小鼠FABP4基因啟動(dòng)子驅(qū)動(dòng)紅色熒光蛋白載體的構(gòu)建及在牛體細(xì)胞中的表達(dá)
2017-01-17 09:57:02岳永莉于海泉
華北農(nóng)學(xué)報(bào) 2016年6期
關(guān)鍵詞:小鼠
岳永莉,于海泉
(內(nèi)蒙古大學(xué) 哺乳動(dòng)物生殖生物學(xué)及生物技術(shù)教育部重點(diǎn)實(shí)驗(yàn)室,內(nèi)蒙古 呼和浩特 010021)
小鼠FABP4基因啟動(dòng)子驅(qū)動(dòng)紅色熒光蛋白載體的
構(gòu)建及在牛體細(xì)胞中的表達(dá)
岳永莉,于海泉
(內(nèi)蒙古大學(xué) 哺乳動(dòng)物生殖生物學(xué)及生物技術(shù)教育部重點(diǎn)實(shí)驗(yàn)室,內(nèi)蒙古 呼和浩特 010021)
小鼠脂肪型脂肪酸結(jié)合蛋白(FABP4)的啟動(dòng)子/增強(qiáng)子元件是脂肪組織特異性啟動(dòng)元件,為了鑒定該元件在牛體細(xì)胞中的啟動(dòng)效果,首先以小鼠肝臟DNA為模板,通過(guò)PCR克隆得到5.9 kb的FABP4基因啟動(dòng)子片段,連入pMD19-T載體后進(jìn)行酶切及測(cè)序鑒定,經(jīng)EcoT 22I酶切去除非核心啟動(dòng)子區(qū)后精簡(jiǎn)為2.3 kb的片段,通過(guò)SacⅡ酶切將5.9 kb片段和2.3 kb片段的啟動(dòng)子連入紅色熒光蛋白載體pDs-Red 2-1的多克隆位點(diǎn),構(gòu)建為表達(dá)質(zhì)粒pMF5.9-Red和pMF2.3-Red,利用脂質(zhì)體法將上述載體分別轉(zhuǎn)染牛脂肪間充質(zhì)干細(xì)胞及牛胎兒成纖維細(xì)胞,24 h后實(shí)時(shí)定量PCR檢測(cè)紅色熒光蛋白轉(zhuǎn)錄水平。結(jié)果表明,克隆得到的5.9 kb小鼠FABP4啟動(dòng)子片段酶切及測(cè)序結(jié)果正確,與紅色熒光蛋白載體相連的載體pMF5.9-Red和pMF2.3-Red酶切結(jié)果與預(yù)期相符,表達(dá)載體構(gòu)建成功,實(shí)時(shí)定量PCR結(jié)果顯示以上2種細(xì)胞在轉(zhuǎn)染后24 h紅色熒光蛋白均有表達(dá),且pMF2.3-Red的轉(zhuǎn)錄水平是pMF5.9-Red的2倍以上。成功構(gòu)建了小鼠FABP4啟動(dòng)子驅(qū)動(dòng)紅色熒光蛋白表達(dá)載體pMF5.9-Red和pMF2.3-Red,5.9 kb片段和2.3 kb片段均可驅(qū)動(dòng)外源基因在牛體細(xì)胞中轉(zhuǎn)錄,且2.3 kb片段啟動(dòng)效率高于5.9 kb片段。
