擬南芥苯丙氨酸解氨酶基因的克隆與表達分析
2017-01-06 02:21:16趙丹
山西農業科學 2016年12期
趙丹
(山西農業大學信息學院,山西太谷030800)
擬南芥苯丙氨酸解氨酶基因的克隆與表達分析
趙丹
(山西農業大學信息學院,山西太谷030800)
苯丙烷途徑是植物體內廣泛存在的次生代謝途徑,此代謝過程中的第1個關鍵酶為苯丙氨酸解氨酶(Phenylalanine ammonia-1yase,PAL)。苯丙氨酸解氨酶通過對基因的起始轉錄、頻率的控制和對外界誘導因子的響應來調控基因的表達,并且其表達水平對于下游黃酮類物質的合成起著非常重要的作用。在一定條件下,苯丙氨酸解氨酶可利用其特殊的酶學性質逆向催化生產L-苯丙氨酸。通過一系列的生物信息學分析,確定擬南芥中苯丙氨酸解氨酶的存在,并進行基因的克隆及表達分析,為L-苯丙氨酸的生產提供新的物種來源。
擬南芥;苯丙氨酸解氨酶;生物信息學;克隆
苯丙氨酸解氨酶是1961年在大麥中發現的[1],其是苯丙烷途徑的第1個關鍵酶,在各種各樣的植物中均有存在,而且在少量微生物中也有存在。苯丙氨酸解氨酶是一種將苯丙氨酸轉化為酚類化合物的重要蛋白酶,決定了酚類化合物合成的速率[2]。同時,苯丙氨酸解氨酶通過對基因的起始轉錄、頻率的控制以及對外界誘導因子的響應來調控基因的表達,并且其表達水平對于下游黃酮類物質的合成起著非常重要的作用。在酸堿值為10,過量的銨根離子和酚類化合物同時存在的條件下,苯丙氨酸解氨酶可逆向催化酚類化合物加氨生成L-苯丙氨酸[3-5],這種特殊的酶學性質被廣泛地應用在生物化工領域,以創造更高的價值。
提高L-苯丙氨酸產量的一個重要方法是在眾多的苯丙氨酸解氨酶源中篩選出一種活力較高并且穩定性好的物種[6-8]。擬南芥的生長過程較短,培養條件也相對較低,便于大量培育,是分子遺傳性與農業研究中良好的材料[9]。同時,由于擬南芥基因高度純合的特點,使用不同的物理因素或者化學因素對擬南芥進行處理時其容易產生突變,生成突變植株,便于對其進行代謝功能及調控酶的研究[10-11]。建立擬南芥中苯丙氨酸解氨酶的研究機制,為L-苯丙氨酸的生產提供新的物種來源,對于我國的農業研究和農作物的發展具有重要的影響及意義。
1 材料和方法
1.1 材料
供試材料為擬南芥幼苗。
1.2 試劑及儀器
1.2.1 試劑PCR引物合成于生工公司;反轉錄試劑盒、TE緩沖液購于TAKARA公司;苯酚、氯仿、戊酮、異丙醇無水乙醇試劑均為分析純。
1.2.2 主要儀器培養箱、PCR儀、-20℃冰箱、冷凍離心機、滅菌鍋、電子天平、pH計、電泳儀、凝膠成像系統。
1.2.3 主要試劑配制提取緩沖液:Rris-HCl,EDTA,NaCl,SDS;氯仿-戊酮-乙醇體積比為80∶4∶16。
1.3 方法
1.3.1 擬南芥苯丙氨酸解氨酶序列查找在NCBI中通過苦蕎苯丙氨酸解氨酶的BLAST分析,查找可能的擬南芥苯丙氨酸解氨酶序列。其他物種的苯丙氨酸解氨酶序列從NCBI蛋白數據庫中獲得。
