常壓室溫等離子體誘變(ARTP)及高通量篩選高產蛋白酶米曲霉的初探
2017-01-06 08:40:26舒冬梅王德良宋緒磊尚柯
中國調味品 2016年12期
舒冬梅,王德良,宋緒磊,尚柯
(1.新疆農業大學 食品科學與藥學學院,烏魯木齊 830052;2.中國食品發酵工業研究院,北京 100015)
常壓室溫等離子體誘變(ARTP)及高通量篩選高產蛋白酶米曲霉的初探
舒冬梅1,王德良2*,宋緒磊2,尚柯1
(1.新疆農業大學 食品科學與藥學學院,烏魯木齊 830052;2.中國食品發酵工業研究院,北京 100015)
為提高醬油發酵菌種米曲霉的蛋白酶活力,采用實驗室保藏菌種H0米曲霉為誘變出發菌株,對H0菌株進行常壓室溫等離子體誘變(ARTP),誘變條件:功率120 W;氣流量10 L/min;10個時間梯度處理。結果表明:當誘變時間為140 s時,致死率接近90%,此時為最佳誘變時間。利用大豆球蛋白作為選擇培養基關鍵因子進行初篩和96孔板高通量復篩,選出3株高蛋白酶活力菌株H5,H12,H15,并且經過福林酚法驗證。最終篩選出米曲霉菌株H15具有高蛋白酶活力:酸性、中性、堿性蛋白酶活力分別為192.35,1816.31,3774.82 U/g,比出發菌株H0分別提高17.49%,19.08%,10.66%。
常壓室溫等離子體誘變(ARTP);高通量篩選;蛋白酶活力
醬油的釀造是以微生物為動力,將原料中的蛋白質、糖類、脂類等經過復雜的生物化學反應,生成多糖、單糖、氨基酸、多肽、酯類、風味物質等的一系列過程。目前醬油微生物中研究較多的微生物是米曲霉,主要是因為米曲霉產生的酶系豐富并且各種酶系綜合作用對醬油風味的形成起重要作用,其中起關鍵作用的是蛋白酶,米曲霉分泌的蛋白酶使原料中的蛋白質降解生成氨基酸,提高醬油氨基酸含量[1]。目前我國大多數醬油廠采用滬釀3.042以及經過誘變后選育的新菌株,但是其蛋白酶活力依然不夠高,導致我國醬油原料蛋白利用率在80%左右,而日式醬油這個指標已經達到90%[2]。
米曲霉菌種的誘變方法包括物理、化學方法,前人對于米曲霉菌種誘變方法有多種[3-7]。袁艷玲等人以米曲霉Y0為出發菌株,通過紫外線與DES復合誘變,0.4%干酪素培養基初步篩選,獲得1株高產中性蛋白酶菌株H2-5,其中性酶活力為2158 U/g,經10代培養,菌株中性蛋白酶活力具有良好的遺傳穩定性[8]。另外,近幾年ARTP(常壓室溫等離子體)誘變方法應用的興起為微生物誘變選育提供了新的方法和思路。由于ARTP誘變產生等離子體均勻,射流溫度低,需氦氣裝置,操作簡便,對細胞與生物大分子作用明顯等優勢[9,10],此方法應用廣泛,如化學凈化劑、熱敏感性醫療儀器消毒以及生物方面[11]。研究已報道其應用于酵母、大腸桿菌、刺糖多孢菌、米曲霉等微生物誘變并取得良好效果[12-15]。本試驗在參考各種文獻的基礎上結合生產實踐創新試驗方法,利用新型ARTP誘變育種方法誘變米曲霉,采用透明圈法和96孔板法建立高通量篩選突變菌株的方法,并獲得高產蛋白酶的米曲霉突變株,提高米曲霉分解蛋白的能力,從而提高醬油中氨基酸含量,提高醬油質量和國際競爭力。