小鼠FABP4啟動(dòng)子;載體構(gòu)建;牛體細(xì)胞
用組織特異性啟動(dòng)子與增強(qiáng)子作為轉(zhuǎn)錄啟始元件調(diào)控目的基因的表達(dá),是動(dòng)物基因工程中限定外源基因時(shí)空特異性表達(dá)常用的手段,在應(yīng)用上比較成熟的如乳腺特異型(酪蛋白、乳球蛋白)、肌肉特異型(肌動(dòng)蛋白、肌酸激酶、生肌因子家族)、肝臟特異型(TTR)、生殖細(xì)胞特異型(Oct-4)、毛囊特異型(超高硫角蛋白)等啟動(dòng)子,已經(jīng)有許多成功獲得轉(zhuǎn)基因動(dòng)物的報(bào)道[1]。
脂肪組織是機(jī)體中重要的能量?jī)?chǔ)存器官,承擔(dān)著維持體溫、保證代謝的正常運(yùn)轉(zhuǎn)和保護(hù)身體及內(nèi)臟器官免受機(jī)械沖擊傷害的功能,還是機(jī)體重要的內(nèi)分泌器官,通過(guò)分泌激素和細(xì)胞因子發(fā)揮作用。小鼠脂肪型脂肪酸結(jié)合蛋白(Fatty acid binding protein 4,F(xiàn)ABP4又稱為Adipocyte protein-2,aP2)啟動(dòng)子作為脂肪組織的特異性啟動(dòng)子,與Cre-loxP系統(tǒng)的成功結(jié)合,是脂肪細(xì)胞特異性基因敲除小鼠的技術(shù)基礎(chǔ)[2-4]。由于脂肪組織在胚胎發(fā)育的后期開始出現(xiàn),對(duì)于胚胎發(fā)育以及新生兒的存活具有重要意義,因此,對(duì)于基因的胚胎致死問(wèn)題則可以通過(guò)aP2-CreERT2條件敲除小鼠品系來(lái)解決[5]。FABP4啟動(dòng)子元件已成功應(yīng)用于胰島素耐受[6]、肥胖[7]、肝脂肪變性[8]等疾病治療及相關(guān)基因的研究。
本研究以紅色熒光蛋白作為報(bào)告基因,對(duì)比分析了小鼠FABP4啟動(dòng)子元件 5.9 kb全長(zhǎng)和酶切后的2.3 kb片段在牛的脂肪間充質(zhì)干細(xì)胞和牛胎兒成纖維細(xì)胞中的啟動(dòng)效率,以期為下一步利用該元件制作轉(zhuǎn)基因牛奠定工作基礎(chǔ)。
1 材料和方法
1.1 試驗(yàn)材料
清潔級(jí)昆明白小鼠,由內(nèi)蒙古大學(xué)實(shí)驗(yàn)動(dòng)物研究中心提供。DNA提取試劑盒、pMD19-T載體、限制性內(nèi)切酶、RNA提取試劑盒、cDNA合成試劑盒、實(shí)時(shí)定量PCR試劑盒為寶生物工程(大連)有限公司提供,脂質(zhì)體LTX為Invitrogen公司產(chǎn)品,HifiTransTaq酶為北京全式金生物技術(shù)有限公司產(chǎn)品。牛脂肪間充質(zhì)干細(xì)胞、牛胎兒成纖維細(xì)胞為本實(shí)驗(yàn)室保存。
