基于遺傳粒子群算法的番茄幼苗光合優化調控模型
2017-01-12 08:13:16王智永辛萍萍張佐經
上海農業學報 2016年6期
關鍵詞:模型
王 東,王智永,裴 雪,辛萍萍,張佐經
(西北農林科技大學機械與電子工程學院,楊凌 712100)
基于遺傳粒子群算法的番茄幼苗光合優化調控模型
王 東,王智永,裴 雪,辛萍萍,張佐經*
(西北農林科技大學機械與電子工程學院,楊凌 712100)
番茄光合速率主要受溫度和光子通量密度影響,動態獲取不同溫度條件下的光飽和點信息是提高光環境調控效率的關鍵。該文結合遺傳粒子算法提出了一種光合優化調控模型:利用光合速率雙因素嵌套試驗獲取多維數據,構建溫度、光子通量密度耦合的光合速率多元非線性回歸模型,采用遺傳算法對光合速率模型進行優化,獲取任意離散溫度值下的光飽和點,以飽和光照強度作為目標值建立光合優化調控模型。以番茄幼苗為例進行了驗證,試驗結果表明:提出的方法可動態獲取不同溫度條件下光飽和點,光飽和點實測值與計算值決定系數為0.9873,最大相對誤差小于4.6%,具有較高精度,對提高設施光環境調控效率具有重要的意義。
番茄;幼苗;光合速率;光飽和點;遺傳算法;粒子群算法;調控模型
番茄是我國的主要設施作物之一,每年總產量世界領先[1]。番茄作為光敏感作物,光照不足會造成其產量大幅下降[2]。光合作用是描述作物有機物質積累的過程,不僅受光照影響,且受溫度、二氧化碳濃度等環境因子影響[3-5],為實現設施番茄光環境高效調控,提高其產量與品質,番茄光合速率研究廣泛開展。其中蔣燕等[6]、張富存等[7]研究表明溫度過低或過高均會導致番茄幼苗光合速率降低,說明溫度和光照是影響光合速率因素中的兩個主要因素[8];胡瑾等[9]采用遺傳算法進行光溫匹配尋優,為本試驗設計有一定借鑒作用。因此,探究溫度、光照環境信息與光合速率間對應關系,尋找不同環境下使番茄達到最大光合速率的飽和光強,建立番茄光合優化調控模型,是提高番茄產量的重要途徑。
為實現基于作物需求的光合最優調控,光合速率模型作為其理論基礎得到廣泛研究。其中李天來等[10]研究建立了基于溫度修正的番茄光合速率模型;賴琳玲等[11]研究發現隨溫度、光照變化番茄生長的變化規律;以色列、美國等科學家合作開發了TOMGRO溫室番茄模型;荷蘭科研人員開發的HORTSIM作物模擬器是目前該領域的典型代表[12-13],以上研究均建立了溫度與光子通量密度關聯的光合速率模型,為本試驗研究提供了依據,但真正的按需調控應建立在不同環境下光飽和點擬合的基礎上,而上述模型未實現光飽和點尋優,因此構建番茄光合速率模型,設計尋優算法獲取光飽和點,是建立光合調控模型的關鍵。