1.3.2 生物信息在線分析網站http://swissmodel. expasy.org/Swiss-model在線數據庫;http://www.ncbi. nlm.nih.gov/NCBI在線數據庫;http://prosite.expasy. org/PROSITE在線數據庫;http://www.expasy.org/Expasy在線數據庫;https://npsa-prabi.ibcp.fr/cgi-bin/ NPS@在線數據庫。
1.3.3 生物信息分析軟件預測蛋白跨膜結構利用TMHMM軟件;信號肽預測工具利用SignalP軟件;亞細胞定位分析利用WOLFPSPORT軟件;蛋白同源性比對利用NCBI中的BLASTX程序;蛋白序列比對分析利用DNA MAN軟件;蛋白理化性質分析利用Protparam工具。
1.3.4 擬南芥基因組的快速提取在50 mL離心管中加入20 mL提取緩沖液,50℃水浴預熱;取擬南芥幼苗5~15 g,粉碎;加入25 mL氯仿-戊酮-乙醇溶液,顛倒混勻;在室溫下5 000 r/min離心5 min;移取上清液放置50 mL離心管中,加入1倍體積異丙醇,搖動混勻,在室溫下靜置,出現絮狀DNA沉淀;在1.5 mL離心管中加入1 mL TE,用玻璃棒取出DNA絮團,用吸水紙吸干水分,再加入含TE的離心管中,使DNA溶解于TE中。
1.3.5 擬南芥總RNA的提取收取擬南芥放在滅過菌的研缽中,加入液氮搗碎,迅速轉入1.5 mL離心管中,以3 000 r/min離心2 min,棄上清,將沉淀彈起;加入1 mL的Trizol試劑于1.5 mL離心管中,輕搖混勻,放置5 min;加入200 mL的氯仿到離心管中,用手劇烈搖晃15 s,在室溫下放置10 min,管內溶液分層,于4℃,12 000 r/min離心15 min;將上層水相轉移到干凈的1.5 mL離心管中后,吸取等體積的異丙醇加入離心管中,上下顛倒搖晃至混勻,室溫下放置10 min,4℃12 000 r/min離心15 min,此時可見RNA沉淀;棄上清,加入用RNA酶無菌水配制的75%乙醇,上下顛倒搖晃洗滌沉淀,4℃12 000 r/min離心10 min;棄上清,無菌條件下干燥5 min,將RNA溶于30 μL的去RNA酶無菌水中。
1.3.6 擬南芥總cDNA的合成參照TAKARA公司的反轉錄試劑盒,以提取到的總RNA為模板反轉錄為cDNA,反轉錄擴增體系為37℃反轉錄15 min,85℃5 s終止反應,產物4℃保存(表1)。

表1 擬南芥總cDNA的合成
1.3.7 擬南芥中PAL基因的克隆以擬南芥cDNA為模板,利用PCR技術擴增PAL基因的開放閱讀框,并使用半定量方式檢驗PAL基因在不同時期擬南芥幼苗中的表達情況(表2)。
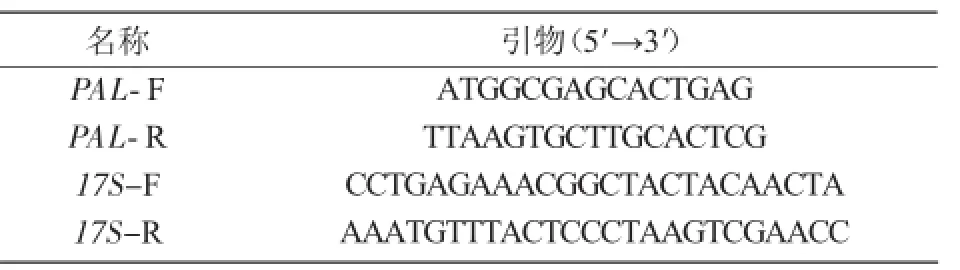
