塔里木河下游胡楊生物量及其空間分布特征
2017-01-04 09:36:46王希義徐海量潘存德凌紅波苑塏燁
西北植物學報 2016年11期
王希義,徐海量,潘存德,凌紅波,苑塏燁
(1 新疆農業大學,烏魯木齊 830052;2 中國科學院新疆生態與地理研究所,烏魯木齊 830011)
塔里木河下游胡楊生物量及其空間分布特征
王希義1,徐海量2*,潘存德1,凌紅波2,苑塏燁1
(1 新疆農業大學,烏魯木齊 830052;2 中國科學院新疆生態與地理研究所,烏魯木齊 830011)
該研究采用野外樣地調查法,以塔里木河下游地區的胡楊為研究對象,以胡楊地上生物量與易測因子(胸徑、樹高)之間的關系模型為基礎,依據監測樣帶胡楊狀況,對研究區內胡楊生物量分布狀況進行分析,并結合區域內胡楊林面積對胡楊總生物量進行了評估。結果表明:(1)胡楊生物量模型B=128.09(D2h)0.6318(R2=0.892),精度較高,并且驗證的實測值與模擬值之間差異不顯著(P> 0.05),此模型可以用來對胡楊生物量進行評估。(2)隨著離河道距離的增加,胡楊生物量呈減少趨勢;胡楊生物量主要分布在離河道700 m以內的范圍,占總生物量的91.37%。(3)塔里木河下游胡楊單位面積生物量為416.613 kg·hm-2,總生物量為2.474×104t。
胡楊;地上生物量;空間分布;塔里木河下游
森林是陸地生態系統的主體[1],與其他生態系統相比具有更為復雜的層次結構和更長的生命周期,是陸地生物光合產量的主體[2-3]。森林生物量指個體、種群或群落生物質數量的多少,是生態系統特征的重要指標之一,也是森林系統環境質量的綜合體現[4]。生物量不僅反映了生態系統生產力水平的高低,也是植被固碳能力的重要標志和評估植被碳收支的重要參數[5-6],其測定在碳循環及氣候變化等研究中具有重要意義[7-8]。
自國際生物學計劃(IBP)實施以來,生物量一直是生態學研究的重點[9-10]。在國外,一些學者針對森林生物量與生產力進行了研究,建立了生物量與碳循環之間的關系[11-13],并對不同區域的生物量進行了評估[14-15];國內針對熱帶雨林、亞熱帶常綠闊葉林、溫帶針葉林等森林系統的生物量進行了探討,分析了森林生物量的分布規律與變化特征[16-19]。然而這些研究大都集中于東部濕潤區或是熱帶亞熱帶地區,有關干旱區森林生物量的研究較為少見。
胡楊是塔里木河下游的植被建群種,對于區域內環境保護和植被恢復程度具有最直接的指示作用[20]。針對塔里木河下游胡楊的研究主要集中于胡楊與水資源的關系[21-22]、胡楊生態分布[23-25]與胡楊生理特征[26-28]等方面,而有關生物量模擬評估的研究較少。本研究通過查詢生物量模擬方法,結合調查資料,探討了塔里木河下游胡楊生物量的空間分布特征,并對胡楊總生物量進行了評估,以便于人們對胡楊在生活與生產中的作用有一個恰當認識,為塔里木河下游植被保護和恢復以及準確估算區域胡楊生物量和碳儲量提供科學依據。
1 研究區概況
塔里木河下游是指從大西海子水庫至臺特瑪湖河段(圖1)。本河段位于庫魯克沙漠與塔克拉瑪干沙漠之間,區域內的荒漠河岸林形成了南北長約400 km的綠色走廊。塔里木河下游屬溫帶干旱荒漠氣候,年降水量僅為20~50 mm,而潛在蒸發量卻高達2 500~3 000 mm[29]。