遺傳算法與粒子算法是實現高效精準尋優的常用手段。目前,在遺傳粒子群算法上,國內外學者也做了不少的研究。其中蔡雁等[14]采用遺傳粒子尋優算法獲取最優鐵礦粉庫存量,為鋼鐵企業的科學采購提供依據;張干清等[15]基于雜交GA-PSO算法的結構協同優化,結果表明提出的雜交GA-PSO協同算法較GA與PSO算法有更快的收斂速度,當相關系數為0.7時,可使該行星架的總體積減小7.06%;陳雙等[16]提出了車輛主動懸架的遺傳粒子群LQG控制方法,其結果表明車輛主動懸架的遺傳粒子群LQG控制方法;Abd-El-Wahed等[17]采用集成粒子群算法與遺傳算法對非線性問題進行了優化;Ricardo等[18]提出了一個混合型GA-PSO優化方法的模糊邏輯控制器(FLC)的自動設計,以盡量減少植物反應的穩態誤差,仿真結果表明了該優化方法對控制應用的可行性;Lee等[19]在數值約束和網絡分解的基礎上使用混合GA-PSO對基因調控網絡進行推斷,結果表明了該方法的有效性。以上研究可為最優需光調控模型建立提供依據,但光飽和點尋優易受算法參數、適應度函數、尋優范圍等條件影響,造成較大誤差。
為此,本研究結合遺傳算法和粒子群算法的的優點,以溫光耦合的番茄光合速率模型為基礎,采用遺傳粒子群算法對光飽和點進行尋優,得到各溫度值下的最優解,進一步建立以飽和光強為目標的番茄光合優化調控模型,為番茄光環境調控提供有效手段。
1 材料與方法
1.1 試驗材料
試驗于2015年3—6月在西北農林科技大學南校區玻璃溫室內進行。對象為處于幼苗期的番茄,品種為‘毛刺802’,種植數量為200株。
1.2 試驗方法
選取定植后植株生長狀況差異較小、健壯的番茄幼苗進行試驗,測量設備為Li-6400XT型便攜式光合儀,該光合儀可按需控制測量葉室的小環境,以設置不同試驗條件。試驗時選擇9:00—11:30和14:30—17:30兩個時間段,以避免“午休現象”;設定二氧化碳濃度為300 μmol·mol-1;6個溫度梯度為16℃、21℃、25℃、29℃、33℃、37℃;10個光子通量密度梯度為0 μmol·m-2·s-1、50 μmol·m-2·s-1、100 μmol·m-2·s-1、200 μmol·m-2·s-1、400 μmol·m-2·s-1、600 μmol·m-2·s-1、800 μmol·m-2·s-1、1 000 μmol·m-2·s-1、1 200 μmol·m-2·s-1、1 500 μmol·m-2·s-1。在60組試驗條件中,每組試驗均選取6株幼苗重復測量6次凈光合速率值,共形成360組樣本數據。
1.3 試驗數據處理
按狄克松準則[20-21]對測量數據進行預處理,完成粗大誤差剔除,該準則判斷粗大誤差從最大抽樣值和最小抽樣值入手。在測量中,對同一指標進行多次等精度重復測量,降低測量誤差。
根據狄克松準則對本試驗進行粗大誤差濾除后,測得剩余樣本的平均值如表1所示。通過計算,可得到該30組數據的方差小于0.27,由此表明采用狄克松準則進行粗大誤差分析及濾除的效果較好。
2 光合作用優化調控模型建立
分三步建立調控優化模型:首先根據處理后的數據利用非線性擬合方法建立3元番茄凈光合速率模型以得到尋優目標函數,然后采用遺傳-粒子群組合智能算法對光合速率進行尋優,最終依據尋優得到的光飽和點建立光合優化調控模型。

表1 試驗所得光合速率值Table 1 Photosynthetic rate values obtained from testsμmol CO2·m-2·s-1
