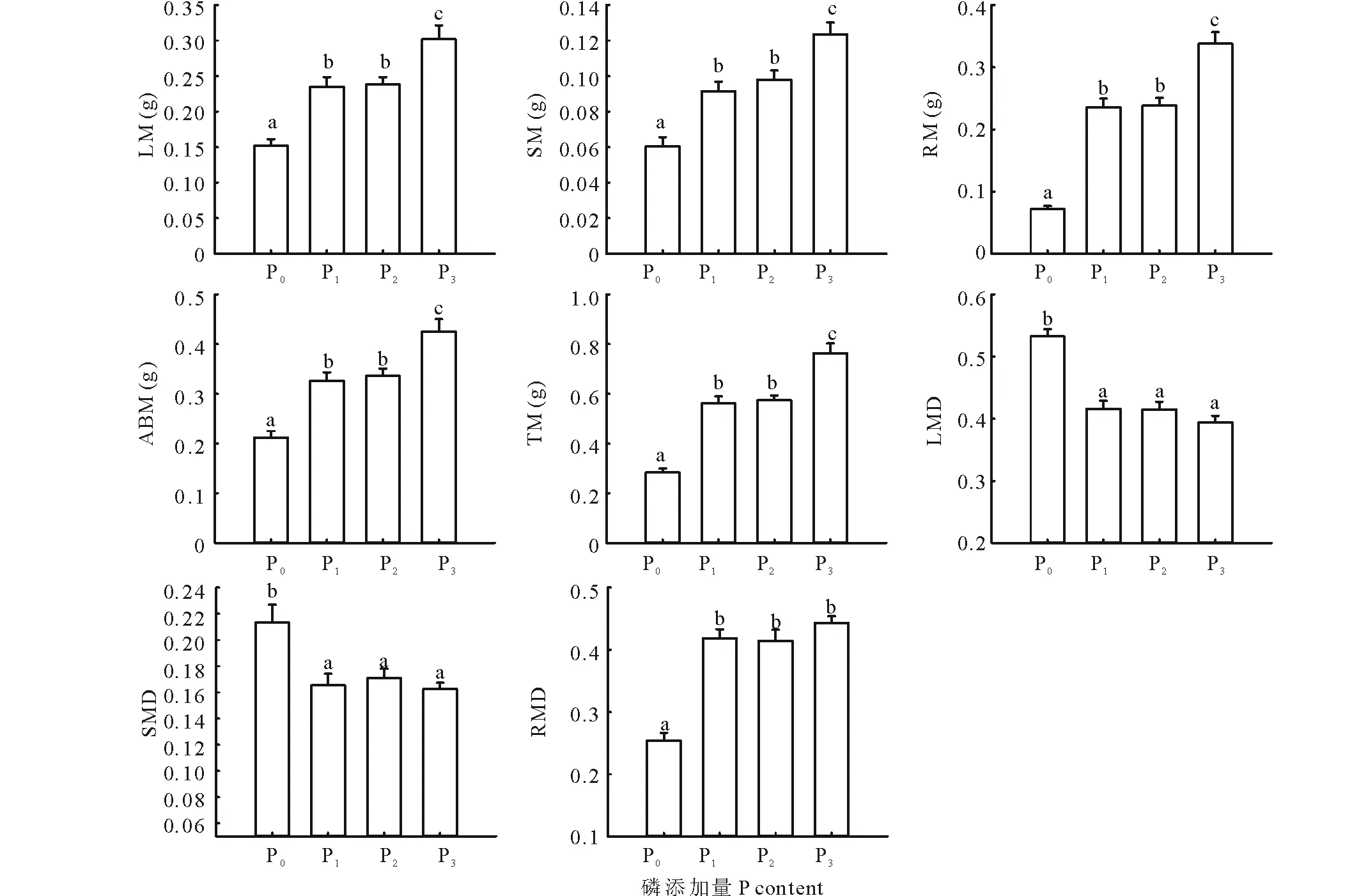
圖2 羊草生物量積累及分配可塑性(平均值±標準誤差)Fig.2 The plasticity of biomass accumulation and distribution of L. chinensis(mean±SE) LM:葉干重;SM:莖干重;RM:根干重;ABM:地上干重;TM:總干重;LMD:葉生物量比;SMD:莖生物量比;RMD:根生物量比;下同。LM: Leaf mass; SM: Stem mass; RM: Root mass; ABM: Aboveground biomass; TM: Total mass; LMD: Leaf mass ratio; SMD: Stem mass ratio; RMD: Root mass ratio; The same below.