由于人類不合理的水土資源開發和利用,使下游的河道曾經長期斷流,生態嚴重退化,植被長勢極度衰敗,區域內建群植物——胡楊基本為過熟林,中幼齡林基本枯死[30]。基于此,中國政府于2000年5月組織實施了以生態恢復和環境保護為目標的塔里木河下游生態輸水工程。在實施生態輸水的過程中,地下水位不斷抬升,地下水的響應寬度不斷擴大[31];另外,河道兩岸出現了大范圍河水漫溢現象,激活了土壤種子庫,在漫溢區萌生了大量幼苗[32],增加了生態系統的穩定性,在一定程度上緩解了區域內的生態退化過程。塔里木河下游喬木主要是胡楊(Populuseuphratica),灌木主要包括檉柳(Tamarixchinensis)、黑刺(Lyciumruthenicum)、鈴鐺刺(Halimodendronhalodendron)等,草本植物包括蘆葦(Phragmitescommunis)、鹿角草(GlossogynetenuifoliaCass)、駱駝刺(Alhagisparsifolia)、花花柴(Kareliniacaspica)、鹽生草(Halogetonglomeratus)等。
2 資料來源與研究方法
2.1 資料來源
于2016年6月上中旬至7月上旬在塔里木河下游進行胡楊樣地調查。選取塔里木河下游英蘇、喀爾達依、阿拉干、依干不及麻4個斷面。對于每一個監測斷面,選擇具有代表性、未受人類破壞的地段,沿垂直于河道方向設置一個長2 000 m、寬100 m的樣帶,然后將此樣帶平均分成20個100 m×100 m的大樣方進行監測。首先統計各個大樣方內胡楊的數量,然后對每個大樣方內的胡楊進行每木檢尺(DBH≥4 cm),記錄其胸徑、樹高、冠幅等,對于DBH<4 cm的幼苗及幼樹,記錄其數量和高度。
由于以前沒有開展胡楊生物量的研究,因此在2016年野外調查時,為了保證實驗的合理性與數據的均勻性,并兼顧保護植被稀疏地帶的胡楊不再遭受破壞,本研究在胡楊分布最多的英蘇和阿拉干斷面對胡楊進行取樣。在對胡楊的取樣過程中,首先選取胡楊分布密集且均勻的地區,一共標記60株幼中齡的、長勢良好的標準木,先記錄這些胡楊的胸徑、樹高、冠幅等,然后在地面根基部位鋸斷伐倒并稱重,記錄這60株胡楊的地上生物量。
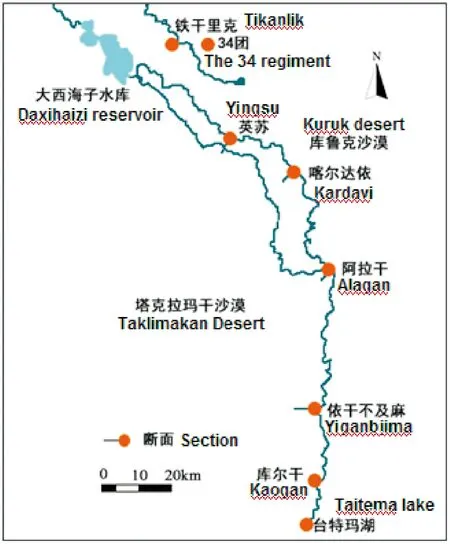
圖1 塔里木河下游各監測斷面狀況Fig.1 Distribution of monitoring sections in the lower reaches of Tarim River
2.2 研究方法
本研究的關鍵方法是構建胡楊生物量估算模型。本文采用唐守正[33]建立的生物量回歸模型計算胡楊單株生物量。
B=a(D2h)b(D為胸徑,h為樹高)
(1)
首先用30株伐倒胡楊(編號1,2,……,30)的相關數據對a與b進行參數擬合,計算出另外30株(編號31,32,……,60)胡楊生物量,并用T檢驗方法,通過計算P值分析模擬值與實測值之間差異的顯著性,若P< 0.05,則模擬值與實測值差異顯著;若P> 0.05,則模擬值與實測值之間不顯著。然后計算樣方內胡楊總生物量,進而推算胡楊單位面積生物量,最后結合胡楊林總面積計算總生物量。
本研究分析和作圖軟件用SPSS16.0與EXCEL。
3 結果與分析
3.1 胡楊生物量模型建立與變量統計
3.1.1 胡楊生物量模型構建 利用伐倒30株胡楊建立回歸模型(方程(2)),且擬合度達到顯著水平。
B= 128.09(D2h)0.6318(R2= 0.892;F= 285.35)
(2)
之后利用剩余30株胡楊特征數據進行驗證,對實測值與模擬值進行對比分析(圖2)。由圖2可知,實測值與模擬值之間沒有顯著差異(P> 0.05),擬合方程合理。因此可以用方程(2)計算胡楊的生物量。
