植物NBS類R基因的分類、進化、調控及應用
2017-01-06 03:20:04劉靜暢志堅郭慧娟李欣張曉軍詹海仙任永康喬麟軼
山西農業科學 2016年3期
劉靜,暢志堅,郭慧娟,李欣,張曉軍,詹海仙,任永康,喬麟軼,
(1.山西大學研究生院,山西太原030006;2.山西省農業科學院作物科學研究所,作物遺傳與分子改良山西省重點實驗室,山西太原030031)
植物NBS類R基因的分類、進化、調控及應用
劉靜1,暢志堅2,郭慧娟2,李欣2,張曉軍2,詹海仙2,任永康2,喬麟軼1,2
(1.山西大學研究生院,山西太原030006;2.山西省農業科學院作物科學研究所,作物遺傳與分子改良山西省重點實驗室,山西太原030031)
NBS類抗病基因作為抗病基因中數目最多的家族,在植物抵御病蟲害中起著關鍵作用。綜述了NBS類抗病基因的分類、功能、數目、起源進化、表達調控以及應用等方面的研究進展,并對該類基因的研究前景進行了展望。
NBS;分類;進化;調控;應用
植物在生長過程中不斷面臨著真菌、細菌、病毒、線蟲等病原體的侵襲,而在長期的進化過程中植物也逐漸形成了一系列復雜的防御機制,特別是植物抗病基因編碼的蛋白對病原體的特異性識別。隨著對抗病基因及其編碼產物的結構與功能研究,以及植物信號轉導途徑中其他組分的進一步解讀,人們對有關植物與病原體互作、抗病基因進化的分子機制的研究不斷深入。盡管基因產物針對的病原體及致病因子截然不同,但其植物抗性基因的分子內結構特征相對十分保守,常見的有蛋白激酶(protein kinase,PK)、跨膜結構域(transmembrane domain,TM)和核苷酸結合位點(nucleotide binding site,NBS)等,其中,NBS類基因是植物中普遍存在的抗病基因,在植物抵抗各種病原物侵襲的過程中發揮著重要作用。
1 NBS類抗病基因的分類、功能及數目
NBS結構域是該類基因所編碼蛋白中最保守的序列,存在于真核生物的許多蛋白中(如ATP合酶的亞基、核糖體延伸因子、ras蛋白等),能夠結合ATP或GTP,可能參與抗病信號的傳導[1]。根據NBS類抗病基因編碼蛋白的N端結構不同,可將其分為TIR-NBS-LRR(TNL)和non-TIR-NBS-LRR(non-TNL)兩類。其中,TIR通過與病原物及其產物的結合,產生信號并將信號傳遞給傳導因子,從而引發抗病反應[2];LRR結構域由一系列相似重復的模體構成,主要參與對Avr蛋白的特異性識別[3-4];non-TNL的N端為CC卷曲螺旋結構[2],該結構由2個或2個以上的螺旋相互作用形成一個超級螺旋,分散在疏水氨基酸殘基中,與病原產生的配體結合,進而感知病原侵染。
NBS類抗病基因廣泛存在于植物基因組中,目前,多種植物的NBS類抗病基因的數目已確定。由表1可知,數目最少的為短柄草,只有23個,而小麥高達1 219個,差異較大可能是由基因組大小不同[5]以及在進化過程中病蟲害和環境選擇壓力不同所造成的[6]。
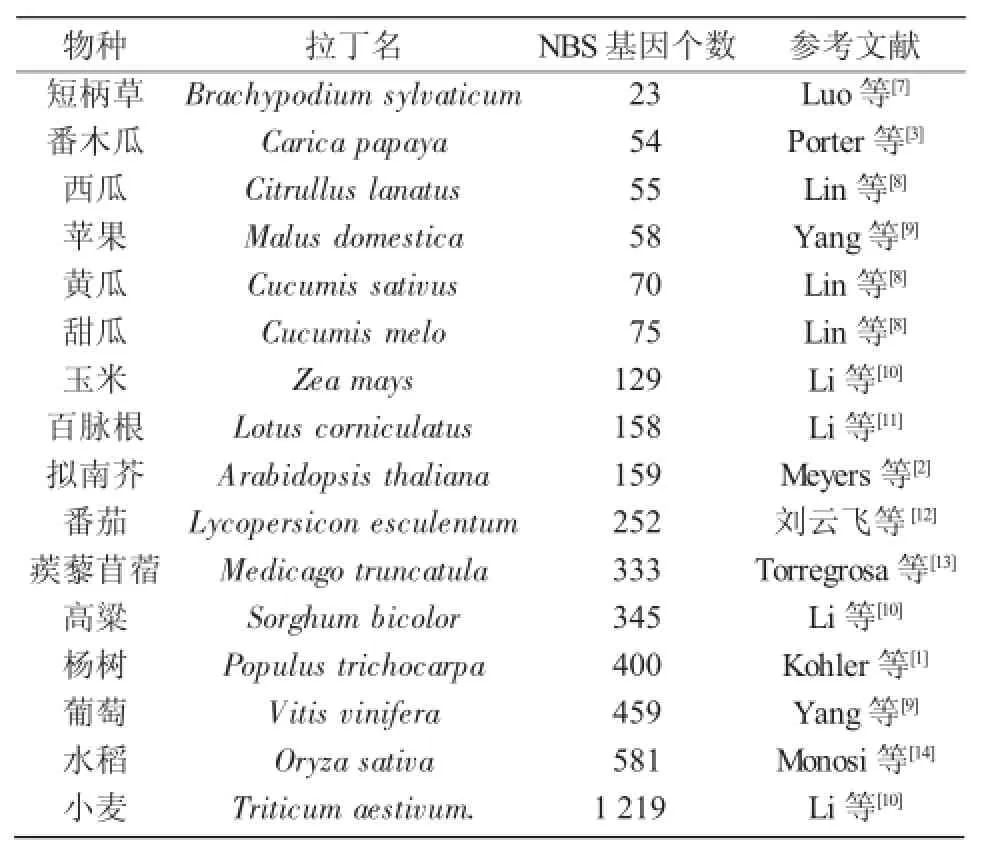
表1 16種植物中NBS類抗病基因數目
2 NBS類抗病基因的起源與進化