2.1 多因子耦合光合速率模型
經過分析,綜合考慮擬合精度及計算復雜性,確定采用3元5次非線性方程進行回歸分析,建立以溫度、光子通量密度為自變量的光合速率模型,如式(1)所示,擬合結果如圖1。
式中,T:溫度/℃;PFD:光子通量密度/μmol·m-2·s-1;Pn:光合速率/μmol CO2·m-2·s-1。
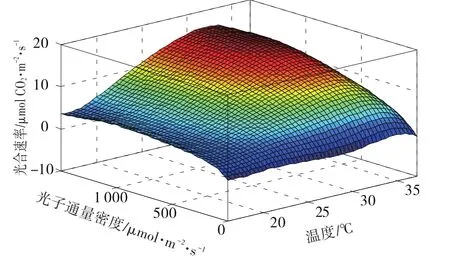
圖1 光合速率擬合圖Fig.1 Fitting of photosynthetic rates
在相同溫度條件下,光合速率隨著光子通量密度增大而增加,在最大值附近趨于變化平緩,但光子通量密度進一步增大時,光合速率會減小;當光子通量密度恒定不變時,隨著溫度的升高光合速率也呈現相似的先升后降的趨勢。以上結果與前人研究結果相符[22-23],且具有良好的擬合效果,說明試驗方案合理。為充分考慮各因子之間的交互作用,構建非線性回歸凈光合速率模型,模型擬合結果的決定系數為0.9929,相比于現有模型,所構建的光合速率模型具有更好的擬合結果。
2.2 基于遺傳粒子群算法的光溫耦合尋優算法
遺傳算法是借鑒生物界適者生存法則的進化算法,被用于解決尋優問題。遺傳算法不容易陷入局部最優,同時適應大規模的并行計算。算法通過復制、交叉、突變等操作產生下一代的解,適應度低的解將被淘汰掉,將適應度高的解保存下來。遺傳算法的優越性主要表現在:具有群體搜索和內在啟發式隨機搜索的特性,不易陷入局部最優;遺傳算法固有的并行性和并行計算的能力,非常適用于大規模并行計算。本研究在粒子群算法中引入遺傳算法的變異算子來增強粒子群算法跳出局部最小值的能力。同時通過引入種群分割策略來維護種群的多樣性,使得各子種群分別進化,子種群之間的相互競爭避免了算法進化過程中陷入局部最優解。
本研究以番茄幼苗不同溫光環境的光合速率值構建樣本數據,建立光合速率模型,設計遺傳粒子尋優算法對光合速率模型進行尋優,獲取最優光合速率解及其對應的飽和光強,具體流程如圖2。
GA-PSO具體算法步驟如下:
1)基于多因子耦合光合速率模型,確定該種群規模為20,由于在該試驗中,個體為溫度與光子通量密度,所以設置個體長度為2,根據經驗,進化代數設為100較為合適。
2)由于本試驗采用了遺傳算法的交叉和變異算子,需要設定其概率參數。不同的交叉概率和變異概率尋優結果會產生較大差異,交叉概率越大,可越快到達最優目標值區域,變異概率過大影響尋優過程的穩定性,因此本文選取交叉概率為0.7,變異概率為0.01。又因為本試驗還采用了粒子群算法,所以還需要設置粒子的位置和速度。溫度粒子是特定的,無需設置;由實際測試得到的光子通量密度可知,其最小值不可能為負,所以設定其下界為0,其最大值不超過1 300,所以設定1 300為其上界,最終設置光子通量密度的初始位置范圍為0—1 300;又根據對粒子群算法的學習,粒子飛行的極限方向是正反兩個方向,因此設光子通量密度的速度范圍為-800—1 000。