3.1.2 胡楊地上變量統計分析 結合胡楊生物量估算模型(1),重點對所調查胡楊的胸徑和樹高進行統計分析(表1)。由表1可知,胡楊胸徑的最小值和最大值都出現在英蘇斷面。由于胡楊的胸徑反映出了樹齡的大小[34],因此英蘇斷面的胡楊樹齡寬度最大。依干不及麻斷面胡楊胸徑平均值最低,高度平均值也顯著低于其他3個斷面,這導致胡楊單株平均生物量顯著低于其他斷面。另外,胡楊單株生物量最小值和最大值均出現在英蘇斷面,表明在英蘇斷面胡楊齡級跨度大;依干不及麻斷面胡楊生物量最小值與最大值差異最小,表明該斷面胡楊齡級跨度小。
3.2 胡楊地上生物量空間分布特征
3.2.1 各斷面胡楊單位面積生物量橫向分布特征 生物量橫向分布特征即為隨著離河距離的增加,生物量的變化趨勢。本文將4條樣帶內不同離河距離胡楊生物量特征作圖討論(圖3)。
由圖3可知,在離河道700 m以內胡楊分布較多,生物量總量較大,占總生物量的91.37%;700 m以外生物量分布較少。這主要是由于隨著離河距離的增加,河水漫溢作用減弱,地下水補給量逐漸減少,水位降低,逐漸鄰近甚至低于胡楊生長的脅迫水位,導致胡楊生長萎靡,生物量降低。對于各個斷面而言,近河岸胡楊生物量高于離河較遠的地區,最為明顯的是英蘇斷面,其次是依干不及麻斷面。喀爾達依斷面的胡楊生物量在離河900 m附近出現一個小高峰,阿拉干斷面在離河600~700 m處胡楊的生物量也相對較高,造成這種現象的原因也許與微地形或是樣帶的隨機設置有關,具體原因需要進一步探討。
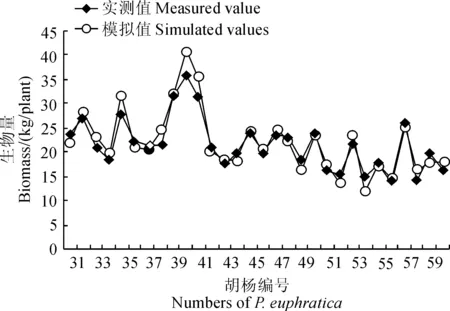
圖2 胡楊實測值與模擬值對比Fig.2 Comparison of measured values and simulation values of P. euphratica
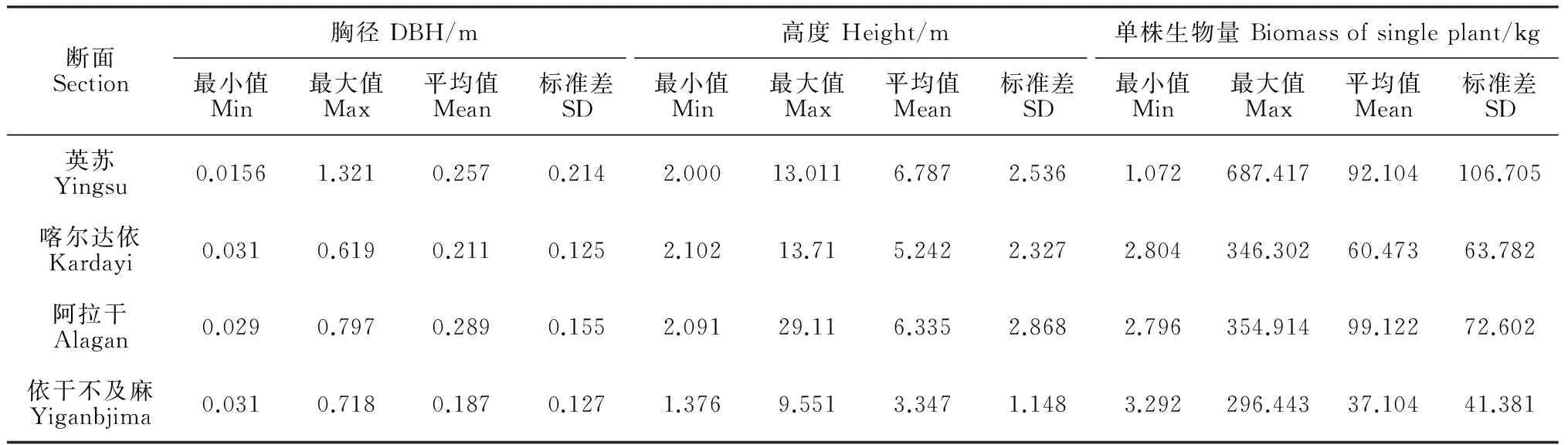
表1 胡楊地上因子統計
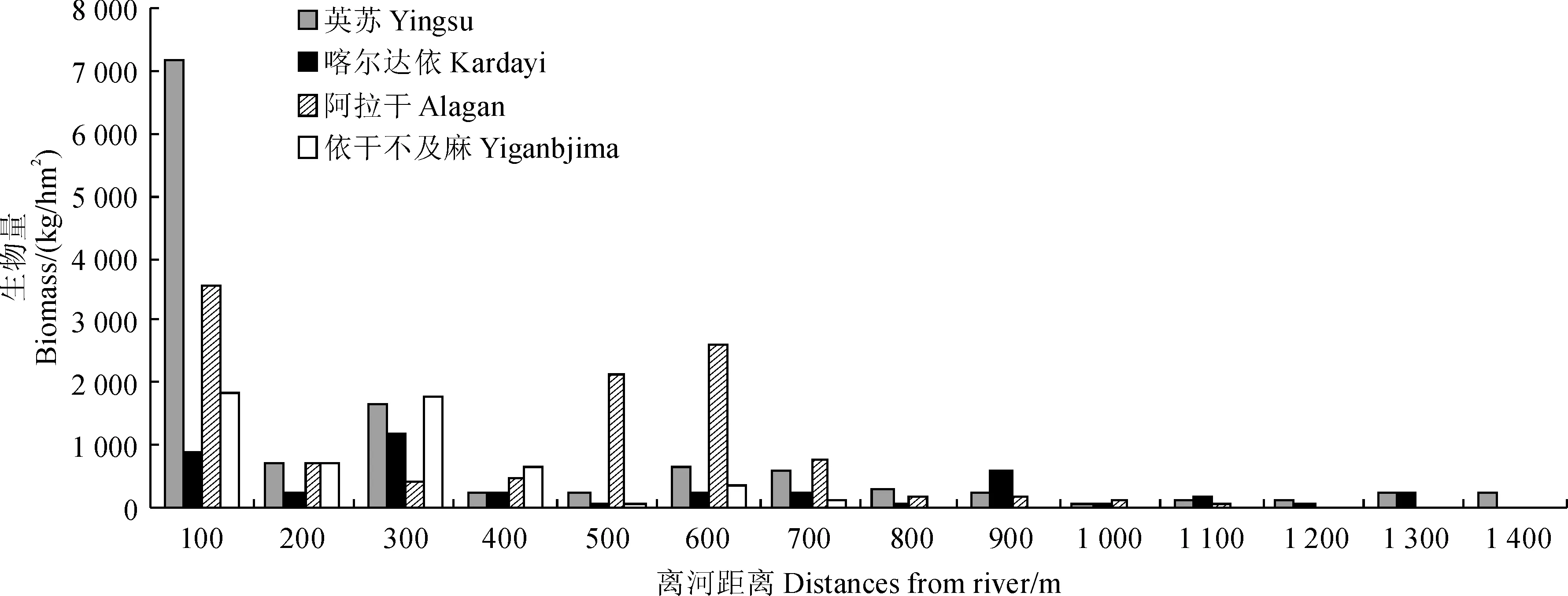
圖3 各斷面不同離河距離上胡楊單位面積生物量分布特征Fig.3 The distribution characteristics of P. euphratica biomass per unit area in different distances from river to each section
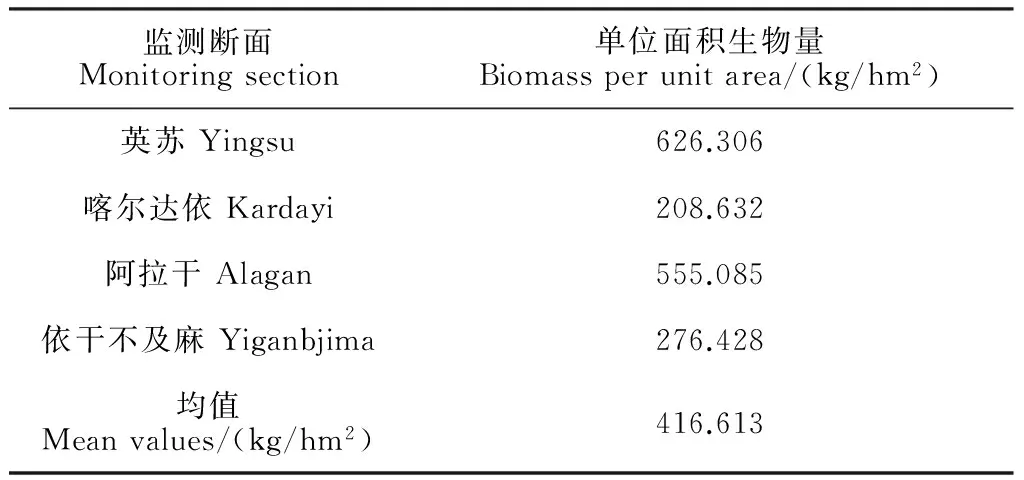
表2 各監測樣帶內胡楊單位面積生物量
3.2.2 胡楊生物量縱向分布特征 生物量縱向分布規律即在沿河道方向上從上游到下游的變化特征。本文將4條樣帶的胡楊總生物量進行了對比分析(圖4)。
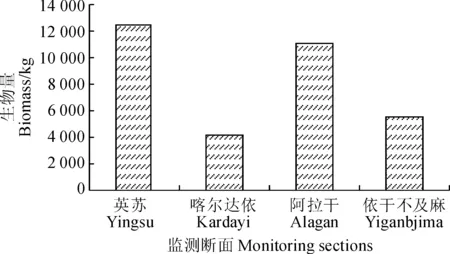
圖4 自上游到下游胡楊生物量特征Fig.4 The distribution characteristics of P. euphratica biomass from upstream to downstream
各樣帶中,英蘇的胡楊生物量最高,阿拉干次之,喀爾達依和依干不及麻的胡楊生物量最小。因為英蘇離放水源最近,斷面內河水對地下水和土壤水的轉化量較大,可以較好地滿足胡楊的生長需求,因此胡楊的數量較多,長勢較好;另外,阿拉干斷面位于其文闊爾河與老塔里木河的交匯處,這使得阿拉干斷面水源也很充足,胡楊生物量也較高;在喀爾達依與依干不及麻斷面,河道的過水量較少,導致河水對地下水和土壤水的轉化量較低,斷面內的水分條件限制了胡楊的生長,使得胡楊的數量少,長勢差,生物量較低。