1.2 載體構(gòu)建
提取6周齡小鼠肝臟的基因組DNA,利用在兩端添加了SacⅡ酶切位點(diǎn)的引物F:5′-CCGCGGAACCTGCTAACGAACTTGACCAT-3′,R:5′-CCGCGGTCCCACAAAGGCATCACACAT-3′進(jìn)行PCR擴(kuò)增,反應(yīng)程序?yàn)?4 ℃預(yù)變性3 min;94 ℃變性30 s,60 ℃復(fù)性30 s,72 ℃延伸6 min,32個(gè)循環(huán);72 ℃延伸10 min。PCR產(chǎn)物進(jìn)行電泳、膠回收后,與T載體連接,該載體命名為pMF5.9,將pMF5.9載體送寶生物工程(大連)有限公司測(cè)序,同時(shí)進(jìn)行酶切鑒定。將pMF5.9進(jìn)行EcoT 22I酶切后回收5.0 kb片段進(jìn)行連接,連接后的載體命名為pMF2.3。分別將pMF5.9和pMF2.3用SacⅡ酶切后,分別回收5.9,2.3 kb片段,與同樣經(jīng)SacⅡ酶切的pDsRed2-1載體相連,分別命名為pMF5.9-Red和pMF2.3-Red載體(圖1)。
1.3 細(xì)胞轉(zhuǎn)染及檢測(cè)
將第5代的牛脂肪間充質(zhì)干細(xì)胞和牛胎兒成纖維細(xì)胞以2×105個(gè)/孔的密度接種于6孔板,根據(jù)脂質(zhì)體說(shuō)明書采用2.5 μg質(zhì)粒DNA+10 μL LTX的條件轉(zhuǎn)染細(xì)胞,24 h后收集細(xì)胞,提取總RNA,反轉(zhuǎn)錄后進(jìn)行實(shí)時(shí)定量PCR檢測(cè),反應(yīng)程序?yàn)?5 ℃ 30 s,然后是95 ℃ 5 s,60 ℃ 30 s,40個(gè)循環(huán)。內(nèi)參基因GAPDH的引物為F:5′-GAPDHTTCAACGGCACAGTCAAG
G-3′,R:5′-ACATACTCAGCACCAGCATCAC-3′,紅色熒光蛋白引物為F:5′-CATCCCCGACTACAAG AAGC-3′,R:5′-TGGTCTTCTTCTGCATCACG-3′。
2 結(jié)果與分析
2.1 小鼠FABP4啟動(dòng)子擴(kuò)增
以小鼠肝臟基因組DNA 為模板,成功克隆了全長(zhǎng)5.9 kb 的小鼠FABP4啟動(dòng)子/增強(qiáng)子(圖2)。經(jīng)測(cè)序得知,克隆片段與GenBank公布的序列同源性為99.3%。經(jīng)序列分析得知,轉(zhuǎn)錄起始位點(diǎn)ATG上游的-311,-1 905,-3 096,-3 476,-3 933 bp位點(diǎn)處存在EcoT 22I酶切位點(diǎn)。