2.3 羊草各性狀指標對土壤供磷能力的敏感度
為了比較羊草各性狀指標對土壤供磷能力敏感度的差異,對其各性狀可塑性指數進行了排序。結果表明,羊草可塑性指數最大的性狀為根干重(0.79);羊草的生物量累積指標可塑性指數平均值、生物量分配指標可塑性指數平均值、形態指標可塑性指數平均值分別為0.58,0.31,0.19,也就是說以其生物量累積對供磷量的敏感度最大,生物量分配次之,形態的敏感度最差(圖3)。
2.4 植株大小與形態指標之間的關系
以地上干重及株高表示植株個體的大小,分析了植株個體大小與各形態指標之間的相關關系。結果表明羊草形態性狀均與植株個體大小呈正相關關系,存在協同變化關系。羊草株高、節間數、葉寬與地上干重相關性達到了極顯著水平,節間數、節間長、葉寬與高也達到了極顯著水平(表1)。
為判斷羊草各形態指標對其地上生物量影響的大小,利用偏最小二乘法確定了各性狀因子的變量投影重要性指標(VIP),并計算了各形態指標對地上生物量的貢獻率。結果表明,在6個羊草個體的形態性狀中葉寬、節間數及株高的VIP都大于1,分別為1.48,1.36,1.28,是羊草個體地上生物量的主要影響因子。各形態指標對地上生物量貢獻率的大小比較為LW>SN>PH>SD>SL>LL(圖4)。其中葉寬、節間數、株高3個形態性狀對羊草地上生物量的貢獻率總和為83%,是解釋羊草個體地上生物量形成和變化的主要指標,而節間粗、節間長、葉長的貢獻率相對要低一些(總和為17%)(圖4)。

圖3 羊草可塑性指數排序Fig.3 Sorting of the plasticity index of L. chinensis
表1 羊草植株個體大小與其形態性狀的相關性分析
Table 1 The correlation of morphological trait and plant size forL.chinensis

性狀TraitPHSNSLSDLLLWABM0.588**0.629**0.1770.190-0.0270.688**PH10.538**0.494**-0.0970.1940.343**
注:數字代表相關系數,**代表顯著性(P<0.01)。
Note: The number is correlation index, ** indicates significant (P<0.01).

圖4 羊草個體地上生物量與形態指標的投影重要性指標(VIP)(A)及各形態指標貢獻率(B)Fig.4 The variable importance in projection values (VIP)(A) and the contribution rate of morphological traits to aboveground biomass(B)
3 討論
課題組前期研究了羊草個體形態及個體生物量對過度放牧的響應,結果表明,羊草對放牧干擾存在非常明顯的可塑性,是過度放牧下天然草原生物量減小、植被演替的原因之一[3]。過度放牧造成的土壤有效磷含量的降低是草原退化的表現之一,加之最近幾年環境中氮沉降的加劇,土壤供磷能力對植物生長的限制越來越受到人們的重視[23-25]。為了研究土壤供磷能力在過度放牧下草原退化過程中的作用,在課題組關于放牧下羊草的可塑性響應研究的基礎上,本試驗針對土壤供磷能力對羊草生長的影響進行了研究。結果表明,羊草的生長對土壤供磷能力非常敏感,表現為株高降低、各構件生物量減少,隨著土壤供磷能力的減弱,羊草單株生物量和株高分別降低了63%和14%,地下、莖、葉生物量分別降低了79%、51%、50%,結合放牧干擾下土壤有效磷含量狀況的變化,推測土壤供磷能力的減弱可能是過度放牧造成天然草原生產力降低、植被演替的原因之一。
天然草原植物在過度放牧干擾下,植株個體變矮、葉片變短變窄、節間縮短、叢幅變小、根系分布淺層化,表現為植株矮小化[13,18]。有研究表明,在放牧干擾下,羊草株高、葉長、節間長為敏感性狀,莖粗、葉寬及葉片數的敏感性相對較低[3,13,34-36],這與羊草植株個體形態對土壤供磷能力的響應規律有所不同。本研究結果表明隨著土壤供磷能力的改變,羊草個體株高、節間數、葉寬、節間粗為敏感性狀,葉長、節間長為惰性性狀。羊草個體形態對放牧的響應更多反映的是羊草對由于放牧而引起的多種改變了的環境因子的響應,是一種“復合式響應”,也是草原退化后植株個體形態的一種表現形式,而土壤供磷能力只是多個環境因子中的一個因子。因此,羊草植株個體形態對土壤供磷能力及對過度放牧響應規律的差異可能是由于在天然草原生態系統中,羊草的生長受到多種生物和非生物因素的影響,植物需要通過資源整合來實現各個性狀之間的權衡,從而在各種環境因素影響下表現出一個最優的生長狀況[37-41],因此其對單一環境因素的響應與其對多個因素的響應可能會出現非常大的差異。