3.3 塔里木河下游胡楊生物量估測
3.3.1 胡楊單位面積生物量 由于樣帶設置的隨機性,樣方內所取胡楊特征也具有隨機性。在本研究中,每個樣帶設置了20個大樣方,但并非所有的樣方內都有胡楊。例如阿拉干斷面,在樣帶內,離河道1 100 m以內有胡楊,遠處沒有胡楊;在樣帶之外離河道1 100 m以外依然有胡楊分布,直至到2 000 m以外的區域都有胡楊。因此本研究中,計算樣帶內胡楊平均生物量(表2)依然使用整條樣帶的面積,而非僅有胡楊分布區域的面積。
由表2可知,英蘇斷面胡楊單位面積生物量最高,阿拉干次之,喀爾達依斷面最小,這與圖4相對應。另外,塔里木河下游胡楊單位面積平均生物量取4個斷面單位面積生物量的平均值,即416.613 kg/hm2。
3.3.2 區域內胡楊總生物量 本研究利用計算得出的胡楊單位面積生物量,然后結合塔里木河下游胡楊林總面積,可得塔里木河下游胡楊總生物量。在本研究中,根據2013年的調查結果,塔里木河下游林地總面積約為593.8 km2[35],即59 380 hm2,得胡楊總生物量為2.474×107kg(416.613 kg/hm2×59 380 hm2)。
4 討 論
喬木生物量估測是森林生物量評估中重要組成部分,其模型中變量的選擇有多種形式,其中最常用的是以胸徑和高度作為模型的變量;另外董道瑞[36]建立了冠幅與生物量之間的關系模型,曾慧卿[37]以冠幅和高度之積為自變量擬合了喬木生物量估測模型,精度均較高,都在0.8左右。本研究中,以胸徑平方與樹高之積作為變量,與胡楊的地上生物量建立了關系模型,精度高于冠幅與生物量間關系模型,并且利用此模型計算的估測值接近實測值,這與Lyon[38]揭示的體積與生物量的關系比面積與生物量關系更為緊密的結論相一致。在實施生態輸水以前,塔里木河下游各個斷面胡楊都十分衰敗,自開展生態輸水以來,胡楊大面積復蘇[39]。英蘇斷面離放水地點較近,地下水位較高,并且河水漫溢作用較強,大大促進了各齡級胡楊的生長發育,胡楊平均生物量也最高。另外,塔里木河下游生態輸水是其文闊爾河和老塔里木河雙河道輸水,兩個河道在阿拉干斷面匯合,使得阿拉干斷面水量也較多,胡楊長勢較好,平均生物量也較高,這與本研究成果相吻合。依干不及麻斷面胡楊齡級跨度小,長勢較差,這是由于依干不及麻斷面離水源較遠,地下水位低,漫溢作用弱,抑制了胡楊的生長發育[40]。
植被的生長發育與生態環境之間存在著一定的關系,生態環境的變化影響著植被的組成、結構、形成以及動態分布等,而植被的變化又會反過來作用于生態環境[41]。在干旱區,植被地上生物量與地下水間的關系十分顯著[42-43]。在離河道近處,地下水埋深較淺,植物根系與地下水的接觸面積相對較大,水分較為充足,漫溢作用較強,植被長勢較好,單位面積生物量較大;隨著與河距離的增加,地下水埋深加大,植物根系與地下水的接觸面積越來越小,水分條件逐漸變差,植被長勢逐漸衰弱,單位面積生物量減小,這正與本研究的結果相符合。本研究所布設的樣帶內,胡楊大都分布在離河道1 000 m的范圍內,1 000 m以外胡楊分布很少。在離河道700 m以內胡楊分布較多,生物量總量較大,700 m以外生物量分布較少。另外,具體到每一個監測樣帶,胡楊生物量分布沒有明顯規律,如喀爾達依斷面胡楊生物量在離河900 m出現一個小高峰,阿拉干斷面在離河600~700 m處胡楊的生物量也較高,這也許與胡楊在荒漠環境中呈斑塊狀分布[44]有關,或是受到微地形或樣帶隨機設置的影響,具體原因需要進一步探討。