1 材料和方法
1.1 菌株
米曲霉(Aspergillusoryzae)H0菌株為本實驗室保藏。
1.2 試劑與儀器
大豆球蛋白 百靈威公司;實驗室制麥汁、茚三酮、果糖、磷酸氫二鉀、磷酸二氫鈉、碘酸鉀、福林酚試劑(均為分析純) Bitopped 公司。
恒溫生化培養箱,水浴鍋(控溫精度±0.1 ℃) 北京天林恒泰科技有限公司;SW-CJ-2FD型雙人單面凈化工作臺 蘇州凈化設備有限公司;ARTP-II型ARTP誘變育種機 無錫源清天木生物科技有限公司;渦旋儀MVS-1 北京東方開物科學器材有限公司;立式電熱壓力蒸汽滅菌器 上海申安醫療器械廠;UV-1700可見分光光度計 島津公司;PHS-3C pH計 雷磁有限公司;恒溫電子天平AL204 梅特勒-托利多儀器(上海)有限公司。
1.3 培養基的配制和滅菌
菌種保藏培養基:馬鈴薯培養基。
大豆球蛋白篩選培養基[16]:大豆球蛋白1 g,磷酸二氫鈉1.07 g,磷酸氫二鈉1 g,硫酸鎂0.5 g,硫酸亞鐵0.01 g,瓊脂20 g,蒸餾水1000 mL,121 ℃滅菌30 min。
0.1%干酪素培養基:干酪素1 g,磷酸二氫鈉1.07 g,磷酸氫二鉀0.36 g,硫酸鎂0.5 g,硫酸亞鐵0.002 g,蒸餾水1000 mL,121 ℃滅菌 30 min。
麩曲培養基:250 mL的三角瓶,裝料量為麩皮15 g,水15 mL,自然pH,121 ℃滅菌30 min。
1.4 氨基氮測量試劑盒溶液
顯色劑:稱取磷酸氫二鈉(Na2HPO4·12H2O)20 g,磷酸二氫鉀(KH2PO4)12 g,超聲溶解,加入茚三酮1.0 g和果糖0.6 g(精確至0.01 g),混勻,用水溶解并定容至200 mL。將溶液貯藏在棕色瓶中,放置4 ℃冰箱保存,1周內使用有效。
稀釋溶液:稱取碘酸鉀(KIO3)2 g(精確至0.01 g)溶于620 mL水中,超聲溶解,加入99.7%乙醇380 mL,混勻,于4 ℃貯存。
甘氨酸標準儲備液(1.072 g/L):稱取甘氨酸0.1072 g,用水溶解并定容至100 mL,于0~5 ℃貯存。
甘氨酸標準使用液:吸取甘氨酸標準貯備液1 mL,用水稀釋至100 mL。該標準使用液含游離氨基氮2 mg/L,使用時現配制。
1.5 生長曲線繪制
將米曲霉從試管斜面轉接到PDA平板上,28 ℃培養72 h。將活化好的米曲霉用接種環接3環到裝有已滅菌12°麥汁的三角瓶中,自保藏斜面中挑取3環米曲霉孢子接入裝有10 mL無菌麥汁培養基的試管中,搖勻,制成孢子懸液,于28 ℃培養箱中培養活化24 h。將10 mL菌懸液倒入裝有300 mL無菌麥汁培養基的三角瓶中,搖勻,制成孢子懸液,于28 ℃培養箱中搖床培養。
取14個2 mL的離心管,稱重并分別記錄離心管的重量,精確到0.0001 g。吸取1 mL的12°麥汁和200 μL的1.5×105的活化菌液,置于28 ℃搖床培養。于培養后的第0,4,8,12,16,20,24,28,32,36,40,48,52,56 h取出相應標號的離心管,置于4 ℃冰箱保存。待全部離心管取出,統一8000 r/min,離心15 min。60 ℃烘干3 h,稱量重量,精確到0.0001 g。