2.2 載體構(gòu)建
載體構(gòu)建流程見圖1。將2.1中的PCR片段與T載體連接后,提取質(zhì)粒分別經(jīng)EcoR Ⅰ與EcoR Ⅴ酶切鑒定,電泳條帶符合預(yù)期,證明pMF5.9質(zhì)粒構(gòu)建成功。將質(zhì)粒pMF5.9經(jīng)EcoT 22I酶切后,回收5.0 kb片段進(jìn)行連接,將連接后載體分別經(jīng)Hind Ⅲ與EcoR Ⅰ單酶切鑒定,電泳條帶符合預(yù)期,證明pMF2.3質(zhì)粒構(gòu)建成功(圖3)。
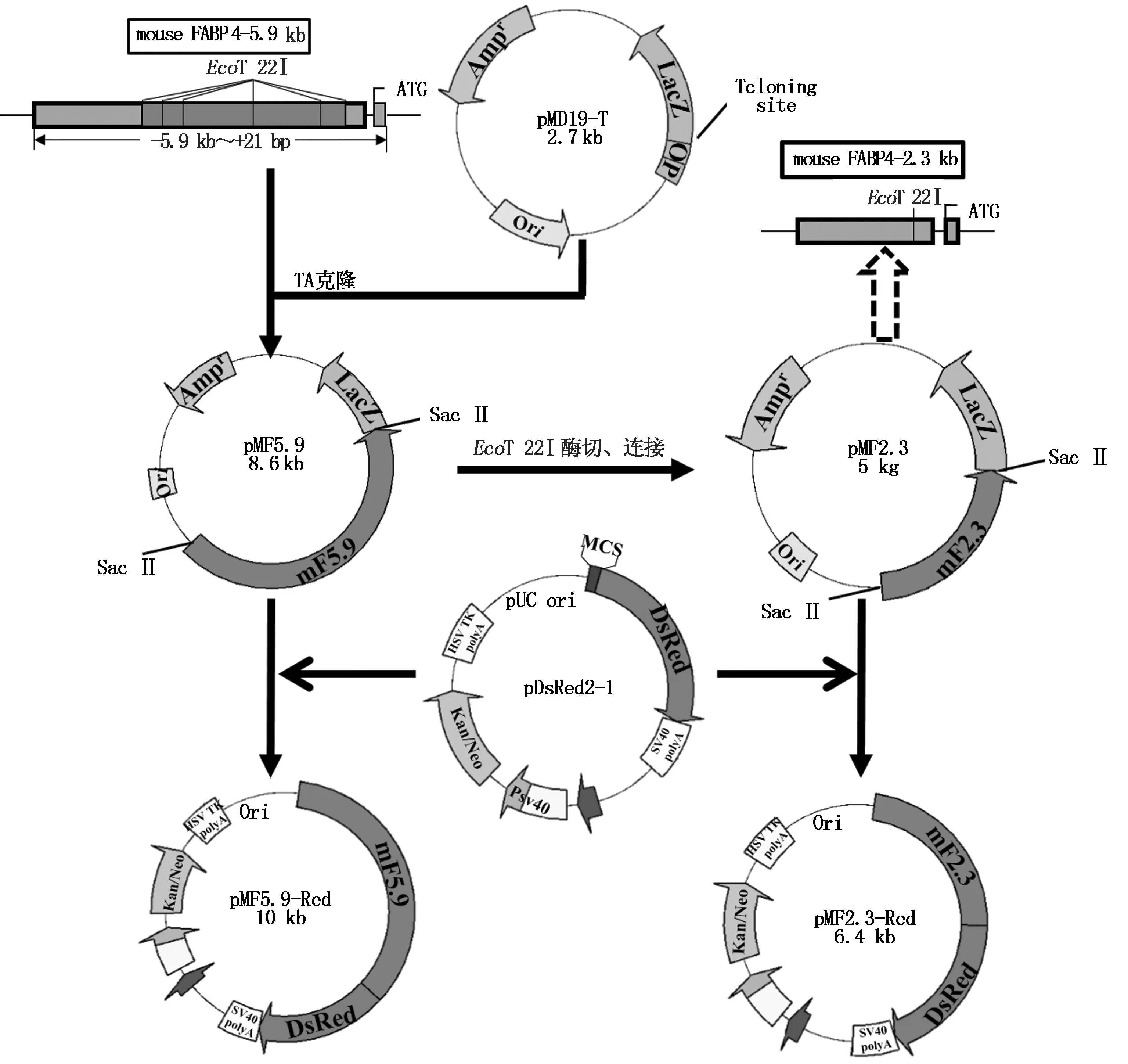
圖1 載體構(gòu)建示意圖Fig.1 Diagram of vector construction

圖2 小鼠FABP4啟動(dòng)子PCR擴(kuò)增Fig.2 Electrophoresis of mouse FABP4 promoter amplification

圖3 pMF5.9與pMF2.3質(zhì)粒的酶切鑒定結(jié)果Fig.3 Enzyme digestion result of plasmid pMF5.9 and pMF2.3
分別將質(zhì)粒pMF5.9和pMF2.3經(jīng)SacⅡ酶切后的小鼠FABP4啟動(dòng)子片段連入pDsRed2-1載體。將連接成功的質(zhì)粒pMF5.9-Red和pMF2.3-Red分別進(jìn)行EcoT 22I、EcoR Ⅴ與Hind Ⅲ、EcoR Ⅴ的酶切鑒定,結(jié)果符合預(yù)期,證明pMF5.9-Red和pMF2.3-Red質(zhì)粒構(gòu)建成功 (圖4) 。
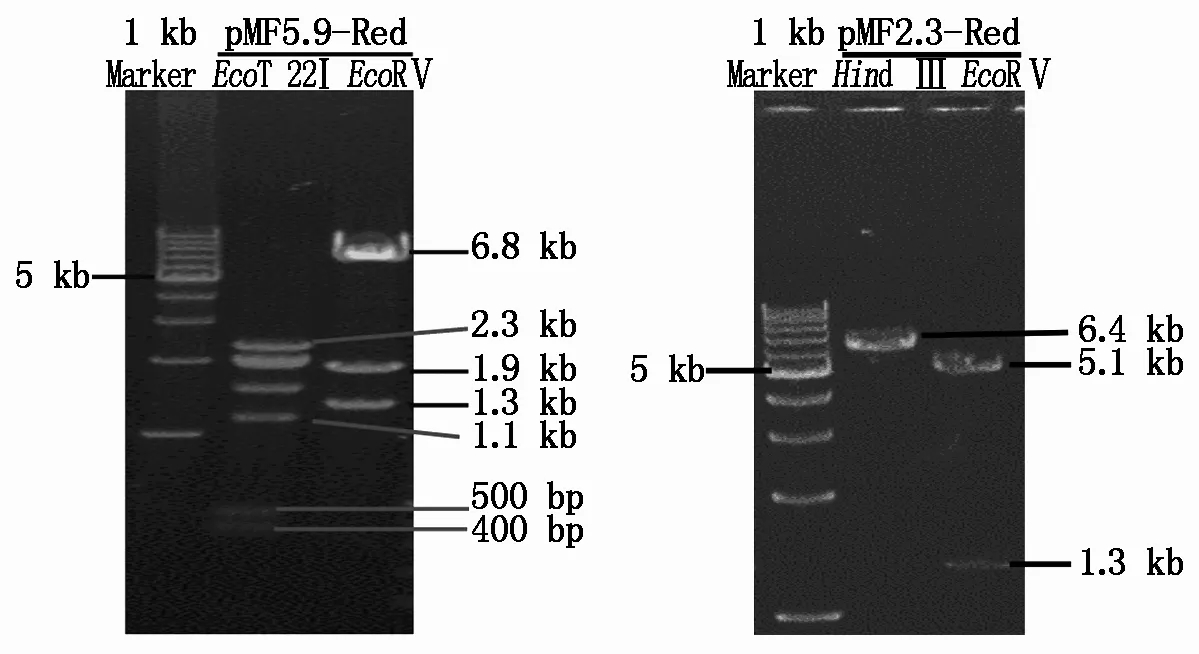
圖4 pMF5.9-Red 與pMF2.3-Red質(zhì)粒的酶切鑒定結(jié)果Fig.4 Enzyme digestion result of plasmid pMF5.9-Red and pMF2.3-Red
2.3 紅色熒光相對(duì)表達(dá)檢測(cè)
細(xì)胞分別轉(zhuǎn)染pMF5.9-Red和pMF2.3-Red載體后,實(shí)時(shí)定量PCR檢測(cè)結(jié)果顯示,轉(zhuǎn)染pMF2.3-Red質(zhì)粒后紅色熒光蛋白mRNA水平在牛脂肪間充質(zhì)干細(xì)胞中是pMF5.9-Red的2.28倍,而在牛胎兒成纖維細(xì)胞中是2.12倍(P<0.05)(圖5)。

*.P<0.05。
3 結(jié)論與討論