3)在特定溫度條件下進行尋優,以2℃為步長構建尋優的溫度樣本集T=(T1,T2,…,Tn),其中Tn=Tmin+2n,Tmin代表特定最小溫度值,n=1,2,3,…。并以此樣本集中數據完成對式(1)所示Pn=f(T,PFD)的溫度實例化。在此基礎上,并基于上文的光合速率模型Pn=f(T,PFD)確定適應度函數式(2):

式中:F表示特定溫度值Tn下的適應度函數。
4)利用rand函數隨機產生初始光子通量密度粒子及其速度。由于rand函數產生隨機數的范圍在0—1之間,基于實際測試結果,我們用位置系數1 500和速度系數1 000調整光子通量密度粒子的位置范圍和速度范圍分別為0—1 500和-1 000—1 000。
5)利用式(2)計算初始化種群的適應度值,即特定溫度下的光合速率值。找出特定溫度下局部范圍內的最大光合速率值,即個體最優位置pbest;再找出特定溫度下全部范圍內的最大光合速率值,即全局最優位置gbest。
6)接下來分別采用式(3)和(4)對光子通量密度粒子的位置和速度進行更新,以產生新的粒子;

式中:v(t)表示進化次數為t時光子通量密度的速度,PFD(t)表示進化次數為t時光子通量密度的位置,Fpbest(t)表示進化次數為t時局部最大光合速率值,Fgbest(t)表示進化次數為t時全局最大光合速率值,c1和c2為學習因子,根據經驗,均設置為1.49445。
7)采用遺傳算子對粒子執行交叉和變異操作。交叉操作如下:


圖2 GA-PSO流程圖Fig.2 Flowchart of GA-PSO
式中,akj為第k個染色體的第j個基因,aij為第i個染色體的第j個基因,b是0—1區間的隨機數。采用如下變異操作過程:

式中,amax是基因aij的上界;amin是基因aij的下界;r為0—1區間的隨機數,g是當前迭代次數,Gmax是最大進化次數。
8)計算經過上述操作的個體的適應度值,并和個體最優pbest比較;若當前個體的適應度值比pbest大,則把該值作為新的個體最優位置,并接著與全局最優gbest比較;若仍比gbest大,則把該值作為新的全局最優位置,并判斷是否滿足最大進化次數100次的這個終止條件,若滿足,則輸出結果。若并未滿足上述條件,則對粒子不斷地進行位置和速度更新以及交叉變異操作,直至滿足條件為止。
9)選取新一組溫度重復步驟3—8,直至完成所有溫度梯度下的尋優,結果如圖3。GA-PSO尋優算法的具體參數如表2所示。

表2 遺傳粒子群算法參數設置Table 2 Parameter setting of GA-PSO

圖3 尋優進化圖Fig.3 Evolutionary optimization figure
按上述步驟1—9分別對18℃,20℃,22℃,24℃,26℃,28℃,30℃,32℃,34℃等9個溫度梯度進行光飽和點尋優,繪制光合速率隨進化代數的變化曲線,如圖3所示。