1.6 ARTP誘變
1.6.1 菌懸液制作
將活化好的米曲霉菌種接種于PDA培養基上,培養溫度28 ℃,培養18~24 h是對數期,本試驗采用對數期36 h時的米曲霉,將對數期的米曲霉用0.8%生理鹽水調其濃度,在光學顯微鏡下計數,將菌濃度調至1×106~1×108cfu/mL。本試驗最終米曲霉菌孢子濃度為2×106cfu/mL。將配制的2×106cfu/mL孢子懸浮液取95 μL,取50%甘油5 μL,兩者均勻混合,以保護米曲霉孢子。
1.6.2 ARTP照射劑量的確定
采用氦氣為工作氣體,使載片與等離子體發生器射流出口間距約為2 mm,功率為120 W,氣流量10 L/min,對10 μL孢子(106cfu/mL)懸液進行誘變,處理時間設置為0,20,60,80,100,120,150,180,210,240 cfu/mL,稀釋孢子懸液為2×104,2×103,2×102cfu/mL,取 100 μL各稀釋倍數孢子懸浮液涂布選擇培養皿,28 ℃培養3天計數,計算致死率(3個重復的平均值)。

1.7 初篩方法
挑選ARTP誘變140 s的平板上的菌落點種于大豆球蛋白初篩選培養基,28 ℃培養72 h觀察記錄透明圈和菌落大小,計算出K值。挑取K值較大并且孢子數較多的菌落接種于馬鈴薯試管斜面培養基,于30 ℃培養72 h,試管內布滿淡綠色米曲霉菌絲停止,然后于4 ℃冰箱保存備用。
1.8 高通量復篩菌種
1.8.1 高通量復篩菌種的培養
將初選的15個于菌株PDA平板上培養72 h,制備每個菌株的菌懸液,菌濃度為2.5×106cfu/mL。采用18 cm×18 mm的試管加入1~15號誘變菌株菌懸液1 mL和1 mL 0.1%干酪素培養基,以試管中加入2 mL為對照試驗,每個處理重復3次。
于28 ℃恒溫生化培養箱搖床培養36 h后,從每個試管中取出40 μL液體置于96孔板中,加入10 μL發色劑,100 ℃水浴準確加熱16 min,在(20±0.1) ℃水浴中冷卻20 min。再各加入稀釋溶液50 μL,充分混勻。用空白試管水調節儀器零點,于波長570 nm下測量吸光度,測量應在30 min內完成。
樣品中的α-氨基氮含量按下式計算:
式中:X為麥芽汁的α-氨基氮含量,mg/L;A1為樣液的平均吸光度;A2為甘氨酸標準使用液的平均吸光度;2為甘氨酸標準使用液中α-氨基氮的含量,mg/L;n為樣液的稀釋倍數。
1.8.2 高通量復篩方法的驗證
采用福林酚法對高通量復篩出的H5,H12,H15 3個菌株進行蛋白酶活力測定[17]。
1.9 突變菌株遺傳穩定性實驗
將總酶活力高的突變菌株在麩曲培養基中連續傳代培養9次,培養條件為30 ℃,60 h。依次稱取一定質量的第1,3,5,7,9代種曲的酸性、中性、堿性蛋白酶活力進行測定,檢測突變菌株的遺傳穩定性。
2 結果與分析
2.1 米曲霉生長曲線繪制

圖1 米曲霉菌絲Fig.1 Aspergillus oryzae hypha

圖2 米曲霉菌株的生長曲線Fig.2 Growth curve of Aspergillus oryzae