許多研究表明,NBS,LRR,TIR,CC結構域在抗病基因發揮作用中必不可少,因此,研究這些結構域的起源是研究抗病基因起源中的最關鍵部分。隨著植物全基因組序列的不斷公布以及基于BLAST的同源搜索和簡并引物PCR的產生,有關結構域起源的研究已取得了一定進展。2002年,Meyers等[15]通過在小立碗蘚的ETS數據庫BLAST搜索發現,其中2個TIR序列在陸生植物中普遍存在。同年,Akita等[16]通過簡并引物PCR法,也發現了TIR類似片段。2012年,Yue等[17]對NBS類抗病基因的起源進行了更深的研究,結果表明,TNL類起源于苔蘚類植物,non-TNL類起源于石松類植物,且NBS,LRR各自在真核生物和原核生物之前已出現,但在陸生植物中才發生融合,隨后擴展到被子植物和裸子植物中。
有研究還發現,NBS結構中既存在保守型區域,也含有多態性的片段,推測NBS基因在進化過程中,一些具有重要功能的結構域被保存下來,而為了適應多變的環境產生了變異。導致真核生物的基因數目及復雜度增加的最重要原因為基因復制[18],其有助于基因網絡的進化,并為新遺傳系統以及基因功能的進化提供了新素材。例如,Martin等[19]對萵苣基因組研究發現了通過染色體復制產生的整套新的抗性基因簇;Pamiske等[20]研究發現,cf9基因至少存在2個同源基因族,暗示cf類型基因是DNA復制的產物;Wang等[21]研究表明,xa21基因家族中含有17 kb的大片段重復,說明有基因參與復制的跡象。
在被子植物和裸子植物中,TNL和non-TNL 2種類型的NBS抗病基因都存在,而在雙子葉植物中只有TNL類型存在。有關數據表明,這2種類型的進化模式不同,Zhong等[22]通過對反義替換Ka與同義替換Ks的比值研究TNL,non-TNL的進化模式,結果發現,不同植物中其比值可能大于1,也可能小于1,即不同植物有不同的生命歷程,其TNL,non-TNL多樣化的進化模式是由其所在的特定生態環境所決定。
3 NBS類抗病基因的表達與調控
植物NBS類抗病基因的表達存在差異性,即在不同物種或同一物種不同組織器官中表達有先后,且表達產物以及表達量有差異。Akita等[16]研究發現,苔蘚植物中,NBS類抗病基因的表達與發育有關,在配子體階段,其表達水平比絲狀體高。宋霄等[23]對MdNBS-LRR1基因的組織表達特異性進行檢測,結果顯示,MdNBS-LRR1在分析的6種組織中均有表達,但在幼葉中的表達量最高。楊慧等[24]也比較了PB-LRR基因在野生型水稻不同器官中的表達模式,結果表明,PB-LRR基因在水稻葉片中的表達水平約是根中的13倍。
植物抗病基因的表達除具有組織特異性外,主要還受環境、生長調節劑、miRNA以及轉錄因子的調控。Wang等[25]對含NBS結構的OsRP1L1基因在高鹽、高低溫、強滲透壓條件下的表達情況進行了研究,其中,高鹽、高溫抑制該基因的表達,而強滲透壓與低溫對其表達影響較小。Que等[26]研究發現,水楊酸對甘蔗NBS基因(SNLR)的表達有一定的影響。Guo等[27]測定了水楊酸(SA)、茉莉酸(JA)、乙烯(TE)等對水稻苗期NBS基因(DEPG1)表達的影響,結果表明,這3種激素均不同程度地抑制DEPG1基因的表達。