整體分析圖3可以發現,該9組最佳溫度與光照強度組合下所得到的最佳光合速率值的進化過程很穩定,無振蕩,番茄最佳光合速率值基本保持不變,由此表明算法參數設置較合理。從進化過程來看,當進化代數較小時,最佳光合速率值緩慢增長,當進化至第十代時,最佳光合速率值基本保持恒定。在算法運行的過程中,不斷實現個體進化,獲取適應度高的種群個體,當接近最優解時產生的新的最優個體適應度值上升幅度不明顯,最終趨于穩定。
3 光合優化調控模型構建
根據GA-PSO混合尋優結果繪制離散溫度下光飽和點變化曲線,如圖4所示。光飽和點隨溫度增加呈現先上升后下降趨勢,與作物光合作用變化規律相吻合。
選取溫度T為輸入量,飽和光照強度PFD為輸出量,通過非線性回歸方法進行擬合,建立番茄幼苗光合最優調控模型,如式(7)所示,基于該模型,可獲取任意溫度條件下光飽和點數據。

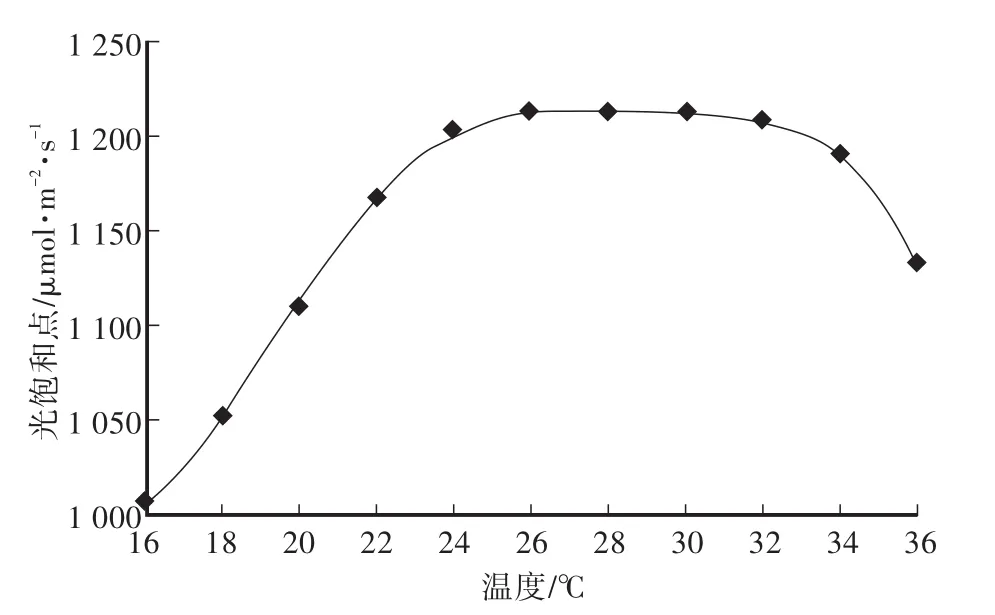
圖4 離散溫度下光飽和點變化曲線Fig.4 Variation of light saturation points at discrete temperatures
4 模型驗證分析與對比
為評價調控模型的準確性與可靠性,對光飽和點計算值與實測值進行對比驗證。使用Li-6400XT型便攜式光合儀,測得16—36℃范圍內13個溫度下的光飽和點,以基于GA-PSO尋優結果建立的需光模型計算得到的13個溫度值下光飽和點作為模擬值,實測值與模擬值二者之間的相關關系如圖5-C所示。為了進一步說明模型的優劣,采用同樣的方法基于GA、PSO尋優結果建立需光模型并驗證實測值與模型模擬值二者之間的相關關系,如圖5-A、5-B所示。
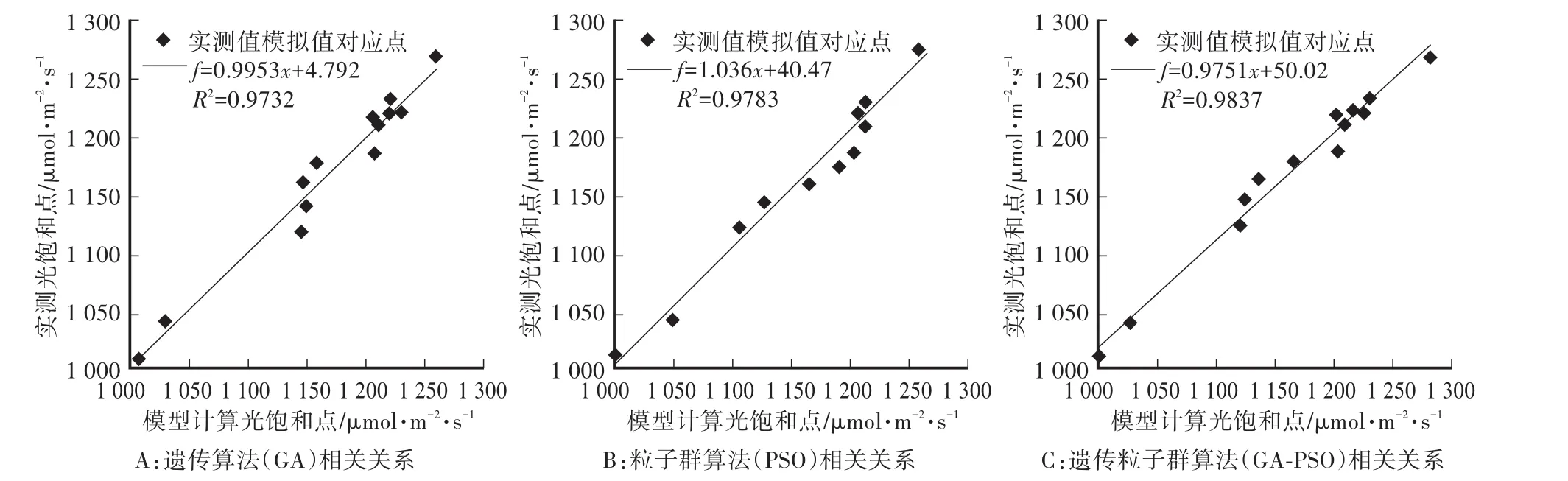
圖5 光飽和點實測值與模擬值的相關性Fig.5 Correlation between the measured and simulated values of light saturation points
通過對圖5所示光飽和點實測值與模擬值的相關性擬合可以得出,基于GA、PSO、GA-PSO三種尋優結果建立的需光模型,得到的決定系數分別為0.9732、0.9783、0.9837,接近于1,均方根誤差分別為12.95、11.16、10.09,可以得出3種基于尋優算法的需光模型效果都較好,其中GA-PSO的效果最優。為了進一步說明模型的準確性,對13組試驗結果進行光飽和點驗證分析,其最大相對誤差不超過4.6%(表3)。

表3 光飽和點誤差分析Table 3 Error analysis of light saturation points
根據表3驗證分析可知,本文所建立的需光調控模型具有較高的精度。說明采用GA-PSO混合算法尋優比采用GA和PSO算法更準確。
5 結論
本文針對番茄光飽和點隨溫度動態變化的特點,將遺傳算法(GA)和粒子群算法(PSO)相結合得到遺傳粒子群算法,采用遺傳粒子群算法(GA-PSO)對光溫進行尋優,獲取不同溫度下光飽和點數據,以其為基礎建立光合優化調控模型,利用番茄幼苗為例進行驗證,具體結論如下:
(1)設計的嵌套試驗,獲取不同溫度、光照對應的光合速率值,以其為樣本數據構建番茄幼苗期光合速率模型。經驗證,模型決定系數R2為0.9925,具有較強的泛化能力,可為光合速率預測提供有效手段。