轉(zhuǎn)基因動(dòng)物的制作過(guò)程中,選擇合適的啟動(dòng)子元件是載體構(gòu)建的重要環(huán)節(jié),組成型啟動(dòng)子如CMV或PGK等,雖然可以獲得大量目的蛋白,但同時(shí)會(huì)對(duì)動(dòng)物的正常生命活動(dòng)造成一系列影響,出于提高試驗(yàn)結(jié)果準(zhǔn)確性以及動(dòng)物福利的考慮,應(yīng)用組織特異性啟動(dòng)子成為趨勢(shì)。
脂肪組織作為重要的能量存儲(chǔ)以及內(nèi)分泌器官,是研究能量代謝、激素信號(hào)通路以及肥胖、糖尿病等疾病的重要位點(diǎn)。最早開發(fā)的脂肪組織特異性啟動(dòng)子是成熟脂肪細(xì)胞的標(biāo)記基因—FABP4基因,它是脂肪酸結(jié)合蛋白基因家族的成員,主要在脂肪細(xì)胞中表達(dá)[9]。FABP4基因啟動(dòng)子/增強(qiáng)子元件在物種間保守性很高[10],小鼠的核心啟動(dòng)子區(qū)位于起始密碼子ATG上游-168~+21 bp的區(qū)域,增強(qiáng)子位于ATG上游-4.9~-5.4 kb的位置[11],人的啟動(dòng)子/增強(qiáng)子的位置和小鼠是一致的[12],而豬最高效的啟動(dòng)子在-2.8 kb~+51 bp[13]。應(yīng)用5.4 kb的FABP4基因全長(zhǎng)啟動(dòng)子/增強(qiáng)子制作的轉(zhuǎn)基因小鼠,可實(shí)現(xiàn)外源基因僅在生殖腺周圍的脂肪與皮下脂肪表達(dá)的目的[14]。利用全長(zhǎng)啟動(dòng)子/增強(qiáng)子驅(qū)動(dòng)Cre酶的轉(zhuǎn)基因小鼠,與帶有l(wèi)oxP位點(diǎn)的轉(zhuǎn)基因小鼠交配,則可以獲得在脂肪組織中敲除特定基因的小鼠子代[15]。
脂聯(lián)素(Adiponectin)基因啟動(dòng)子是最近鑒定的一種脂肪組織特異性啟動(dòng)子,該基因最有效的啟動(dòng)子區(qū)域同樣位于ATG上游5.4 kb的區(qū)域[16]。而PEPCK基因ATG上游-1 242~-828 bp則存在脂肪細(xì)胞特異的增強(qiáng)子元件[17]。以上兩者同樣也是制作脂肪細(xì)胞特異轉(zhuǎn)基因動(dòng)物常用的組織特異性元件。
組織特異啟動(dòng)子必須包含轉(zhuǎn)錄啟始的核心元件以及特異轉(zhuǎn)錄因子的結(jié)合位點(diǎn)。經(jīng)TRANSFAC軟件分析[10],本試驗(yàn)中克隆的FABP4基因啟動(dòng)子/增強(qiáng)子元件經(jīng)EcoT 22I酶切后的2.3 kb的啟動(dòng)子區(qū),保留了轉(zhuǎn)錄核心元件TATA box、CAAT box與增強(qiáng)子元件以及AP-1、TBF-2a、GATA-1等上游啟動(dòng)子元件,并且保留了PPRE、FSE、CEBPα、CEBPδ等脂肪特異轉(zhuǎn)錄因子結(jié)合位點(diǎn),本試驗(yàn)證明以上元件可保證外源基因在牛體細(xì)胞中的表達(dá)。