病原菌侵染導致抗病基因的瞬時表達,由于表達量過高可能會引起過敏性反應,從而導致細胞的局部死亡,這對植物生長發育非常不利;而植物可通過其內源性抗病毒防御系統的轉錄及轉錄后水平調控引起基因沉默,進而影響基因的表達。He等[28]在白菜中發現了一個特有的miRNA,可以靶向沉默TNL類基因,并證明這個miRNA可以被TuMV誘導表達,使TNL基因被切割降解。2009年,Klevebring等[29]對楊樹基因組中的miRNA進行預測驗證,結果發現了1個新的miRNA,其靶基因為NBS-LRR類抗病基因,可能調控該類基因的表達。當病原菌侵染植物后通過信號傳遞把轉錄因子激活,轉錄因子與抗病基因的啟動子結合,從而引起基因表達。
4 NBS類抗病基因的應用
隨著克隆技術的發展,對抗病基因的結構有了全面的認識,發現這些基因都含有幾個高度保守的結構域,利用其保守性,目前在許多植物中已克隆出抗病基因同源物。Klevebring等[29]利用TIGR和NCBI數據庫,對R基因保守結構域的關鍵詞進行搜索,獲得900條RGA。高樹廣等[30]根據已知植物抗病基因的NBS-LRR類保守結構域,合成1對簡并、2對特異引物,對6個芝麻品種中的抗病基因進行擴增,得到11條抗病基因同源序列。Ilag等[31]利用擬南芥的NBS-LRR抗病基因Rps2作為探針,從水稻IR64中篩選出3個NBS-LRR同源序列NRH1,NRH2和NRH3。張立榮等[32]利用已克隆植物抗病基因NBS序列中的保守結構P-loop和GLPL合成簡并引物,對小麥近等基因系TcLr24基因組DNA進行PCR擴增,得到13條具有連續ORF的抗病基因類似物。利用NBS抗病基因克隆出的RGA通常被應用于分子標記的開發,基于RGA開發的標記主要有RFLP,STS,SSCP,CAPS和SSR等。Liu等[33]從25個花生RGA中開發出28個SSR標記,并將其中一個標記RGA121定位到連鎖群AhIV上。Samuelian等[34]同樣也從9個覆盆子RGA中開發出3個CAPS標記。除以上2種外,通過轉基因技術NBS類抗病基因已在植物抗病育種中得到應用。李海霞等[35]采用農桿菌介導法將從毛白楊中克隆得到的TNL抗病基因(PtDRG01)導入煙草中,經抗生素篩選和PCR分子檢測,獲得一批陽性轉化植株。郭立佳等[36]通過構建NBS-LRR類基因Osxoc1334的超量表達載體,并在農桿菌EHA105介導下轉化中花11號水稻,最終獲得6株再生植株。
5 展望
近年來,隨著生物信息學的快速發展,部分植物的基因組測序相繼完成。目前,已對NBS類抗病基因在全基因組水平上進行了較全面的分析,主要包括基因結構、數量、分布以及與其他植物NBS類基因的進化和親緣關系的比較分析,但是對于NBS類基因的表達調控機制研究還有待進一步深入。對于大部分NBS類抗病基因來說,其只具有專化抗性,隨著病原菌的變異,植物抗病性很容易喪失。因此,發掘和克隆具有廣譜性的抗病基因將是NBS類抗病基因研究的重要內容。
[1]Kohler A,Rinaldi C,Duplessis S,et al.Genome-wide identification of NBS resistance genes in Populus trichocarpa[J].Plant Molecular Biology,2008,66(6):619-636.