表2 試驗所用引物
2 結果與分析
2.1 擬南芥苯丙氨酸解氨酶序列
在NCBI中通過苦蕎苯丙氨酸解氨酶的BLAST分析,查找可能的擬南苯丙氨酸解氨酶序列,其結果如下。
MEINGAHKSNGGGVDAMLCGGDIKTKNMVIN AEDPLNWGAAAEQMKGSHLDEVKRMVAEFRKPV VNLGGETLTIGQVAAISTIGNSVKVELSETARAGVN ASSDWVMESMNKGTDSYGVTTGFGATSHRRTKNG VALQKELIRFLNAGIFGSTKETSHTLPHSATRAAML VRINTLLQGFSGIRFEILEAITSFLNNNITPSLPLRGTI TASGDLVPLSYIAGLLTGRPNSKATGPNGEALTAEE AFKLAGISSGFFDLQPKEGLALVNGTAVGSGMASM VLFETNVLSVLAEILSAVFAEVMSGKPEFTDHLTHR LKHHPGQIEAAAVMEHILDGSSYMKLAQKLHEMD PLQKPKQDRYALRTSPQWLGPQIEVIRYATKSIERE INSVNDNPLIDVSRNKAIHGGNFQGTPIGVSMDNTR LAIRAIGKLMFAQFSELVNDFYNNGLPSNLTASRNP SLDYGFKGAEIAMASSIEREINSVNDNPLIDVSRNK AIHGGNFQGTPIGVSMDNTRLAIRAIGKLMFAQFSELVNDFYNNGLPSNLTASRNPSLDYGFKGAEIAMAS YCSELQYLANPVTSHVQSAEQHNQDVNSLGLISSR KTSEAVDILKLMSTTFLVAICQAVDLRHLEENLRQT VKNTVSQVAKKVLTTGVNGELHPSRFCEKDLLKV VDREQVYTYADDPCSATYPLIQKLRQVIVDHALVN GESEKNAVTSIFHKIGAFEEELKAVLPKEVEAARA AYDNGTSAIPNRIKECRSYPLYRFVREELGTELLTG EKVTSPGEEFDKVFTAICEGKIIDPMMECLNEWNG APIPIC
2.2 蛋白質的理化分析
為了更加準確地了解擬南芥苯丙氨酸解氨酶,研究其理化性質是必不可少的,理化性質更能直接簡單地為植物學研究提供很大的幫助。利用生物信息分析軟件Protparam對擬南芥苯丙氨酸解氨酶進行理化分析,其結果如表3所示。
由表3可知,擬南芥苯丙氨酸解氨酶是由725個氨基酸組成的蛋白質,其分子量為78 782.7 Da,等電點PI為5.96。
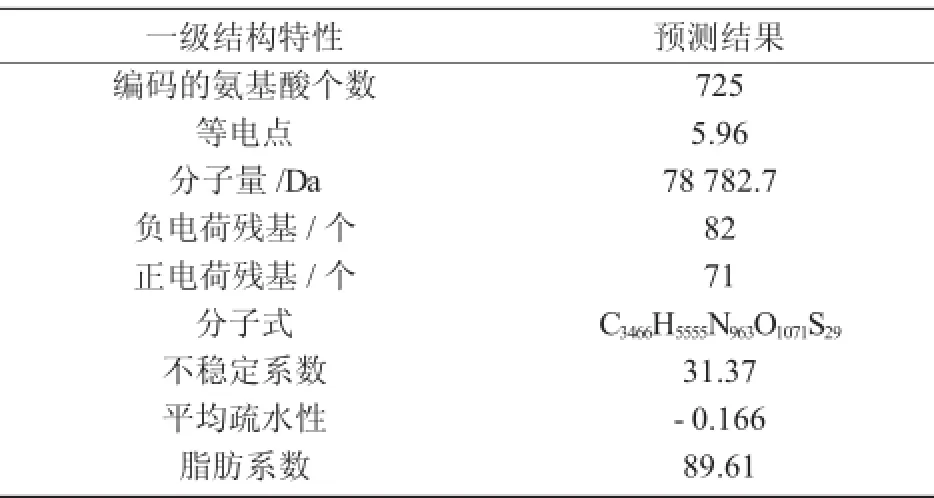
表3 蛋白質的理化分析
2.3 蛋白的功能域及保守域分析