(2)采用遺傳算法和粒子算法對番茄幼苗光合速率模型進行尋優,得到離散溫度值對應的光飽和點數據,以其為理論基礎建立光合優化調控模型,利用試驗數據對調控模型輸出的光飽和點進行驗證,結果表明,光飽和點計算值與實測值決定系數達0.9837,最大相對誤差不超過4.6%,具有較高的擬合精度。
(3)本文提出的基于遺傳粒子的光合優化調控模型,可實現以光飽和點為目標值的按需調控,從而克服不同作物的光合特性差異,為設施光環境高效調控提供理論基礎。
[1]張春奇,查素娥,李紅波.番茄育種研究概況及展望[J].農業科技通訊,2011(3):29-33.
[2]吳鍇,張靜,郝稱意.溫室番茄群體光合作用模擬模型的研究[J].山西農業大學學報(自然科學版),2013,33(3):225-229.
[3]王銘,李濤,李愛芬,等.光照、溫度和pH對雨生紅球藻光合特性的影響[J].水生生物學報,2009,33(3):400-405.
[4]郭尚,王宇楠.不同光照強度西瓜光合速率日變化的研究[J].華北農學報,2011,26(5):170-173.
[5]張治安,徐克章,陳展宇,等.影響人參葉片光合速率對CO2水平響應的因素[J].南京農業大學學報,2006,29(2):11-14.
[6]蔣燕,趙會杰.低溫弱光處理對番茄幼苗生長的影響[J].河南農業科學,2006(1):87-91.
[7]張富存,張波,王琴,等.高溫脅迫對設施番茄光合作用特性的影響[J].中國農學通報,2011,27(28):211-216.
[8]趙玉萍,鄒志榮,楊振超,等.不同溫度和光照對溫室番茄光合作用及果實品質的影響[J].西北農林科技大學學報(自然科學版),2010(5):125-130.
[9]胡瑾,何東健,任靜,等.基于遺傳算法的番茄幼苗光合作用優化調控模型[J].農業工程學報,2014,30(17):220-227.
[10]李天來,顏阿丹,羅新蘭,等.日光溫室番茄單葉凈光合速率模型的溫度修正[J].農業工程學報,2010,26(9):274-279.
[11]賴琳玲,程智慧,陳學進,等.番茄器官生長動態與環境溫度和光照關系的模擬[J].西北農業學報,2010,19(2):127-132.
[12]DAYAN E,VAN KEULEN H,JONES J W,et al.Development,calibration and validation of a greenhouse tomato growth model:I.Description of the model[J].Agricultural Systems,1993,43(2):145-163.
[13]GIJZEN H,HEUVELINK E,CHALLA H,et al.HORTISIM:A model for greenhouse crops and greenhouse climate[J].Acta Horticulturae,1998,456:441-450.