FABP4基因啟動(dòng)子/增強(qiáng)子元件已在小鼠[18]、大鼠[19]、山羊[20]、人[21]等的細(xì)胞中得到功能驗(yàn)證,為檢測(cè)上述元件在牛的體細(xì)胞中的啟動(dòng)效果,本研究選擇了牛胎兒成纖維細(xì)胞和脂肪間充質(zhì)干細(xì)胞進(jìn)行比較鑒定,其中胎兒成纖維細(xì)胞是常用的核移植供體細(xì)胞,脂肪間充質(zhì)干細(xì)胞可定向誘導(dǎo)分化為脂肪細(xì)胞。結(jié)果顯示5.9,2.3 kb的FABP4啟動(dòng)子/增強(qiáng)子均能夠驅(qū)動(dòng)目的基因在牛的上述細(xì)胞中表達(dá),表明該元件可以用于牛的轉(zhuǎn)基因研究。而且在這2種細(xì)胞中2.3 kb片段的轉(zhuǎn)錄水平均為5.9 kb的2倍以上,說(shuō)明2.3 kb片段的啟動(dòng)效率高于5.9 kb片段。但是該啟動(dòng)子在牛其他組織細(xì)胞中的啟動(dòng)特異性還有待進(jìn)一步的試驗(yàn)進(jìn)行驗(yàn)證。
綜上所述,本試驗(yàn)成功克隆了5.9 kb的FABP4基因全長(zhǎng)啟動(dòng)子區(qū),并通過(guò)酶切去除非核心啟動(dòng)子片段后,保留了2.3 kb的啟動(dòng)子區(qū),通過(guò)脂質(zhì)體轉(zhuǎn)染牛脂肪間充質(zhì)干細(xì)胞和牛胎兒成纖維細(xì)胞后,結(jié)果轉(zhuǎn)染pMF2.3-Red質(zhì)粒的細(xì)胞紅色熒光蛋白mRNA轉(zhuǎn)錄水平是pMF5.9-Red的2倍以上,證明小鼠FABP4啟動(dòng)子區(qū)可以驅(qū)動(dòng)外源基因在牛的體細(xì)胞中表達(dá)的同時(shí),經(jīng)過(guò)精簡(jiǎn)的2.3 kb啟動(dòng)子可將全長(zhǎng)啟動(dòng)子的啟動(dòng)效率提高1倍。為下一步制作脂肪特異表達(dá)外源基因的轉(zhuǎn)基因牛奠定了基礎(chǔ)。
[1] Bertolini L R,Meade H,Lazzarotto C R,et al.The transgenic animal platform for biopharmaceutical production[J].Transgenic Res,2016,25(3):329-343.
