溫度、鹽度及交互作用對(duì)僧帽牡蠣(Saccostreacucullata)呼吸排泄的影響
2017-02-08 05:58:27焦海峰嚴(yán)巧娜尤仲杰嚴(yán)小軍
生態(tài)學(xué)報(bào) 2017年2期
焦海峰,鄭 丹,嚴(yán)巧娜,尤仲杰,*,嚴(yán)小軍
1 寧波市海洋與漁業(yè)研究院,寧波 315012 2 寧波大學(xué)海洋學(xué)院,寧波 315211 3 寧波市江北區(qū)海洋與漁業(yè)局,寧波 315020
溫度、鹽度及交互作用對(duì)僧帽牡蠣(Saccostreacucullata)呼吸排泄的影響
焦海峰1,2,鄭 丹1,2,嚴(yán)巧娜3,尤仲杰1,2,*,嚴(yán)小軍2
1 寧波市海洋與漁業(yè)研究院,寧波 315012 2 寧波大學(xué)海洋學(xué)院,寧波 315211 3 寧波市江北區(qū)海洋與漁業(yè)局,寧波 315020
采用實(shí)驗(yàn)生態(tài)學(xué)方法研究了溫度(13、18、23、28、31、34℃)和鹽度(18、23、28、33)對(duì)僧帽牡蠣(Saccostreacucullata)耗氧率和排氨率的影響,并分析了二者交互作用對(duì)呼吸排泄的影響。結(jié)果表明,不同溫度條件下,僧帽牡蠣的耗氧率變化范圍為0.43—3.15mg g-1h-1,排氨率的變化范圍為72.62—1245.77μg g-1h-1,方差分析顯示,溫度和鹽度對(duì)僧帽牡蠣耗氧率(F=60.440,P<0.01)和排氨率(F=61.083,P<0.01)的影響均極顯著;二者的交互作用對(duì)僧帽牡蠣耗氧率(F=4.907,P<0.01)和排氨率(F=11.412,P<0.01)影響也極顯著,耗氧率(Y)、排氨率(Y)與溫度(T)和鹽度(S)之間的二元線性回歸方程為Y=a+bT+cS+dT×S。在一定溫度范圍內(nèi)(13—31℃),僧帽牡蠣單位體重耗氧率(Y)與溫度(T)呈正相關(guān),符合指數(shù)方程Y=aebT,其中a的取值范圍為0.1378—0.2117,b的取值范圍為0.0757—0.0912;單位體重排氨率也與溫度呈正相關(guān),符合上述指數(shù)方程,其中a的取值范圍為20.364—30.634,b的取值范圍為0.1004—0.1341。不同溫度和鹽度條件下,僧帽牡蠣呼吸排泄的氧氮比在2.20—7.78之間,平均值為3.81。
僧帽牡蠣;溫度;鹽度;耗氧率;排氮率
僧帽牡蠣(Saccostreacucullata)隸屬于軟體動(dòng)物門(Mollusca)、雙殼綱(Bivalvia)、牡蠣科(Ostreidae)、牡蠣屬(Ostrea),自然環(huán)境中主要棲息在中、低潮區(qū)的巖石上,是我國(guó)東南沿海重要的經(jīng)濟(jì)牡蠣[1-3],同時(shí),也是沿海典型的污損生物[4],是浙江沿海潮間帶生物的重要組成種類[3]。目前對(duì)僧帽牡蠣繁殖習(xí)性、生長(zhǎng)[5]、胚胎發(fā)育[2]、人工采苗及養(yǎng)殖[1]等基礎(chǔ)生物學(xué)方面的研究已有報(bào)道,但對(duì)其基礎(chǔ)生理代謝的研究資料缺乏,僅有焦海峰等[6]開展過僧帽牡蠣周年的耗氧率與排泄率的研究,林元燒等[7]開展過其濾水率的研究。
雙殼貝類是海洋生態(tài)系統(tǒng)的重要組成部分。資料表明,呼吸和排泄是貝類新陳代謝的基本生理活動(dòng),研究環(huán)境因子(如溫度、鹽度、pH等)和內(nèi)在因子(如規(guī)格、活動(dòng)狀態(tài)等)對(duì)呼吸和排泄的影響不但能夠了解動(dòng)物的生理狀態(tài)還能反映出對(duì)環(huán)境的適應(yīng)狀況[8]。在自然海區(qū)中,雙殼貝類的耗氧率、排氨率等生理生態(tài)學(xué)參數(shù)通常隨著季節(jié)性變化而變化[9],這些參數(shù)既能反應(yīng)雙殼貝類的生理特征,也能反映雙殼貝類與環(huán)境變化之間的相互影響[10]。目前,國(guó)內(nèi)外對(duì)雙殼類呼吸和排泄的已經(jīng)有較多研究[11-16]。綜合分析表明,溫度顯著地影響著雙殼貝類的新陳代謝活動(dòng),而鹽度對(duì)雙殼貝類呼吸、排泄影響的研究較少[15,17-18],但是溫度和鹽度又是決定水生生物自然分布的重要因子,它們是在貝類生理生態(tài)學(xué)研究中必須考慮的因素[19]。本文利用實(shí)驗(yàn)生態(tài)學(xué)的方法探討了溫度和鹽度對(duì)僧帽牡蠣呼吸與排泄的影響,以期了解僧帽牡蠣新陳代謝活動(dòng)的規(guī)律和變化特點(diǎn),為進(jìn)一步的能量學(xué)提供科學(xué)依據(jù),有助于評(píng)價(jià)雙殼貝類在海洋生物沉積中扮演的角色。