由圖2可知,0~12 h米曲霉生長處于遲緩期(停滯期);16~48 h菌體生長迅速,處于對數生長期;52~60 h 處于平衡期。
微生物生長曲線是以微生物數量(活細菌個數或細菌重量)為縱坐標,培養時間為橫坐標繪制的曲線。典型的微生物生長曲線包括四個時期:遲緩期、對數期、穩定期、衰亡期。一般來說,大多數細菌的繁殖速度都很快,它的生長曲線是以細菌數目為縱坐標,以時間為橫坐標。然而霉菌在液體培養時(150 r/min)大量菌絲生長會相互交織成球或塊狀,從而統計微生物個數或測定OD值不能反映霉菌的各個生長時期,因此,霉菌的生長曲線制作采用其生長量為縱坐標,時間為橫坐標。
2.2 致死率曲線
采用平板活菌計數法繪制米曲霉菌株的ARTP誘變致死率曲線,ARTP誘變時間與致死率的關系曲線見圖3。
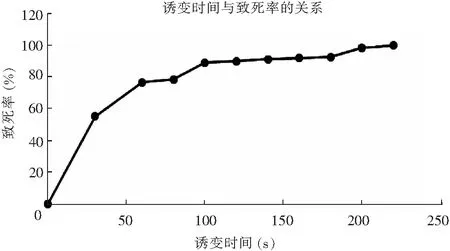
圖3 ARTP誘變時間與致死率的關系曲線Fig.3 Relation between ARTP mutation time and lethality
由圖3可知,等離子體對米曲霉菌株的致死率有明顯劑量效應關系,隨時間延長,在1~100 s范圍內致死率迅速增加;誘變時間為100 s時,致死率達到89.12%;140 s時致死率達到91.22%;200 s時其接近100%。為獲取較高的正突變率、較高蛋白酶活力菌株,本試驗將致死率設定為90%以上[18],因此,選取140 s為ARTP誘變時間。
2.3 初篩結果
誘變前后米曲霉在初選培養基上生長見圖4和圖5。

圖4 誘變前米曲霉在初選培養基上生長 Fig.4 Growth of Aspergillus oryzae on initial screening culture medium before mutation

圖5 誘變后米曲霉在初選培養基上生長Fig.5 Growth of Aspergillus oryzae on initial screening culture medium after mutation

測量值D(cm)d(cm)K對照0.90.81.13H11.21.11.09H20.60.51.20H30.80.71.14H41.10.91.22H51.40.72.00H60.60.51.20H710.91.11H80.90.81.13H91.40.81.75H101.50.91.67H111.30.81.63H121.50.81.88H131.10.61.83H141.20.91.33H151.50.72.14
ARTP誘變后菌種有正突變,菌落長勢好并且K值比對照值大;有負突變,菌落生長速度慢且K值比對照值小,將負突變的菌株舍棄,正突變菌株測量其菌落直徑和透明圈直徑并且計算K值。由表1可知,突變菌株H5,H12,H15 3個菌株的K值較大,分別為2.00,1.88,2.14,其中菌株H15的K值為2.14,最大。僅僅依據初篩培養基的K值不能最終確定米曲霉蛋白酶活的高低,還需進一步采用蛋白酶活測定方法驗證,確定蛋白酶活力高的菌種。
2.4 甘氨酸標準曲線