塔里木河下游荒漠河岸林衰敗、老化與退化現象嚴重[45],胡楊平均生物量僅為416.613 kg·hm-2,遠遠低于中國東部地區,如貢嘎山區[46]森林單位面積生物量最高達568 t·hm-2,是塔里木河下游的十幾倍;又如浙江林分單位面積生物量雖遠低于全國平均水平[47],但是依然高于塔里木河下游。這一方面說明塔里木河下游胡楊生物量與生產力都很低,另一個方面也反映了胡楊恢復還有很大發展潛力。但是,在所設立的樣帶內,不僅僅分布有胡楊,還有檉柳、駱駝刺、花花柴、鹿角草等灌木和草本植物,胡楊的生長發育與這些林下植被有密切關系,如胡楊根系提水作用促進灌草的生長[48],而灌草的物種多樣性維持了生態系統的穩定[49]等,以后應加強此方面的研究。另外,就本研究的取樣方式而言,也存在著一些誤差和不確定性,如取樣選取的是幼中齡胡楊并建立模型,用此模型模擬老齡胡楊生物量時就存在誤差。本研究采用每木檢尺、實地采伐測量的方式,從理論上來講,對于所取的胡楊樣品實測的各項數據應該沒有誤差。但是在實際的操作過程中,由于受到人為、儀器等因素的影響,計算出的結果也存在著一定誤差。但是與機械角規樣地實測法、樣圓實測法、角規典型選樣調查法等方法相比,精度是最高的[50],結果可信。因此,在以后的研究中應深入分析模型誤差的范圍、來源及影響因素,提高建模過程中各個環節的精確度。
綜上所述,本研究構建了胡楊生物量估測模型:B= 128.09(D2h)0.6318(R2= 0.892;F= 285.35),且實測值與模擬值之間沒有顯著差異(P> 0.05);英蘇斷面胡楊樹齡跨度大,各齡級胡楊分布多,依干不及麻斷面胡楊樹齡跨度小。胡楊大都分布在離河道1 000 m的范圍內,1 000 m以外胡楊分布很少;在離河道700 m以內胡楊生物量占總生物量的91.37%;700 m以外生物量較少。根據調查結果,塔里木河下游胡楊單位面積生物量約為416.613 kg·hm2,胡楊總生物量約為2.474×104t。
[1] LYON L J. Estimating twig production of serviceberry from crown volumes[J].JournalofWildlifeManagement, 1968,32(1):115-119.
[2] CONSTANZA R, RUDOLFDE G. The value of the world ecosystem service and natural capital [J].Nature, 1997,387:253-260.
[3] NELSON R, KRABILL W, TONELLI J. Estimating forest biomass and volume using airbone laser data[J].RemoteSensingofEnvironment, 1988, 24(88):247-267.
[4] 張遠彬.CO2濃度升高對紅樺幼苗生理與生長的影響[D].北京:中國科學院大學,2007.
[5] 奧得姆.生態學基礎[M].北京:人民教育出版社,1971.
[6] 王 昆.基于Quickbird和極化雷達數據的森林生物量估測方法研究[D].北京:北京林業大學,2013.
[7] 楊永恬.基于多源遙感數據的森林蓄積量估測方法研究[D].北京:中國林業科學研究院,2010.
[8] 韋龍斌.九萬山自然保護區森林生物量及碳貯量研究[D].長沙:中南林業科技大學,2015.
[9] 翟曉江.基于遙感的陜北黃龍山森林生物量模型[D].陜西楊陵:西北農林科技大學,2013.