本研究結果與其他研究中所出現的差異也反映出草原生態系統中土壤供磷能力之外的各種環境因子,包括放牧啃食與踐踏、水分、光照、氮素營養等,都可能會對植物個體形態的影響產生作用,甚至有可能存在耦合作用。必須指出的是磷元素主要對植物的發育產生影響,而氮元素主要對植物的伸長生長產生影響,這可能是羊草對過度放牧和對土壤供磷能力的形態可塑性響應規律出現差異的原因之一,因此,氮元素可能在天然草原退化過程中起著重要的作用[30]。在復雜的生態系統中分別針對植物個體形態在單一環境因子與多環境因子下進行可塑性研究,更有助于闡明植物形態變化在天然草原生態系統中的作用,進而揭示天然草原退化的內在機理。
植物在不同環境下同化物分配的差異反映了其對環境變化的響應方式及適應策略[42-43]。在草原退化過程中,植物會將更多的光合產物分配到地下,同時在地上增加對葉片等光合器官的分配,以增大捕獲光資源的能力,提高光合速率,增強植物的養分吸收能力,保證其在放牧干擾下的生存和再生[44-45]。大量研究表明土壤養分條件的改善可以增加植物干物質的產量、降低根冠比率,增加同化物向地上部的分配[46-49]。但是本研究表明隨著土壤供磷能力的增強,植物向莖葉的同化物分配減少,向根的分配增加,這與很多養分添加試驗的結果并不一致。一個可能的原因是羊草屬于根莖型多年生禾草,在天然草原其繁殖主要靠根莖的無性繁殖,在土壤供磷能力差時優先分配光合器官以提高光合能力,在土壤供磷能力強時優先分配給地下的繁殖器官,增加根莖節間數,提高其種群密度,以保證其繁殖能力,因此這種現象是羊草的一種繁殖策略。另外一種可能是這種現象是羊草對磷素養分的一種適應策略,在土壤供磷能力差時可以將更多的光合產物分配給地上以提高光合能力,而在地下通過改變根系形態、增加根系吸收面積,以最少的根部生物量獲得最大的養分吸收面積。有關羊草在土壤養分影響下的資源分配規律及其他的適應策略還有待進一步研究。
在環境梯度下,很多植物的各個性狀之間均具有協同變化的特征[50]。課題組前期研究發現放牧干擾下羊草的各個性狀之間存在明顯的協同變化趨勢,隨著羊草株高的降低,莖葉各性狀也出現了降低的趨勢,且株高與各性狀指標之間存在顯著的相關性[3]。本研究表明羊草地上生物量分別與株高、節間數、葉寬,羊草株高分別與節間數、節間長、葉寬的相關性均達到了極顯著水平,也就是說在土壤供磷能力梯度下羊草的個體大小與部分形態指標之間存在協同變化規律。偏最小二乘法分析發現葉寬、節間數、株高對羊草個體地上生物量的貢獻率分別為30%,27%,26%,此3個形態指標為導致羊草個體地上生物量在不同土壤供磷能力下發生差異的主要驅動因子。放牧干擾下驅動羊草地上生物量減少的形態指標為莖長、株高、葉長[3],因此,土壤供磷能力可能只是放牧干擾對羊草的影響途徑之一,放牧對天然草原的作用還會受到氮、水分等其他因素的影響。
4 結論
1)土壤有效磷含量的匱乏明顯減小了羊草的植株高度、各構件生物量,造成了羊草個體植株的矮小化。
2)雖然羊草在土壤磷元素匱乏下表現出植株變矮,生物量減小的矮小化現象,但是其個體形態、生物量分配對土壤供磷能力的響應與課題組前期研究的羊草對放牧干擾的響應有所不同。
3)不同土壤供磷能力條件下,羊草的葉寬、節間數、株高是驅動其個體生物量的主要因子,對羊草個體地上生物量的貢獻率分別為30%,27%,26%,這與放牧對羊草生物量的驅動因子并不一致。
[1] Mou P, Jones R H, Tan Z,etal. Morphological and physiological plasticity of plant roots when nutrients are both spatially and temporally heterogeneous. Plant and Soil, 2013, 364(1/2): 373-384.
[2] Hu Q P, Guo Z H, Li C Y,etal. Advance at Phenotypic Plasticity in plant responses to abiotic factors. Scientia Silae Sinicae, 2008, (5): 135-142.
[3] Li X L, Hou X Y, Wu X H,etal. Plastic responses of stem and leaf functional traits inLeymuschinensisto long-term grazing in a meadow steppe. Chinese Journal of Plant Ecology, 2014, (5): 440-451.