圖6 甘氨酸標準曲線Fig.6 Standard curve of glycine
甘氨酸標準曲線是甘氨酸不同濃度標準物質的儀器與試劑體系響應之間的函數關系。建立甘氨酸標準曲線是為了推導待測物的濃度。在分析食品化學實驗中,常用標準曲線法進行物質定量分析,一般情況下標準工作曲線呈直線分布。只有標準曲線與校正曲線重合度越好的條件下,測量結果越準確,這樣才可以用標準曲線來代替校正曲線。由圖6可知,甘氨酸標準曲線為y=0.1677x+0.0643,R2=0.992,甘氨酸含量與吸光值的關系幾乎呈線性關系,并且相關性較好,可以用此標準曲線來測定待測物的α-氨基酸含量。
2.5 復篩結果
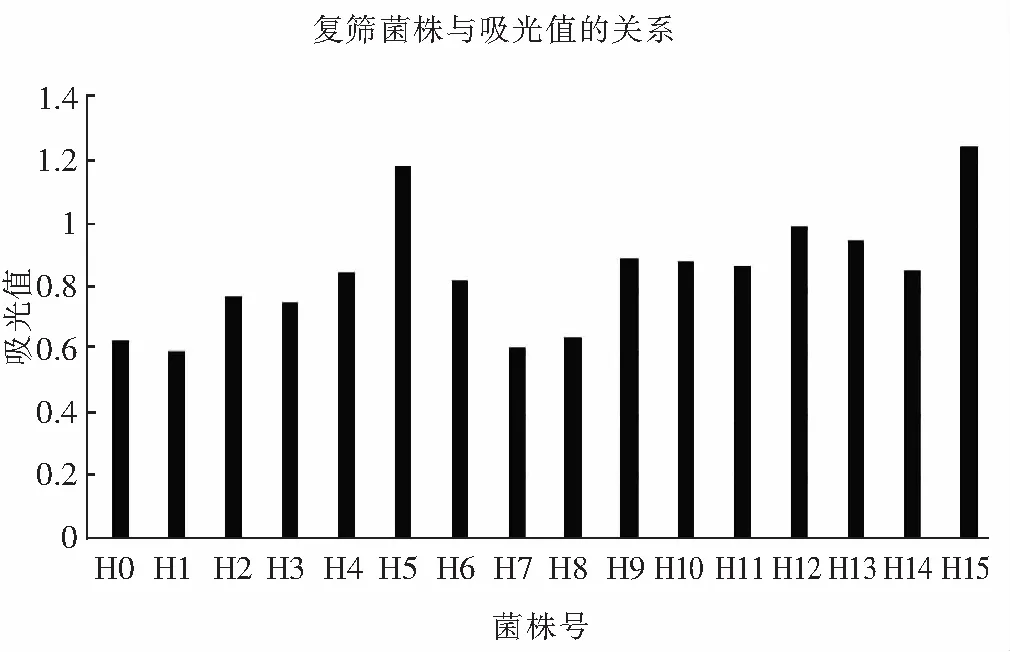
圖7 菌株高通量復篩Fig.7 High-throughput for secondary screening of strain

菌株酸性蛋白酶活中性蛋白酶活堿性蛋白酶活H0163.721525.283411.19H5197.481536.943486.25H12169.421603.593724.56H15208.431792.793825.36
由圖7可知,采用吸光度法測定α-氨基氮含量,根據甘氨酸標準曲線得到α-氨基氮含量在0~10 mg/L的范圍內,吸光值在0~1.6范圍內,吸光度隨其濃度增大而增大,并且遵循y=0.1677x+0.0643(R2=0.992)直線關系。米曲霉蛋白酶活越高,分解酪蛋白變成α-氨基酸的含量就越高,α-氨基酸與茚三酮反應。所以,可以依據米曲霉與酪蛋白反應后測得的吸光值來判斷米曲霉的綜合蛋白酶活力。由表2可知,ARTP誘變后菌H5,H12,H15菌株總蛋白酶活力明顯高于出發菌株H0菌株。菌株H5的酸性、中性和堿性蛋白酶活力分別為197.48,1536.94,3486.25 U/g;H12的三種蛋白酶活分別為169.42,1603.59,3724.56 U/g;H15的3種蛋白酶活分別為208.43,1792.79,3825.36 U/g。根據米曲霉酶活力測定,最終3個菌株的蛋白酶活與采用96孔板篩選結果一致,即H5,H12,H15菌株總蛋白酶活力明顯高于出發菌株H0并且高于其他菌株。
2.6 米曲霉遺傳穩定性檢測
經過1~9代傳代培養,種曲制備和福林酚法測定蛋白酶活力等步驟來考察菌株的遺傳穩定性。

圖8 H5菌株遺傳穩定性Fig.8 Genetic stability of strain H5
由圖8可知,菌株H5的酸性蛋白酶活力從197.48 U/g降至185.12 U/g;中性蛋白酶活力從1536.94 U/g降至1247.52 U/g;堿性蛋白酶活力從3486.25 U/g降至3317.22 U/g。

圖9 H12菌株的遺傳穩定性Fig.9 Genetic stability of strain H12
由圖9可知,菌株H12的酸性蛋白酶活力從169.42 U/g升至175.89 U/g;中性蛋白酶活力從1603.59 U/g降至1580.42 U/g;堿性蛋白酶活力從3524.56 U/g降至3527.98 U/g。