[2] Barlow C,Schroeder M,Lekstrom-Himes J,et al.Targeted expression of Cre recombinase to adipose tissue of transgenic mice directs adipose-specific excision of loxP-flanked gene segments[J].Nucleic Acids Res,1997,25(12):2543-2545.
[3] Abel E D,Peroni O,Kim J K,et al.Adipose-selective targeting of theGLUT4 gene impairs insulin action in muscle and liver[J].Nature,2001,409(6821):729-733.
[4] He W,Barak Y,Hevener A,et al.Adipose-specific peroxisome proliferator-activated receptor gamma knockout causes insulin resistance in fat and liver but not in muscle[J].Proc Natl Acad Sci U S A,2003,100(26):15712-15717.
[5] Imai T,Jiang M,Chambon P,et al.Impaired adipogenesis and lipolysis in the mouse upon selective ablation of the retinoid X receptor alpha mediated by a tamoxifen-inducible chimeric Cre recombinase (Cre-ERT2) in adipocytes[J].Proc Natl AcadSci U S A,2001,98(1):224-228.
[6] Zhang X,Xu A,Chung S K,et al.Selective inactivation of c-Jun NH2-terminal kinase in adipose tissue protects against diet-induced obesity and improves insulin sensitivity in both liver and skeletal muscle in mice[J].Diabetes,2011,60(2):486-495.
[7] Wong K E,Kong J,Zhang W,et al.Targeted expression of human vitamin D receptor in adipocytes decreases energy expenditure and induces obesity in mice[J].J Biol Chem,2011,286(39):33804-33810.
[8] Kanda H,Tateya S,Tamori Y,et al.MCP-1 contributes to macrophage infiltration into adipose tissue,insulin resistance,and hepatic steatosis in obesity[J].J Clin Invest,2006,116(6):1494-1505.
[9] Storch J,Thumser A E.Tissue-specific functions in the fatty acid-binding protein family[J].J Biol Chem,2010,285(43):32679-32683.
[10] Shin J,Li B,Davis M E,et al.Comparative analysis of fatty acid-binding protein 4 promoters:conservation of peroxisome proliferator-activated receptor binding sites[J].J Anim Sci,2009,87(12):3923-3934.
[11] Ross S R,Graves R A,Greenstein A,et al.A fat-specific enhancer is the primary determinant of gene expression for adipocyte P2invivo[J].Proc Natl Acad Sci U S A,1990,87(24):9590-9594.
[12] Rival Y,Stennevin A,Puech L,et al.Human adipocyte fatty acid-binding protein (aP2) gene promoter-driven reporter assay discriminates nonlipogenic peroxisome proliferator-activated receptor gamma ligands[J].J Pharmacol Exp Ther,2004,311(2):467-475.
[13] Jung E M,An B S,Kim Y K,et al.Generation of porcine fibroblasts overexpressing 11beta-HSD1 with adipose tissue-specific aP2 promoter as a porcine model of metabolic syndrome[J].Mol Med Rep,2013,8(3):751-756.
[14] Jurczak M J,Danos A M,Rehrmann V R,et al.Transgenic overexpression of protein targeting to glycogen markedly increases adipocytic glycogen storage in mice[J].Am J Physiol Endocrinol Metab,2007,292(3):952-963.
[15] Shi S Y,Luk C T,Brunt J J,et al.Adipocyte-specific deficiency of Janus kinase (JAK) 2 in mice impairs lipolysis and increases body weight,and leads to insulin resistance with ageing[J].Diabetologia,2014,57(5):1016-1026.
[16] Wang Z V,Deng Y,Wang Q A,et al.Identification and characterization of a promoter cassette conferring adipocyte-specific gene expression[J].Endocrinology,2010,151(6):2933-2939.
[17] Devine J H,Eubank D W,Clouthier D E,et al.Adipose expression of the phosphoenolpyruvate carboxykinase promoter requires peroxisome proliferator-activated receptor gamma and 9-cis-retinoic acid receptor binding to an adipocyte-specific enhancer in vivo[J].J Biol Chem,1999,274(19):13604-13612.