[10] KARL M G, HEITSCHMIDT R K, HAFERKAMP M R. Vegetation biomass dynamics and patterns of sexual reproduction in a Northern Mixed-Grass Prairie[J].AmericanMidlandNaturalist, 2013, 141(Apr 1999):227-237.
[11] MUNEEPEERAKUL R, RINALDO A, RODRIGUEZ I I. Effects of river flow scaling properties on riparian width and vegetation biomass[J].WaterResourcesResearch, 2007, 43(12):55-60.
[12] RYAN J G, FYFE C T, MCALPINE C A. Biomass retention and carbon stocks in integrated vegetation bands: a case study of mixed-age brigalow-eucalypt woodland in southern Queensland, Australia[J].RangelandJournal, 2015, 37(3):261-271.
[13] SIMPSON L G, BOTKIN D B. Vegetation, the Global Carbon Cycle, and Global Measures[C].1992:413-425.
[14] ADAMS J M, PIOVESAN G. Uncertainties in the role of land vegetation in the carbon cycle[J].Chemosphere, 2002, 49(8):805-819.
[15] SINGH T P, DAS S. Predictive analysis for vegetation biomass assessment in Western Ghat Region (WG) using geospatial techniques[J].JournaloftheIndianSocietyofRemoteSensing, 2014, 42(3):549-557.
[16] ISABIRYE M, VERBIST B, MAGUNDA M K,etal. Tree density and biomass assessment in agricultural systems around Lake Victoria, Uganda[J].AfricanJournalofEcology, 2008, 46(Supplement s1):59-65.
[17] 楊存建,劉紀遠,黃 河,等.熱帶森林植被生物量與遙感地學數據之間的相關性分析[J].地理研究,2005,24(3):473-479.
YANG C J, LIU J Y, HUANG H,etal. Correlation analysis of the biomass of the tropical forest vegetation, meteorological data and topographical data[J].GeographicalResearch, 2005,24(3):473-479.
[18] 邢艷秋,王立海.基于森林調查數據的長白山天然林森林生物量相容性模型[J].應用生態學報,2007,18(1):1-8.
XING Y Q, WANG L H. Compatible biomass estimation models of natural forests in Changbai Mountains based on forest inventory[J].ChineseJournalofAppliedEcology, 2007,18(1):1-8.
[19] 范文義,張海玉,于 穎,等.三種森林生物量估測模型的比較分析[J].植物生態學報,2011,35(4):402-410.
FAN W Y, ZHANG H Y, YU Y,etal. Comparison of three models of forest biomass estimation[J].ChineseJournalofPlantEcology, 2011,35(4):402-410.
[20] 徐 婷,曹 林,申 鑫,等.基于機載激光雷達與Landsat 8 OLI數據的亞熱帶森林生物量估算[J].植物生態學報,2015,39(4):309-321.
XU T, CAO L, SHEN X,etal. Estimates of subtropical forest biomass based on airborne LiDAR and Landsat 8 OLI data[J].ChineseJournalofPlantEcology, 2015,39(4):309-321.
[21] 徐海量,宋郁東,王 強.胡楊生理指標對塔里木河下游生態輸水的響應[J].環境科學研究,2003,16(4):24-27.
XU H L, SONG Y D, WANG Q. Response of Physiological Indexes ofPopuluseuphraticato Ecological Water Transport in the Lower Reaches of the Tarim River[J].ResearchofEnvironmentalSciences, 2003,16(4):24-27.
[22] 鄧潮洲,張希明,李 利,等.河道輸水對塔里木河下游胡楊生長狀況的影響[J].中國沙漠,2010,30(2):312-318.
DENG C Z, ZHANG X M, LI L,etal. Effects of replenishing water into river on the growth ofPopuluseuphraticain Lower Reaches of Tarim River[J].JournalofDesertResearch,2010,30(2):312-318.
[23] 涂文霞,葉 茂,徐海量,等.塔里木河不同河段胡楊徑向生長及徑流的影響差異[J].干旱區研究,2014,31(3):508-515.
TU W X, YE M, XU H L,etal. Radial growth ofPopuluseuphraticaand the effect of runoff in the Tarim River Basin[J].AridZoneResearch, 2014,31(3):508-515.
[24] 蔡富艷,玉米提·哈力克,艾爾肯·艾白不拉,等.塔里木河下游阿拉干斷面胡楊樹高的分布和密度[J].生態環境學報,2008,17(3):1 086-1 090.
CAI F Y, UMUT H, ARKIN A,etal. Tree height distribution and density ofPopuluseuphraticaforest along the Argan section in the lower reaches of Tarim River[J].EcologyandEnvironment, 2008,17(3):1 086-1 090.
[25] 陳亞寧,陳亞鵬,李衛紅,等.塔里木河下游胡楊脯氨酸累積對地下水位變化的響應[J].科學通報,2003,48(9):958-961.
CHEN Y N, CHEN Y P, LI W H,etal. Response of proline accumulation ofPopuluseuphraticato the changes of groundwater level in the Lower Reaches of Tarim River[J].ChineseScienceBulletin, 2003,48(9):958-961.
[26] 張繪芳,李 霞.塔里木河下游胡楊種群空間分布格局分析[J].西北植物學報,2006,26(10):2 125-2 130.
ZHANG H F, LI X. Spatial distribution pattern ofPopuluseuphraticapopulations in the Lower Reaches of Tarim River[J].ActaBotanicaBoreali-OccidentaliaSinica, 2006,26(10):2 125-2 130.