圖10 H15菌株的遺傳穩定性Fig.10 Genetic stability of strain H15
由圖10可知,菌株H15的酸性蛋白酶活力從208.43 U/g升至192.35 U/g;中性蛋白酶活力從1792.79 U/g升至1816.31 U/g;堿性蛋白酶活力從3825.36 U/g升至3774.82 U/g。
綜合3個菌株的3種蛋白酶活的變化趨勢可知,H5菌株的3種蛋白酶活力都有下降的趨勢,并且中性和堿性蛋白酶力下降大,分別下降289.42,169.03 U/g;H12菌株的3種蛋白酶活力相對較穩定,酸性和堿性蛋白酶活力下降,中性蛋白酶活下降,但數值變化都不大(<40 U/g);H15菌株的3種蛋白酶活都比較高,并且傳代培養后,蛋白酶活力穩定性較好,波動范圍在51 U/g之內,并且H15菌株的酸性蛋白酶活力最終為192.35 U/g,增長率為17.49%;中性蛋白酶活力1816.31 U/g,增長率為19.08%;堿性蛋白酶活力3774.82 U/g,增長率為10.66%。3種蛋白酶活力遺傳穩定性較好,波動范圍較小且綜合蛋白酶活力高于菌株H5和H12,所以,最終篩選出菌株H15為實驗室出發菌株發酵醬油,進一步探索菌株的其他特性。
3 結論
采用96孔板高通量篩選蛋白酶活力較高菌株的方法與國家標準采用的福林酚法測定蛋白酶活力結果一致,96孔板高通量篩選蛋白酶活力可作為一種復篩方法,為高蛋白酶活力米曲霉篩選提供了一種簡便、快速、相對準確的方法。
ARTP誘變米曲霉菌株H0具有良好效果,時間為140 s時其致死率接近90%。
通過初篩和復篩,選育出米曲霉菌種H15,其酸性蛋白酶活力最終為192.35 U/g,中性蛋白酶活力為1816.31 U/g,堿性蛋白酶活力為3774.82 U/g,分別比出發菌株H0增長17.49%,19.08%,10.66%,為后續醬油生產提供了較高蛋白酶活力米曲霉菌種。本試驗為米曲霉菌種誘變選育提供了新方法的理論參考,并且構建了高蛋白酶活力菌株高通量篩選方法,為醬油釀造提供具有高蛋白酶活力的菌種。
[1]陳紅梅,潘力.米曲霉3.042種曲蛋白酶特性研究[J].中國調味品,2009,34(3):63-71.
[2]徐德峰,趙謀明.中國醬油菌株 AS 3.951的改良現狀與思考[J].中國調味品,2009,34(5):32-38.
[3]Yasuhara A, Ogawa A,Tanaka T,et al.Production of neutral protease fromAspergillusoryzaeby a novel cultivation method on a microporous membrane[J].Biotechnology Techniques, 1994,8(4):249-254.
[4]鄧靜,吳華昌,吳明霞,等.米曲霉產蛋白酶條件的優化[J].中國釀造,2008,27(12):51-53.
[5]孫春華.米曲霉菌株的誘變選育及其制劑的應用研究[D].泰安:山東農業大學,2008.
[6]唐潔.利用原生質體融合技術進行米曲霉新菌株的選育[D].成都:西華大學,2007.