[18] Kershaw E E,Morton N M,Dhillon H,et al.Adipocyte-specific glucocorticoid inactivation protects against diet-induced obesity[J].Diabetes,2005,54(4):1023-1031.
[19] Pravenec M,Kazdova L,Landa V,et al.Transgenic and recombinant resistin impair skeletal muscle glucose metabolism in the spontaneously hypertensive rat[J].J Biol Chem,2003,278(46):45209-45215.
[20] 陳建文,劉 星,桂 濤,等.fat-1基因脂肪組織特異性表達(dá)載體的構(gòu)建及其山羊轉(zhuǎn)基因細(xì)胞系的建立[J].安徽農(nóng)業(yè)大學(xué)學(xué)報(bào),2012,39(5):746-750.
[21] Vargas D,Shimokawa N,Kaneko R,et al.Regulation of human subcutaneous adipocyte differentiation by EID1[J].J Mol Endocrinol,2016,56(2):113-122.
Construction of Red Fluorescent Protein Driven by Mouse FABP 4 Promoter and Expression in Bovine Cells
YUE Yongli,YU Haiquan
(Key Laboratory of Ministry of Education of China for Mammal Reproductive Biology and Biotechnology,Inner Mongolia University,Huhhot 010021,China)
The promoter/enhancer cassette of mouse adipocyte-type fatty acid binding protein(FABP4)was widely used as the adipocyte specific element.To detect the effect ofmFABP4 promoter on gene expression in bovine cells,5.9 kbmFABP4 promoter fragment was cloned from mouse liver genome and identified by digestion and sequencing after ligated to pMD19-T vector.By digestion of enzymeEcoT 22I the non-core promoter fragment was deleted from the 5.9 kb fragment and another 2.3 kb core promoter fragment was obtained.The 5.9 kb and 2.3 kb fragment were inserted into the multiple cloning site of plasmid pDs-Red 2-1 by enzymeSacⅡ,respectively.As a result the vectors,pMF5.9-Red and pMF2.3-Red,were constructed successfully.By lipofection,the constructed pMF5.9-Red and pMF2.3-Red vectors were transfected into bovine adipose-derived stem cells and bovine embryo fibroblast cells,respectively.24 h after transfection,Real-time PCR was applied to detect the transcription of red fluorescent protein in transfected cells.Results showed that the sequence of cloned 5.9 kb fragment was correct,and the 5.9 kb and the 2.3 kb fragments were correctly ligated into vectors.The mRNA of red fluorescent protein was both detected in transfected bovine adipose-derived stem cells and bovine embryo fibroblast cells,and the expression level displayed 2 fold higher in pMF2.3-Red than that of pMF5.9-Red.In conclusion,this study constructed the expression vector pMF5.9-Red and pMF2.3-Red with 5.9 kb or 2.3 kb fragment of mouseFABP4 gene as promoter region could initiate the transcription of foreign genes in bovine cells and the transcription efficiency by 2.3 kb promoter fragment was higher than that of 5.9 kb promoter fragment.
MouseFABP4 promoter;Vector construction;Cattle cells
2016-07-20
轉(zhuǎn)基因生物新品種培育科技重大專項(xiàng)(2014ZX08010005)
岳永莉(1982-),女,內(nèi)蒙古呼和浩特人,助理研究員,碩士,主要從事生殖生物學(xué)研究。
于海泉(1969-),男,內(nèi)蒙古烏海人,研究員,博士,主要從事生殖生物學(xué)研究。
Q782
A
1000-7091(2016)06-0026-05
10.7668/hbnxb.2016.06.005
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級(jí))(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學(xué)大眾(2021年6期)2021-07-20 07:42:44
科學(xué)(2020年3期)2020-11-26 08:18:30
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂(lè)園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學(xué)生學(xué)習(xí)指導(dǎo)(低年級(jí))(2017年10期)2017-10-10 01:00:05
湖北師范大學(xué)學(xué)報(bào)(自然科學(xué)版)(2015年2期)2016-01-10 08:41:55
云南中醫(yī)學(xué)院學(xué)報(bào)(2014年3期)2014-07-31 18:57:34