[4] Kattia Palacio-Lopez B B. The ubiquity of phenotypic plasticity in plants: A synthesis. Ecology & Evolution, 2015, 5(16):3389-3400.
[5] Xie Y H, Yu D, Geng X H. Effects of elevated CO2concentration on phenotypic physiological and biochemical characteristics of submersed plant potamogeton crispus leaf. Acta Phytoecologica Sinica, 2003, 27(2): 218-222.
[6] Sheng H Y, Li W C, Chang J. Comparison of plasticity of seedling growth of two umbellaceae species in response to light intensity. Acta Ecologica Sinica, 2006, (6): 1854-1861.
[7] Chen S Y, Zhang J L, Jia P,etal. Impact of neighborhood effects on adaptive plasticity of plant height under light competition environment. Journal of Lanzhou University: Natural Sciences, 2009, (6): 76-81.
[8] Tao Y, Zhang Y M, Quan Y W,etal. Morphology and biomass characteristics of ephemeroid plantAlliumpallasiiand their relationships in junggar basin, China. Journal of Desert Research, 2012, (5): 1328-1334.
[9] Geng Y P, Zhang W J, Li B,etal. Phenotypic plasticity and invasiveness of alien plants. Biodiversity Science, 2004, (4): 447-455.
[10] Li X H, Li X L, Jiang D M,etal. A comparative study of the individual biomass and modular biomass of 70 herbaceous species found in the Horqin Sandy Land. Arid Zone Research, 2009, 26(2): 56-62.
[11] Zhang W H, Li H, Li J X,etal. Individual and modular biomass dynamics ofKingdoniauninflorapopulation in Qinling Mountain. Chinese Journal of Applied Ecology, 2003, (4): 530-534.
[12] Zhu J D, Meng T T, Ni J,etal. Within-leaf allometric relationships of mature forests in different bioclimatic zones vary with plant functional types. Chinese Journal of Plant Ecology, 2011, (7): 687-698.
[13] Wang W, Liang C Z, Liu Z L,etal. Analysis of the plant individual behaviour during the degradation and restoring succession in steppe community. Acta Phytoecologica Sinica, 2000, 24(3): 268-274.