使用PROSITE數據庫[12]對擬南芥苯丙氨酸解氨酶氨基酸序列進行功能域和功能位點分析,以確定其是否具有保守功能域及其功能域的具體位置(圖1)。由圖1可知,擬南芥苯丙氨酸解氨酶氨基酸序列在第207~223位點存在一段保守功能域,序列為GTITASGDLVPLSYIAG,而此序列為丙氨酸解氨酶家族蛋白的特征序列,證明查找的蛋白序列存在苯丙氨酸解氨酶的功能基礎。

2.4 蛋白質疏水性預測
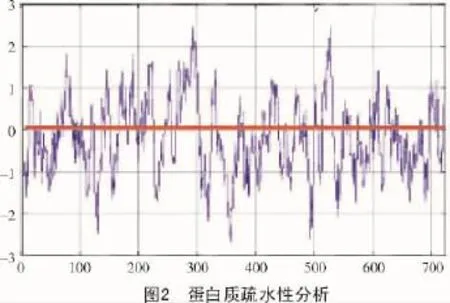
采用ProtScale(http://ca.expasy.org/tools/protsca le.html)軟件[13]對擬南芥苯丙氨酸解氨酶進行疏水性分析(圖2)。圖2橫軸表示的是氨基酸的位置,縱軸代表的是氨基酸平均疏水指標,并且圖中的正值和負值分別表示的是疏水性和親水性;由圖2可知,較粗的橫線為親疏水性零點,此蛋白序列在親疏水性正負兩側分布均勻,結合蛋白質一級結構分析可以得出,此蛋白總體沒有表現出明顯的親水性或疏水性,符合苯丙氨酸解氨酶的特性。
2.5 蛋白跨膜結構預測
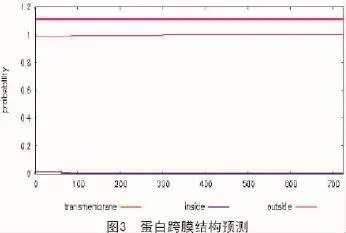
本試驗利用TMHMM在線生物信息分析網站[14]中專業預測蛋白跨膜結構的軟件,對擬南芥苯丙氨酸解氨酶進行蛋白跨膜結構預測(圖3)。圖3橫軸表示的是氨基酸的位置,縱軸代表的是跨膜結構的可能性,圖3結果顯示,擬南芥苯丙氨酸解氨酶所有位點均沒有存在跨膜結構的可能性。結合跨膜結構預測結果以及蛋白質疏水性預測結果表明該蛋白亞單位不含有跨膜結構域。
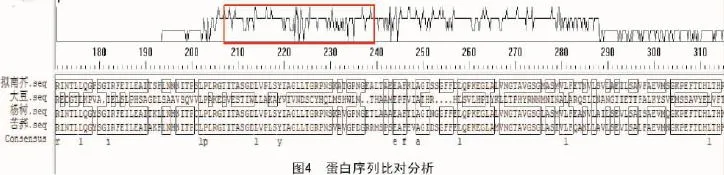
2.6 蛋白序列比對分析
通過NCBI查找發現,擬南芥中苯丙氨酸解氨酶有725個氨基酸,用DNA MAN軟件[15]把其氨基酸的序列和大豆、楊樹、苦蕎中的苯丙氨酸解氨酶蛋白序列進行對比,結果顯示,不同生物物種的苯丙氨酸解氨酶蛋白序列存在一定的序列一致性,并存在一個較保守的結構域,主要集中在210~230個氨基酸之間(圖4)。這樣的序列一致性符合苯丙氨酸解氨酶保守功能域的特性,證明擬南芥中苯丙氨酸解氨酶的結構域是保守的,能夠執行相應的生物學功能,具有和其他物種苯丙氨酸解氨酶相似的生理意義。
2.7 蛋白質的三級結構分析
蛋白質的三級結構是蛋白質在二級結構的基礎上進一步組成的,它具有疏水核位于內部、親水鏈位于內部的特點[16-17]。利用Swiss-model在線數據庫[18](http://swissmodel.expasy.org/)對擬南芥苯丙氨酸解氨酶進行三級結構預測,其結果如圖5所示。
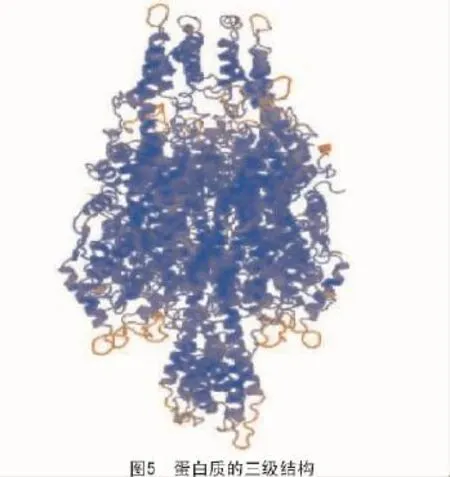
從圖5可以看出,擬南芥苯丙氨酸解氨酶主要由α-螺旋組成,結構十分穩定。而α-螺旋在DNA結合基序(DNA binding motifs)中有非常重要的作用,比如在鋅指結構、亮氨酸拉鏈、螺旋-轉角-螺旋等基序中都含有α-螺旋。擬南芥苯丙氨酸解氨酶含有豐富的α-螺旋結構,其可能指導苯丙氨酸解氨酶對相關基因的起始轉錄、頻率的控制和對外界誘導因子的響應,從而調控黃酮類物質的合成。