[2]Meyers B C,Kozik A,Griego A,et al.Genome-wide analysis of NBS-LRR-encoding genes in Arabidopsis[J].The Plant Cell,2003,15(4):809.
[3]Porter B W,Paidi M,MingR,et al.Genome-wide analysis of Carica papaya reveals a small NBSresistance gene family[J].Molecular Genetics and Genomics,2009,281(6):609-626.
[4]曹必好,宋洪元,雷建軍.植物抗病基因研究進展[J].天津農業科學,2001,7(4):13-18.
[5]Richter T E,Ronald P C.The evolution of disease resistance genes [J].Plant Molecular Biology,2000,42(1):195-204.
[6]Michelmore R W,Meyers B C.Clusters of resistance genes in plants evolve by divergent selection and a birth-and-death process[J]. Genome Research,1998,8(11):1113-1130.
[7]Luo S,Zhang Y,HuQ,et al.Nucleotide-binding site and leucine-rich repeat-encoding genes in the grass family[J].Plant Physiol,2012,159(1):197-210.
[8]Lin X,Zhang Y,Kuang H.Frequent loss of lineages and deficient duplications accounted for low copy number of disease resistance genes in Cucurbitaceae[J].BMCGenomics,2013,14(1):1-13.
[9]Yang S H,Zhang X H,Yue J X,et al.Recent duplications dominate NBS encoding gene expansion in two woody species[J].Molecular Genetics and Genomics,2008,280(3):187-198.
[10]Li J,Ding J,Zhang W,et al.Unique evolutionary pattern of numbers of gramineous NBS-LRR genes[J].Mol Genet Genomics,2010,283(5):427-438.
[11]Li XY,ChengY,Ma W,et al.Identification and characterization of NBS-encoding disease resistance genes in Lotus japonicus[J]. Plant Systemat Evol,2010,289(1/2):101-110.
[12]劉云飛,萬紅建,韋艷萍,等.番茄NBS-LRR抗病基因家族全基因組分析[J].核農學報,2014,28(5):790-799.
[13]Torregrosa C A,Wang B B,O'Bleness MS,et al.Identification and characterization of nucleotide-binding site-leucine-rich repeat genes in the model plant Medicago truncatula[J].Plant Physiology,2008,146(1):5-21.
[14]Monosi B,Wisser R J,Pennill L,et al.Full-genome analysis of resistance gene homologues in rice[J].Theor Appl Genet,2004,109(7):1434-1447.
[15]Meyers B C,Morgante M,Michelmore R W.TIR-X and TIR-NBS proteins:twonewfamiliesrelatedtodiseaseresistance TIR-NBS-LRR proteins encoded in Arabidopsis and other plant genomes[J].BMCResearch Notes,2002,32(1):77-92.
[16]Akita M,Valkonen J P.Anovel gene familyin moss(Physcomitrella patens)shows sequence homology and a phylogenetic relationship with the TIR-NBS class of plant disease resistance genes[J]. Journal ofMolecular Evolution,2002,55(5):595-605.
[17]Yue J X,Meyers B C.Tracingthe origin and evolutionary history of plant nucleotide-binding site-leucine-rich repeat(NBS-LRR)genes[J].NewPhytologist,2012,193(4):1049-1063.
[18]Himmelreich R,Hilbert H,Plagens H,et al.Complete sequence analysis of the genome of the bacterium Mycoplasma pneumoniae [J].Nucleic Acids Res,1996,24(22):4420-4449.
[19]Martin G B,Frary A,Chunwongse J,et al.A member of the tomato Pto gene familyconfers sensitivitytofenthion resultingin rapid cell death[J].Plant Cell,1994,6(11):1543-1552.
[20]Pamiske M,Hammond-Kosack KE,Golsten C,et al.Novel disease resistance specificities result from sequence exchange between tandemly repeated genes at the cf-4/9 locus of tomato[J].Cell,1997,91(6):821-832.
[21]Wang G L,Ruan D L,Song W Y,et al.xa21D encodes a receptor-like molecule with a leucine-rich repeat domain that determines race-specific recognition and is subject toadaptive evolution [J].Plant Cell,1998,10(5):765-779.