[14] Connell J H, Slatyer R O. Mechanisms of succession in natural communities and their role in community stability and organization. American Naturalist, 1977, 111: 1119-1144.
[15] Bai Y F, Han X G, Wu J G,etal. Ecosystem stability and compensatory effects in the Inner Mongolia grassland. Nature, 2004, 431: 181-184.
[16] Zhang Z H. Silent crisis-desertification and degeneration of grassland. Pratacultura Science, 2000, 17(2): 10-12.
[17] Mcconnaughay K, Coleman J S. Biomass allocation in plants: ontogeny or optimality? A test along three resource gradients. Ecology, 1999, 80(8): 2581-2593.
[18] Liu Z L, Wang W. The discussion about the mechanism of degradation and restoring succession in Inner Mongolia. Journal of Arid Land Resources and Environment, 2002, 16(1): 84-91.
[19] Wang W, Liang C Z, Liu Z L,etal. Mechanism of degradation succession inLeymuschinensis+Stipagrandissteppe community. Acta Phytoecologica Sinica, 2000, (4): 468-472.
[20] An H. Effect of grazing on morphological plasticity and biomass allocation of dominant species in desert steppe. Journal of Arid Land Resources and Environment, 2014, (11): 116-121.
[21] Li J H, Li Z Q. Clonal morphological plasticity and biomass allocation pattern ofArtemisiafrigidaandPotentillaacaulisunder different grazing intensities. Acta Phytoecologica Sinica, 2002, (4): 435-440.
[22] Huang H X, Zhao X Y, Zhang H X. Responses ofAgriophyllumsquarrosumphenotypic plasticity to the changes of soil nutrient and moisture contents and population density. Chinese Journal of Plant Ecology, 2008, (12): 2593-2598.
[23] An Y, Han G D. The difference between the grass and soil in different stage of grassland deterioration. Grassland and China, 1999, (4): 31-36.
[24] Clark C M, Tilman D. Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature, 2008, 451: 712-715.
[25] Yang Y, Ji C, Ma W,etal. Significant soil acidification across northern China’s grasslands during 1980s-2000s. Global Change Biology, 2012, 18(7): 2292-2300.
[26] Aber J D, Nadelhoffer K J, Steudler P,etal. Nitrogen saturation in northern forest ecosystems. Bioscience, 1989, 39(6): 286-378.
[27] Bobbink R, Hicks K, Galloway J,etal. Global assessment of nitrogen deposition effects on terrestrial plant diversity: a synthesis. Ecological Applications, 2010, 20(1): 30-59.
[28] Menge D N, Field C B. Simulated global changes alter phosphorus demand in annual grassland. Global Change Biology, 2007, 13(12): 2582-2591.
[29] Liu N N, Tian Q Y, Zhang W H. Comparison of adaptive strategies to phosphorus-deficient soil between dominant speciesArtemisiafrigidaandStipakryloviiin typical steppe of Nei Mongol. Chinese Journal of Plant Ecology, 2014, (9): 905-915.
[30] Bai X, Cheng J H, Zheng S X,etal. Ecophysiological responses ofLeymuschinensisto nitrogen and phosphorus additions in a typical steppe. Chinese Journal of Plant Ecology, 2014, 38(2): 103-115.
[31] Xu M G, Li J M. Plant Nutrition Elements of Soil Chemistry[M]. Beijing: Graduate School of Chinese Academy of Agricultural Sciences, 2005.
[32] Li X L. Effects of Irrigation and Phosphorus Fertilizer Application for Consecutive Years on Alfalfa Yield and Soil Environment[D]. Beijing: Chinese Academy of Agricultural Sciences, 2014.
[33] Valladares F, Wright S J, Lasso E,etal. Plastic phenotypic response to light of 16 congeneric shrubs from a Panamanian rainforest. Ecology, 2000, 81(7): 1925-1936.
[34] Wu N E, Hai T. The effects of grazing on the morphological characteristics ofLeymuschinensisand soil physical and chemical properties in typical grassland. Journal of Inner Mongolia Agricultural University: Natural Science Edition, 2015, (4): 71-76.