[7]韓志雙,劉軍,郇阿梅,等.應用基因組改組技術選育米曲霉酸性蛋白酶高產菌株[J].中國調味品,2015,40(1):18-22.
[8]袁艷玲,楊生玉,錢深,等.米曲霉中性蛋白酶高產菌株的復合誘變選育[J].科技信息,2013,15(1):69-58.
[9]Li H P, Li G, Sun W T. Radio-frequency,atmospheric-pressure glow discharges:producing methods,characteristics and applicationsin bio-medical fields[C].Complex Systems: 5th International Workshop on Complex Systems. AIP Conference Proceedings, 2008,98(2):584-591.
[10]Li G, Li H P, Wang L Y, et al. Genetic effects ofradio-atmospheric pressure glow discharges with helium[J].Appl Phys Lett, 2008, 92(22):221504.
[11]Laroussi M. Nonthermal decontamination of biological media by atmospheric-pressure plasmas: review,analysis, and prospects[J].IEEE Transactions on Plasma Science, 2002, 30(4): 1409-1415.
[12]王方方,孫沛勇,銀會娟,等.常壓室溫等離子體快速誘變酒精酵母及其突變株的特性研究[J].中國釀造,2013,10(8):117-119
[13]Liu Rongming, Liang Liya, Ma Jiangfeng, et al. An engineeringEscherichiacoli mutant with high succinic acid production in the defined medium obtained by the atmospheric and room temperature plasma[J].ProcessBiochemistry, 2013, 48(11): 1603-1609.
[14]萬青,曹偉佳,張常青,等.常壓室溫等離子體誘變高效利用木糖產丁二酸菌株[J].生物工程學報,2013,29(11):1692-1695.
[15]陳錫劍,陳建華.常壓室溫等離子體誘變選育高產曲酸米曲霉[J].化學與生物工程,2015,32(5):52-60.
[16]高獻禮,趙斯薇,孫鵬飛,等.醬油渣蛋白質的分離、鑒定和氨基酸組成特征研究[J].現代食品科技,2013,29(10):2512-2516.
[17]SB/T 10317-1999,蛋白酶活力測定法[S].
[18]范新蕾,肖成建,顧秋亞,等.ARTP誘變選育葡萄糖氧化酶高產菌株及發酵條件優化[J].工業微生物,2015,45(1):15-19.
Initial Study on High Proteinase-producing Aspergillus oryzae with ARTP Mutation and High-throughput Screening
SHU Dong-mei1, WANG De-liang2*, SONG Xu-lei2, SHANG Ke1
(1.College of Food Science and Medicine, Xinjiang Agricultural University, Urumqi 830052, China; 2.China National Research Institute of Food and Fermentation Industries, Beijing 100015, China)
To increase the proteinase activity ofAspergillusoryzaeduring the brewing of sauce, takeAspergillusoryzaestrain H0 from laboratory as the starting strain, and then ARTP mutation of strain H0 is carried out. The process conditions are as follows: power is 120 W, air flow volume is 10 L/min, 10 time gradient. The results show that the lethality is about 90% as the mutation time is 140 s, and it is the best time. Three strains such as H5, H12, H15 are selected by initial screening(take glycinin as the key factor in the selectivity culture medium) and high-throughput secondary screening (by 96 hole plate), and checked by Folin-phenol method. Eventually,Aspergillusoryzaestrain H15 with high proteinase activity is selected; its acidic, neutral and alkaline proteinase activity is 192.35, 1816.31,3774.82 U/g respectively, which is increased by 17.49%, 19.08%,10.66% respectively compared with the starting strain H0.
ARTP; high-throughput screening; proteinase activity
2016-06-08 *通訊作者
科技部科研院所基金項目(2014EG111217)
舒冬梅(1988-),女,碩士,研究方向:食品生物技術。
TS201.25
A
10.3969/j.issn.1000-9973.2016.12.014
1000-9973(2016)12-0067-07