2.8 擬南芥中苯丙氨酸解氨酶基因序列的克隆
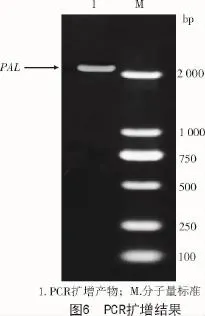
上述的試驗驗證了通過生物信息學方式查找到的擬南芥中苯丙氨酸解氨酶具有保守的功能結構域,是具有生物功能和意義的活性蛋白。為了進一步研究擬南芥中苯丙氨酸解氨酶的功能,進行基因過表達或轉基因操作,以PAL-F與PAL-R為引物(表2),以擬南芥cDNA作為模板,PCR擴增出約2 178 bp的PAL基因片段(圖6)。
2.9 擬南芥中苯丙氨酸解氨酶基因的表達分析
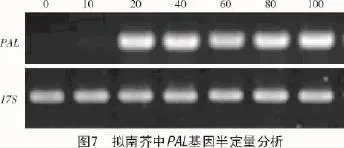
取擬南芥第0,10,20,40,60,80,100天的苗株進行qRT-PCR分析(圖7),以17S為對照,結果表明,擬南芥中苯丙氨酸解氨酶基因在幼苗前20 d表達量不高,20 d后開始大量表達,并保持穩定的表達趨勢,這可能是由次生代謝的開始所引起的。證明在擬南芥中苯丙氨酸解氨酶能夠穩定存在,為L-苯丙氨酸的生產提供了可能的新的物種來源。
3 結論
生物信息技術的使用使我們對龐大基因庫的分析變得更加高效、快速,這種分析方法可以使日后的研究實現“事半功倍”的效果,能夠為之后試驗研究提供重要線索,提升了日常工作效率[19-20]。本研究運用生物信息學分析軟件從結構組成、基本理化性質、疏水性/親水性、三級結構以及功能域等方面對擬南芥苯丙氨酸解氨酶進行預測和分析,結果表明,擬南芥PAL基因是由2 628個堿基所組成,其所編碼含有725個氨基酸殘基的多肽序列,蛋白分子量為78 782.7 Da;氨基酸中大部分沒有顯現出明顯的親水性或疏水性,并且等電點PI為5.96;該蛋白不可以進行跨膜運動,不含有跨膜結構域。對擬南芥苯丙氨酸解氨酶氨基酸序列進行功能域和功能位點分析,結果發現,其含有保守結構域。基因的克隆和表達譜分析表明,擬南芥中苯丙氨酸解氨酶基因在幼苗前20 d表達量不高,20 d后開始大量表達,并保持穩定的表達趨勢,這可能是由于次生代謝的開始所引起的。證明在擬南芥中苯丙氨酸解氨酶能夠穩定存在,為L-苯丙氨酸的生產提供了可能的新的物種來源。
[1]蔡朱男,余應年,羅建紅.水稻苯丙氨酸氨解酶基因在大腸桿菌BL21DE3中的表達[J].中國病理生理雜志,2000(1):12-16.
[2]江柯.黏紅酵母CIBASA1401苯丙氨酸解氨酶的分離純化與性質研究[D].成都:四川大學,2004.
[3]李冀新,張超.L-苯丙氨酸的生產及研究進展[J].氨基酸和生物資源,2006(2):51-54.
[4]劉衛紅,王燕,杜何為.銀杏葉苯丙氨酸解氨酶分離純化條件的研究[J].湖北農學院學報,2004(3):196-198.