[35] Zhao W. Physio-ecological Responses ofLeymuschinensisto Overgrazing and Clipping[D]. Beijing: Chinese Academy of Agricultural Sciences, 2006.
[36] Yu Y J. The Changes in Eco-physiological and Epigenetic Features ofLeymuschinensisin Responses to Warming and Nitrogen Addition[D]. Changchun: Northeast Normal University, 2014.
[37] Li X L, Liu Z Y, Hou X Y,etal. Plant functional traits and their trade-offs in response to grazing: a review. Chinese Bulletin of Botany, 2015, 50(2): 159-170.
[38] He M, Huang J H. Influence of grazing on seed production ofCaraganamicrophylla. Chinese Bulletin of Botany, 2010, (1): 59-65.
[39] Chen H J. The Response of Reproductive Characteristics and Ecological Stoichiometry of Main Plant Population to Stocking Rate in Inner Mongolia Desert Steppe[D]. Hohhot: Inner Mongolia Agricultural University, 2011.
[40] Bai Y, Wu J, Clark C M,etal. Grazing alters ecosystem functioning and C∶N∶P stoichiometry of grasslands along a regional precipitation gradient. Journal of Applied Ecology, 2012, 49(6): 1204-1215.
[41] Frank D A. Ungulate and topographic control of nitrogen: phosphorus stoichiometry in a temperate grassland; soils, plants and mineralization rates. Oikos, 2008, 117(4): 591-601.
[42] Smith S E. Variation in response to defoliation between populations ofBoutelouacurtipendulavar.caespitosa(Poaceae) with different livestock grazing histories. American Journal of Botany, 1998, 85(9): 1266-1272.
[43] Geber M A, Watson M A, de Kroon H. Development and resource allocation in perennial plants: The significance of orgon preformation[M]//Plant Resource Allocation. New York: Academic Press, 1997.
[44] Ren H Y, Zhen S X, Bai Y F. Effects of grazing on foliage biomass allocation of grassland communities in Xi Lin River Basin, Inner Mongolia. Chinese Journal of Plant Ecology, 2009, 33(6): 1065-1074.
[45] Roy J, Winner W E, Pell E J. Response of Plants to Multiple Stresses[M]. USA: Academic Press, 2012.
[46] Weigelt A, Bol R, Bardgett R D. Preferential uptake of soil nitrogen forms by grassland plant species. Oecologia, 2005, 142(4): 627-635.
[47] Wang S P, Wang Y F, Chen Z Z,etal. Sulphur concentration of soils and plants and its requirement for ruminants in the Inner Mongolia steppe of China. Grass and Forage Science, 2001, 56(4): 418-422.
[48] Reich P B, Buschena C, Tjoelker M G,etal. Variation in growth rate and ecophysiology among 34 grassland and savanna species under contrasting N supply: a test of functional group differences. New Phytologist, 2003, 157(3): 617-631.
[49] Ping X Y, Zhou G S, Sun J S. Advances in the study of photosynthate allocation and its controls. Chinese Journal of Plant Ecology, 2010, (1): 100-111.
[50] Hodge A. The plastic plant: root responses to heterogeneous supplies of nutrients. New Phytologist, 2004, 162(1): 9-24.
[2] 胡啟鵬, 郭志華, 李春燕, 等. 植物表型可塑性對非生物環境因子的響應研究進展. 林業科學, 2008, (5): 135-142.
[3] 李西良, 侯向陽, 吳新宏, 等. 草甸草原羊草莖葉功能性狀對長期過度放牧的可塑性響應. 植物生態學報, 2014, (5): 440-451.
[5] 謝永宏, 于丹, 耿顯華. CO2濃度升高對沉水植物菹草葉表型及生理生化特征的影響. 植物生態學報, 2003, 27(2): 218-222.
[6] 盛海燕, 李偉成, 常杰. 傘形科兩種植物幼苗生長對光照強度的可塑性響應. 生態學報, 2006, (6): 1854-1861.
[7] 陳書燕, 張甲林, 賈鵬, 等. 光競爭條件下鄰域效應對植物高生長可塑性的影響. 蘭州大學學報: 自然科學版, 2009, (6): 76-81.