[27] 安紅燕,葉 茂,徐海量,等.塔里木河下游胡楊徑向生長量對生態輸水的響應[J].中國沙漠,2011,31(4):957-962.
AN H Y, YE M, XU H L,etal. Response of radial increment ofPopuluseuphraticato ecological water conveyance in Lower Reaches of the Tarim River[J].JournalofDesertResearch, 2011,31(4):957-962.
[28] 朱緒超,袁國富,邵明安,等.塔里木河下游河岸帶植被的空間結構特征[J].植物生態學報,2015,39(11):1 053-1 061.
ZHU X C, YUAN G F, SHAO M A,etal. Spatial pattern of riparian vegetation in desert of the lower Tarim River basin[J].ChineseJournalofPlantEcology, 2015,39(11):1 053-1 061.
[29] 蘇里坦,李玉生,艾爾肯·艾比布拉,等.塔里木河下游河岸胡楊(Populuseuphratica)林耗水過程模擬[J].中國沙漠,2015,35(2):380-384.
SULITAN, LI Y S, AERKEN A,etal. Simulation of water consumption ofPopulouseuphraticaforests in the Lower Reaches of the Tarim River[J].JournalofDesertResearch, 2015,35(2):380-384.
[30] 張青青,徐海量,安紅燕,等.塔里木河下游河水漫溢后胡楊幼苗分布格局初探[J].西北植物學報,2011,31(11):2 298-2 303.
ZHANG Q Q, XU H L, AN H Y,etal. Distribution patterns ofPopuluseuphraticaseedlings after river overflowing in the Lower Reaches of Tarim River[J].ActaBotanicaBoreali-OccidentaliaSinica, 2011,31(11):2 298-2 303.
[31] 楊鵬年,鄧銘江,李 霞,等.塔里木河下游間歇輸水下地下水響應寬度——以塔里木河下游英蘇斷面為例[J].干旱區研究,2008,25(3):331-335.
YANG P N, DENG M J, LI X,etal. Respond width of groundwater level after conveying stream water to the Lower Reaches of the Tarim River, Xinjiang——A case study along the Yengisu Section in the Lower Reaches of Tarim River[J].AridZoneResearch, 2008,25(3):331-335.
[32] 徐海量,宋郁東,王 強,等.塔里木河中下游地區不同地下水位對植被的影響[J].植物生態學報,2004,28(3):400-405.
XU H L, SONG Y D, WANG Q,etal. The effect of groundwater level on vegetation in the Middle and Lower Reaches of The Tarim River, Xinjiang, China[J].ChineseJournalofPlantEcology, 2004,28(3):400-405.
[33] 趙振勇,王讓會,孫洪波,等.塔里木河下游輸水廊道植被恢復的生態學評價[J].科學通報,2006,51(S1):31-35.
ZHAO Z Y, WANG R H, SUN H B,etal. Ecological evaluation of vegetation restoration in water delivery corridor in The Lower Reaches of Tarim River[J].ChineseScienceBulletin, 2006,51(S1):31-35.
[34] 唐守正,張會儒,胥 輝.相容性生物量模型的建立及其估計方法研究[J].林業科學,2000,36(S1):19-27.
TANG S Z, ZHANG H R, XU H. Study on establish and estimate method of Compatible Biomass Model[J].ScientiaSilvaeSinicae, 2000,36(S1):19-27.
[35] 顧亞亞,張世卿,李先勇,等.瀕危物種胡楊胸徑與樹齡關系研究[J].塔里木大學學報,2013,25(2):66-69.
GU Y Y, ZHANG S Q, LI X Y,etal. Relationship between diameter at breast height and age of endangered speciesPopulusEuphraticaOliv[J].JournalofTarimUniversity, 2013,25(2):66-69.
[36] 白 元,徐海量,劉新華,等.塔里木河干流景觀格局梯度分析[J].干旱區研究,2013,30(6):1 064-1 072.
BAI Y, XU H L, LIU X H,etal. Gradient change of landscape pattern in the Mainstream Area of the Tarim River[J].AridZoneResearch, 2013,30(6):1 064-1 072.
[37] 董道瑞,李 霞,萬紅梅,等.塔里木河下游檉柳灌叢地上生物量估測[J].西北植物學報,2012,32(2):384-390.
DONG D R, LI X, WAN H M,etal. Aboveground biomass estimation ofTamarixramosissimaShrub in the Lower Reaches of Tarim River[J].ActaBotanicaBoreali-OccidentaliaSinica, 2012,32(2):384-390.
[38] 趙萬羽,陳亞寧,周洪華,等.塔里木河下游生態輸水后衰敗胡楊林更新能力與條件分析[J].中國沙漠,2009,29(1):108-113.
ZHAO W Y, CHEN Y N, ZHOU H H,etal. Reproductive ability and relative environment factors of degradedPopuluseuphraticaforest in ecological water delivery project at Lower Reaches of Tarim River[J].JournalofDesertResearch, 2009,29(1):108-113.
[39] 曾慧卿,劉琪璟,馬澤清,等.基于冠幅及植株高度的檵木生物量回歸模型[J].南京林業大學學報(自然科學版),2006,30(4):101-104.
ZENG H Q, LIU Q J, MA Z Q,etal. The Regression Model ofLoropetalumchinensebiomass based on canopy diameter and plant height[J].JournalofNanjingForestryUniversity(Natural Sciences Edition), 2006,30(4):101-104.
[40] 葉 茂,徐海量,龔君君,等.不同胸徑胡楊徑向生長的合理生態水位研究[J].地理科學,2011,31(2):172-177.