[22]Zhong Y,Yin H,Sargent D J,et al.Species-specific duplications driving the recent expansion of NBS-LRR genes in five Rosaceae species[J].BMCGenomics,2015,16(1):1471-2164.
[23]宋霄,柏素花,戴洪義.蘋果NBS-LRR1基因的鑒定與表達分析[J].園藝學報,2013,40(7):1233-1243.
[24]楊慧,薛彥久,王瑩瑩.一個光敏色素B調控的水稻NBS-LRR基因的表達模式分析[J].山東農業科學,2011(6):1-9.
[25]Wang X M,Chen J,Yang Y,et al.Characterization of a novel NBS-LRR gene involved in bacterial blight resistance in rice[J]. Plant Molecular BiologyReporter,2013,31(3):649-656.
[26]Que Y X,Lin J W,Zhang J S,et al.Molecular cloning and characterisation of a non-TIR-NBS-LRR type disease resistance gene analogue fromsugarcane[J].Sugar Tech,2008,10(1):71-73.
[27]Guo L J,Li M,Wang W J,et al.Over-expression in the nucleotide-binding site-leucine rich repeat gene DEPG1 increases susceptibility to bacterial leaf streak disease in transgenic rice plants[J].Molecular BiologyReports,2012,39(4):3491-3504.
[28]He X F,FangY Y,FengL,et al.Characterization of conserved and novel microRNAs and their targets,including a TuMV-induced TIR-NBS-LRR class R gene-derived novel miRNA in Brassica[J]. FEESLett,2008,582(16):2445-2452.
[29]KlevebringD,Street NR,Fahlgren N,et al.Genome-wide profiling ofpopulus small RNAs[J].BMCGenomics,2009,10:620-638.
[30]高樹廣,劉紅彥,王俊美,等.芝麻NBS-LRR類抗病基因同源序列的分離與分析[J].河南農業科學,2011,40(9):81-85.
[31]Ilag L L,Yadav R C,Huang N,et al.Isolation and characterization of disease resistance gene homologues from rice cultivar IR64[J]. Gene,2000,255(2):245-255.
[32]張立榮,楊文香,劉大群.小麥NBS類抗病基因類似序列的多樣性和進化關系研究[J].華北農學報,2011,26(4):23-26.
[33]Liu Z J,Feng S,Pandey MK,et al.Identification of expidentification of expressed resistance gene analogs from peanut(Arachis hypogaea L.)expressed sequence tagss[J].Journal Integrative Plant Biology,2013,55(5):453-461.
[34]Samuelian S K,Baldo A M.Isolation and linkage mapping of NBS-LRR resistance gene analogs in red raspberry(Rubus idaeus L.)and classification among 270 Rosaceae NBS-LRR genes[J]. Tree Genetics&Genomes,2008,4(4):881-896.
[35]李海霞,張志毅,張謙,等.毛白楊TIR-NBS-LRR基因轉化煙草的研究[J].北京林業大學學報,2009,31(1):73-78.
[36]郭立佳,毛倩,汪衛,等.一個NBS-LRR蛋白質基因轉化水稻及其超量表達[J].廈門大學學報:自然科學版,2010,49(4):552-557.
Classification,Evolution,Regulation and Application of NBS R-genes in Plants
LIUJing1,CHANGZhi-jian2,GUOHui-juan2,LI Xin2,ZHANGXiao-jun2,ZHANHai-xian2,RENYong-kang2,QIAOLin-yi1,2
(1.College ofGraduate,Shanxi University,Taiyuan 030006,China;2.Shanxi Province KeyLaboratoryofCrop Genetics and Molecular Improvement,Institute ofCrop Sciences,Shanxi AcademyofAgricultural Sciences,Taiyuan 030031,China)
Nucleotide binding site(NBS)-encoding gene family,which contains the largest number of members,plays a critical role in plant resistance to pathogens and pests.In this review,we summarized the research progress of classification,function,number, evolution,expression and application ofNBSgenes in plant,and discussed the prospect ofresearch on them.
NBS;classification;evolution;regulation;application
Q943.2
A
1002-2481(2016)03-0423-04
10.3969/j.issn.1002-2481.2016.03.37
2015-10-20
山西省科技攻關項目(20150311001-1);山西省青年基金項目(2015021145);山西省農業科學院攻關項目(15YGG01);山西省農業科學院重點科研項目(YZD1501)
劉靜(1991-),女,山西呂梁人,在讀碩士,研究方向:種質資源創新與利用。喬麟軼為通信作者。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