[8] 陶冶, 張元明, 全永威, 等. 準噶爾荒漠小山蒜的形態與生物量特征及其相互關系. 中國沙漠, 2012, (5): 1328-1334.
[9] 耿宇鵬, 張文駒, 李博, 等. 表型可塑性與外來植物的入侵能力. 生物多樣性, 2004, (4): 447-455.
[10] 李雪華, 李曉蘭, 蔣德明, 等. 科爾沁沙地70種草本植物個體和構件生物量比較研究. 干旱區研究, 2009, (2): 200-205.
[11] 張文輝, 李紅, 李景俠, 等. 秦嶺獨葉草種群個體和構件生物量動態研究. 應用生態學報, 2003, (4): 530-534.
[12] 祝介東, 孟婷婷, 倪健, 等. 不同氣候帶間成熟林植物葉性狀間異速生長關系隨功能型的變異. 植物生態學報, 2011, (7): 687-698.
[13] 王煒, 梁存柱, 劉鐘齡, 等. 草原群落退化與恢復演替中的植物個體行為分析. 植物生態學報, 2000, 24(3): 268-274.
[16] 張自和. 無聲的危機——荒漠化與草原退化. 草業科學, 2000, 17(2): 10-12.
[18] 劉鐘齡, 王煒. 內蒙古草原退化與恢復演替機理的探討. 干旱區資源與環境, 2002, 16(1): 84-91.
[19] 王煒, 梁存柱, 劉鐘齡, 等. 羊草+大針茅草原群落退化演替機理的研究. 植物生態學報, 2000, (4): 468-472.
[20] 安慧. 放牧干擾對荒漠草原優勢植物形態可塑性及生物量分配的影響. 干旱區資源與環境, 2014, (11): 116-121.
[21] 李金花, 李鎮清. 不同放牧強度下冷蒿、星毛委陵菜的形態可塑性及生物量分配格局. 植物生態學報, 2002, (4): 435-440.
[22] 黃迎新, 趙學勇, 張洪軒, 等. 沙米表型可塑性對土壤養分、水分和種群密度變化的響應. 應用生態學報, 2008, (12): 2593-2598.
[23] 安淵, 韓國棟. 不同退化梯度草地植物和土壤的差異. 中國草地, 1999, (4): 31-36.
[29] 劉娜娜, 田秋英, 張文浩. 內蒙古典型草原優勢種冷蒿和克氏針茅對土壤低磷環境適應策略的比較. 植物生態學報, 2014, (9): 905-915.
[30] 白雪, 程軍回, 鄭淑霞, 等. 典型草原建群種羊草對氮磷添加的生理生態響應. 植物生態學報, 2014, 38(2): 103-115.
[31] 徐明崗, 李菊梅. 植物營養元素的土壤化學[M]. 北京: 中國農業科學院研究生院, 2005.
[32] 李新樂. 連續多年灌水施磷肥對紫花苜蓿產量和土壤環境的影響[D]. 北京: 中國農業科學院, 2014.
[34] 烏尼爾, 海棠. 放牧對典型草原羊草形態特征及土壤理化性質的影響. 內蒙古農業大學學報: 自然科學版, 2015, (4): 71-76.
[35] 趙威. 羊草對過度放牧和刈割的生理生態響應[D]. 北京: 中國科學院研究生院(植物研究所), 2006.
[36] 于英杰. 增溫和施氮導致羊草生理生態與表觀遺傳變化[D]. 長春: 東北師范大學, 2014.
[37] 李西良, 劉志英, 侯向陽, 等. 放牧對草原植物功能性狀及其權衡關系的調控. 植物學報, 2015, 50(2): 159-170.
[38] 何茂, 黃建輝. 放牧對小葉錦雞兒種子產量的影響. 植物學報, 2010, (1): 59-65.
[39] 陳海軍. 荒漠草原主要植物種群繁殖性狀及化學計量特征對載畜率的響應[D]. 呼和浩特: 內蒙古農業大學, 2011.