YE M, XU H L, GONG J J,etal. Rational ecological groundwater level ofPopuluseuphraticawith different diameter in Lower Reaches of Tarim River[J].ScientiaGeographicaSinica, 2011,31(2):172-177.
[41] 田 雨.森林群落的水分自營作用及其定量模式研究[D].北京:中國科學院研究生院,2012.
[42] 趙文智,常學禮,李啟森,等.荒漠綠洲區蘆葦種群構件生物量與地下水埋深關系[J].生態學報,2003,23(6):1 138-1 146.
ZHAO W Z, CHANG X L, LI Q S,etal. Relationship between structural component biomass ofreedpopulation and ground water depth in desert oasis[J].ActaEcologicaSinica, 2003,23(6):1 138-1 146.
[43] 王希義,徐海量,潘存德,等.塔里木河下游地下水埋深對草本植物地上特征的影響[J].生態學雜志,2015,34(11):3 057-3 064.
WANG X Y, XU H L, PAN C D,etal. The influence of groundwater depth on aboveground herbaceous characteristics in the lower reaches of Tarim River[J].ChineseJournalofEcology, 2015,34(11):3 057-3 064.
[44] 白 元.漫溢干擾對胡楊種群空間分布格局的影響[D].北京:中國科學院大學,2013.
[45] 徐海量,李吉玫,張占江,等.塔里木河下游退化荒漠河岸林地上植被與土壤種子庫關系初探[J].中國沙漠,2008,28(4):657-664.
XU H L, LI J M, ZHANG Z J,etal. Study on relation between soil seed bank and standing vegetation along different degradation levels of desert riverside forest in Lower Reaches of Tarim River[J].JournalofDesertResearch, 2008,28(4):657-664.
[46] 羅 輯,楊 忠.貢嘎山森林生物量和生產力的研究[J].植物生態學報,2000,24(2):191-196.
LUO J, YANG Z. A study on the biomass and production of forest on the Gongga Mountain[J].ChineseJournalofPlantEcology, 2000,24(2):191-196.
[47] 張茂震,王廣興,劉安興.基于森林資源連續清查資料估算的浙江省森林生物量及生產力[J]. 林業科學,2009,45(9):13-17.
ZHANG M Z, WANG G X, LIU A X. Estimation of forest biomass and net primary production for Zhejiang Province based on continuous forest resources inventory[J].ScientiaSilvaeSinicae, 2009,45(9):13-17.
[48] 蘇 芮.胡楊的水力提升作用及功能研究[D].北京:中國科學院研究生院,2011.
[49] 江小雷,岳 靜,張衛國,等.生物多樣性,生態系統功能與時空尺度[J].草業學報,2010,19(1):219-225.
JIANG X L, YUE J, ZHANG W G,etal. Biodiversity, ecosystem functioning and spatio-temporal scales[J].ActaPrataculturaeSinica, 2010,19(1):219-225.
[50] 梁 倍,邸 利,趙傳燕,等.祁連山天老池流域灌叢地上生物量空間分布[J].應用生態學報,2014,25(2):367-373.
LIANG B, DI L, ZHAO C Y,etal. Spatial distribution of aboveground biomass of shrubs in Tianlaochi catchment of the Qilian Mountains[J].ChineseJournalofAppliedEcology, 2014,25(2):367-373.
(編輯:裴阿衛)
Spatial Distribution ofPopuluseuphraticaBiomass in the Lower Reaches of Tarim River
WANG Xiyi1,XU Hailiang2*,PAN Cunde1,LING Hongbo2,YUAN Kaiye1
(1 Xinjiang Agricultural University, Urumqi 830052; 2 Xinjiang Institute of Ecology and Geography,Chinese Academy of Sciences,Urumqi 830011,China)
Based on sample-plot survey to thePopuluseuphraticain the lower reaches of Tarim River, we established the model between biomass and factors such as DBH and height; Besides, the distribution of biomass ofP.euphraticawas analyzed in the district; Finally, the total biomass was assessed based on the area ofP.euphraticaforest in the lower reaches of Tarim River. The results are as follows: (1) biomass model ofP.euphratica:B=128.09(D2h)0.6318(R2=0.892),which was high in precision, and the discrepancy between measured value and simulated value was non-significant (P>0.05), thus the model could be used to assess the biomass ofP.euphratica. (2) The biomass presented a trend that the biomass declining was relevant to the distance from river;P.euphraticadistributed within 700 m of river; and the biomass of this area accounted for 91.37% of total. (3) The biomass per unit area was 416.613 kg·hm-2in the lower reaches of Tarim river and the total biomass was 2.474×104t.
Populuseuphratica;biomass;spatial distribution;lower reaches of Tarim River
1000-4025(2016)11-2314-08
10.7606/j.issn.1000-4025.2016.11.2314
2016-07-27;修改稿收到日期:2016-09-21
國家自然科學基金(31370551,41471099, 31400466);中國科學院“西部之光”人才培養計劃(XBBS-2014-13);中國科學院特色研究所主要服務項目2課題2(TSS-2015-014-FW-2-2)
王希義(1987-),男,在讀博士研究生,主要從事生態需水與恢復生態學研究。E-mail:binzhouwxy@163.com.
*通信作者:徐海量,博士,研究員,碩士生導師,主要從事恢復生態學研究。E-mail:xuhl@ms.xjb.ac.cn
Q948.11;Q948.15+6
A
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
光學精密工程(2016年6期)2016-11-07 09:07:19