[5]繆元穎,李化,楊亞力.苯丙氨酸解氨酶高活性紅酵母(Rhodotorula sp.)CIBASA1401菌株的鑒定[J].應用與環境生物學報,2004(2):242-245.
[6]榮瑞芬,郭堃,厲重先.紫外照射誘導采后番茄苯丙氨酸解氨酶的分離純化[J].北方園藝,2007(12):1-4.
[7]歐陽光察,應初衍,沃少根.植物苯丙氨酸解氨酶的研究:Ⅵ.水稻、小麥PAL的純化及基本特性[J].植物生理與分子生物學學報,1985(2):204-214.
[8]董艷珍.植物苯丙氨酸解氨酶基因的研究進展[J].生物技術通報,2006(S1):21-24.
[9]馬立安,江濤,張忠明.擬南芥Ran2基因的原核表達及產物的純化[J].長江大學學報:自科版農學卷,2007(1):4-6.
[10]宋柬,馬會勤,郝佳.大豆苯丙氨酸解氨酶在大腸桿菌中的重組表達及活性鑒定[J].食品科學,2006(7):29-35.
[11]徐虹,歐陽平凱.深紅酵母轉化反式肉桂酸生成L-苯丙氨酸的研究[J].工業微生物,2000(2):30-33.
[12]Havir E A.Phenylalanine ammonia-lyase:Purification and characterization from soybean cell suspension cultures[J].Archives of Biochemistryand Biophysics,1981(2):556-563.
[13]孫亞莉,郝輝芳,郭夢瑤.水稻耐鹽基因ZFP1及蛋白的生物信息學分析生物技術[J].山西農業科學,2015,43(12):1585-1588,1616.
[14]Zhang X F,Yang F Y.Further study on the role of Mg2+in lipid-protein interaction of the reconstituted porcine heart mitochondrial H+-ATPase[J].Biochim Biophys Acta,1989,976:53-62.
[15]王東,楊金水.棉花類耐鹽鋅指蛋白基因的克隆與結構分析[J].復旦學報:自然科學版,2002(1):46-50.
[16]鄒蔚然,郭萬株.蛋白激酶基因的克隆和生物信息學分析[D].成都:四川農業大學,2008:21-26.
[17]Matus J T,Loyola R,Vega A,et al.Post-veraison sunlight exposure induces MYB-mediated transcriptional regulation of anthocyanin and flavonol synthesis in berryskins ofVitis vinifera[J].Journal of Experimental Botany,2009,60(3):853-867.
[18]王劍利,楊章民.利用生物信息學技術研究蛋白功能的幾種方法[J].國外醫學:分子生物學分冊,2001,23(4):255-256.
[19]陳潤生.生物信息學[J].生物物理學報,1999,15(1):5-6.
[20]李松,王英.生物信息學在生命科學研究中的應用[J].熱帶醫學雜志,2009(10):1218-1221.
Cloning and Expression Analysis of Phenylalanine Ammonia-lyase Gene fromArabidopsis thaliana
ZHAODan
(College ofInformation,Shanxi Agricultural University,Taigu 030800,China)
Phenylalanine ammonia-lyase(PAL)is the first key enzyme involved in the metabolism of phenylpropanoid,which is a widespread metabolic pathwayin plants.Phenylalanine ammonia-lyase(PAL)regulates the expression ofgenes bytranscription initiation, frequency control and response to external factors,and its expression level plays an important role in downstream flavonoid synthesis. Under certain conditions,phenylalanine ammonia-lyase can be used to reverse-catalyze the production ofL-phenylalanine by its special enzymatic properties.In this paper,the presence of phenylalanine ammonia-lyase in Arabidopsis thaliana was determined by a series of bioinformatics analysis,and cloning and expression of the gene analysis,for the production of L-phenylalanine to provide a newspecies source.
Arabidopsis thaliana;phenylalanine ammonia-lyase;bioinformatics;clone
Q943.2
A
1002-2481(2016)12-1767-05
10.3969/j.issn.1002-2481.2016.12.07
2016-10-27
趙丹(1986-),男,山西太原人,助教,主要從事生物化學與分子生物學研究工作。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