[44] 任海彥, 鄭淑霞, 白永飛. 放牧對內蒙古錫林河流域草地群落植物莖葉生物量資源分配的影響. 植物生態學報, 2009, 33(6): 1065-1074.
[49] 平曉燕, 周廣勝, 孫敬松. 植物光合產物分配及其影響因子研究進展. 植物生態學報, 2010, (1): 100-111.
Plasticity of plant morphology and biomass allocation ofLeymuschinensisunder different phosphorus conditions
GUO Feng-Hui1, HOU Xiang-Yang1*, DING Yong1, LI Xi-Liang1, WU Zi-Nian1, WANG Zhen1, HU Jing1, LI Xin-Le2, MA Wen-Jing1, ZHAO Jin-Bo1
1.InstituteofGrasslandResearchofChineseAcademyofAgricultureSciences,NationalForageImprovementCenter,Hohhot010010,China; 2.ExperimentalCenterofDesertForestryofChinaAcademyofForestry,Dengkou015200,China
Changes in plant morphology and biomass allocation are the main factors in the natural grassland degradation process. Studying the responses of plant morphology and biomass allocation under different conditions could help to reveal the mechanism of grassland degradation. We conducted a pot experiment and evaluated the morphological responses and biomass accumulation and allocation ofLeymuschinensisunder different phosphorus (P) conditions. The treatments consisted of 0, 30, 90, and 180 mg P2O5/kg added to soil to provide four different P-availability conditions for cultivation ofL.chinensis. The results showed that: 1) plant biomass and height were reduced under low-P availability, and were significantly increased under higher-P availability. Plant height, leaf biomass, stem biomass, root biomass, and total biomass were 14%, 50%, 51%, 79%, and 63% lower, respectively, under low-P availability than under high-P availability. 2) Plant morphology and biomass allocation showed different responses to P availability. Biomass accumulation was the most sensitive to P conditions, and morphology was the least sensitive; the plasticity indexes were 0.58, 0.31, and 0.19, respectively, for biomass accumulation, allocation, and morphology. The most sensitive single trait indicator was root dry mass with the plasticity index of 0.79. 3) The correlations between aboveground dry mass and leaf width, internode number, and plant height were all significant. Understanding the responses ofL.chinensisto different P conditions can help to reveal the mechanisms of vegetation succession and degradation in the grassland ecosystem.
Leymuschinensis; P conditions of soil; phenotypic plasticity; plasticity index
10.11686/cyxb2016061
http://cyxb.lzu.edu.cn
2016-03-01;改回日期:2016-04-12
國家重點基礎研究發展計劃(973計劃)(2014CB138806),“十二五”國家科技支撐計劃(2012BAD12B02),內蒙古自治區自然科學基金重大項目(2015ZD02),中央級科研院所基本科研業務費項目(1610332015001)和國家自然基金(31502008)資助。
郭豐輝(1987-),男,山東濱州人,碩士。E-mail:guofhui@163.com*通信作者Corresponding author. E-mail: houxy16@126.com
郭豐輝, 侯向陽, 丁勇, 李西良, 武自念, 王珍, 胡靜, 李新樂, 馬文靜, 趙勁博. 羊草形態及生物量分配可塑性對土壤供磷能力的響應研究. 草業學報, 2016, 25(12): 53-62.
GUO Feng-Hui, HOU Xiang-Yang, DING Yong, LI Xi-Liang, WU Zi-Nian, WANG Zhen, HU Jing, LI Xin-Le, MA Wen-Jing, ZHAO Jin-Bo. Plasticity of plant morphology and biomass allocation ofLeymuschinensisunder different phosphorus conditions. Acta Prataculturae Sinica, 2016, 25(12): 53-62.
猜你喜歡
發明與創新(2022年30期)2022-10-03 08:40:56
動漫星空(興趣百科)(2020年12期)2020-12-12 05:31:40
人大建設(2018年6期)2018-08-16 07:23:10
文理導航·科普童話(2017年5期)2018-02-10 19:42:14
無人機(2017年10期)2017-07-06 03:04:36
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