[14]蔡雁,鐘茜怡,吳敏,等.基于GA-PSO算法的燒結料場原料庫存量優化[J].化工學報,2012,63(9):2824-2830.
[15]張干清,龔憲生.變量相關情況下基于雜交GA-PSO算法的結構協同優化[J].機械工程學報,2012,48(15):113-125.
[16]陳雙,宗長富.車輛主動懸架的遺傳粒子群LQG控制方法[J].汽車工程,2015(2):189-193.
[17]ABD-EI-WAHED W F,MOUSA A A,EL-SHORBAGY M A.Integrating particle swarm optimization with genetic algorithms for solving nonlinear optimization problems[J].Journal of Computational and Applied Mathematics,2011,235(5):1446-1453.
[18]RICARDO M S,OSCAR C,AGUILAR L T.Type-1 and Type-2 fuzzy logic controller design using a hybrid PSO-GA optimization method[J]. Information Sciences,2014,285:35-49.
[19]LEE W P,HSIAO Y T.Inferring gene regulatory networks using a hybrid GA-PSO approach with numerical constraints and network decomposition[J].Information Sciences,2012,188:80-99.
[20]范滿紅,馬勝前,陳彥,等.基于多傳感器數據融合的溫濕度監測系統[J].壓電與聲光,2012,34(3):459-462,465.
[21]劉志平.檢測儀表中粗大誤差的剔除分析[J].電子測量技術,2009,32(11):55-57,84.
[22]張潔,李天來,徐晶.晝間亞高溫處理時期對日光溫室番茄光合作用與產量的影響[J].農業工程學報,2008,24(3):193-197.
[23]張潔,李天來.日光溫室亞高溫對番茄光合作用及葉綠體超微結構的影響[J].園藝學報,2005,32(4):614-619.
(責任編輯:程智強)
A regulatory model of tomato seedlings’photosynthesis based on genetic algorithm-particle swarm optimization
WANG Dong,WANG Zhi-yong,PEI Xue,XIN Ping-ping,ZHANG Zuo-jing*
(College of Mechanical and Electronic Engineering,Northwest A&F University,Yangling 712100,China)
Tomato plants’photosynthetic rate is mainly influenced by temperature and photon flux density,and acquisition of dynamic information of light saturation points at different temperatures is the crux of improving the regulating efficiency of light environment.According to genetic algorithm-particle swarm optimization(GAPSO)the paper proposes a regulatory model of photosynthetic optimization:multidimensional data are acquired by means of the two-factor nested tests of photosynthetic rate,a multivariate nonlinear regression model of photosynthetic rate coupling temperature and photon flux density is built and then optimized by using GA-PSO,thus acquiring the light saturation point at any discrete temperature,and lastly with the saturated light intensity as a desired value,a regulatory model of photosynthetic optimization is established.The model is verified by taking tomato seedlings,and the results show that the light saturation points at different temperatures can dynamically be acquired by the proposed method,the determination coefficient between the light saturation points’measured and calculated values is 0.9873,and the maximum relative error is less than 4.6%,indicating that the proposed method has a high precision and an important significance of improving the regulating efficiency of light environment.
Tomato;Seedling;Photosynthetic rate;Light saturation point;Genetic algorithm;Particle swarm optimization;Regulatory model
S641.2
A
1000-3924(2016)06-026-07
2016-10-11
國家自然科學基金資助項目(31501224);陜西省農業科技創新與攻關項目(2016NY-125)
王東(1982—),男,碩士,講師,研究方向:計算機在農業中的應用。Tel:13991838234,E-mail:wangdong510@163.com
*通信作者:張佐經(1982—),男,講師,主要從事無線傳感器網絡、智能檢測與控制等方面研究。E-mail:zhangzuojing@126.com
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網絡安全與數據管理(2022年1期)2022-08-29 03:15:20
導航定位學報(2022年4期)2022-08-15 08:27:00
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:36
成都醫學院學報(2021年2期)2021-07-19 08:35:14
新世紀智能(數學備考)(2020年9期)2021-01-04 00:25:14
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