1 材料與方法
1.1 實(shí)驗(yàn)材料
僧帽牡蠣3—4月份取自象山港自然海區(qū)的巖礁相潮間帶,采集時(shí)保留外殼完整的個(gè)體,帶回實(shí)驗(yàn)室。挑選大小較勻稱的單個(gè)個(gè)體,用軟毛刷清除牡蠣表面的附著生物,在大型水族箱中暫養(yǎng),每天上午和下午分別投喂單細(xì)胞藻類,期間充氣,每隔1d換水1次,暫養(yǎng)時(shí)間不少于7d,實(shí)驗(yàn)前24h停止投喂。
1.2 方法
實(shí)驗(yàn)海水用砂濾后的自然海水,虹吸入水族箱中(水族箱容積約500L),用電加熱棒加熱調(diào)節(jié)溫度,每天升溫1—2℃,用市售海水晶和曝氣后的自來水調(diào)節(jié)鹽度,每天升降1—2。當(dāng)海水調(diào)制至實(shí)驗(yàn)條件后,穩(wěn)定控溫和控鹽12h以上,然后進(jìn)行實(shí)驗(yàn)。設(shè)置6個(gè)溫度梯度:13、18、23、28、31、34℃,設(shè)置4個(gè)鹽度梯度:18、23、28、33;每個(gè)溫度條件下均進(jìn)行4個(gè)鹽度梯度的實(shí)驗(yàn),共開展24組實(shí)驗(yàn)。實(shí)驗(yàn)?zāi)迪犜诟鳒囟群望}度條件下穩(wěn)定2d以上開始實(shí)驗(yàn)。
實(shí)驗(yàn)時(shí),將僧帽牡蠣沿瓶壁放入錐形瓶中,錐形瓶體積約5L,每瓶10只。利用虹吸法將錐形瓶中裝滿過濾海水后,海水適當(dāng)溢出1/4以清除氣泡對(duì)溶解氧的影響,然后用保鮮膜封口。每組實(shí)驗(yàn)設(shè)置3個(gè)重復(fù),并設(shè)對(duì)照組(不放牡蠣)3個(gè),水浴控溫。以牡蠣貝殼張開為起始點(diǎn)進(jìn)行計(jì)時(shí),實(shí)驗(yàn)2h后測(cè)定實(shí)驗(yàn)組水樣的溶解氧(DO)、氨氮(NH4-N)濃度和牡蠣的體積。
1.3 數(shù)據(jù)處理與計(jì)算方法
采用Winkler碘量法測(cè)定溶解氧,采用次溴酸鈉氧化法測(cè)定氨氮(水樣需要經(jīng)過0.45μm的GF/C膜過濾),在實(shí)驗(yàn)結(jié)束后用游標(biāo)卡尺測(cè)定僧帽牡蠣的殼長(zhǎng)(SL),然后解剖出的軟體部,在60℃下將烘干至恒重,用電子天平分別稱量軟體濕重(WW)和軟體干重(WD)。
實(shí)驗(yàn)開始前,測(cè)定3組對(duì)照瓶中溶解氧和氨氮濃度。實(shí)驗(yàn)結(jié)束后,根據(jù)測(cè)定瓶與對(duì)照瓶?jī)?nèi)水體的溶解氧和氨氮的濃度,計(jì)算僧帽牡蠣的耗氧率和排氨率。
OR=[(DO0-DOt)×(V-Vi)]/(WD×t)
式中,OR為單位軟體干重的耗氧率(mg g-1h-1),DO0和DOt為實(shí)驗(yàn)結(jié)束后對(duì)照組和測(cè)定組水中的DO含量(mg/L),DO0為對(duì)照組DO的平均值,V為呼吸瓶的水體積(L),Vi為僧帽牡蠣體積,WD為僧帽牡蠣軟體干重(g),t為實(shí)驗(yàn)時(shí)間(h)。
NR=[(Nt-N0)×(V-Vi)]/(WD×t)
式中,NR為單位軟體干重的排氨率(μg g-1h-1),N0和Nt為實(shí)驗(yàn)結(jié)束后對(duì)照組和測(cè)定組中水中的NH4-N的濃度 (μg/L),N0對(duì)照組NH4-N的平均值。
實(shí)驗(yàn)結(jié)果用平均值±標(biāo)準(zhǔn)誤(Mean±SE)表示,數(shù)據(jù)先用Excell預(yù)處理,然后用SPSS19.0軟件進(jìn)行統(tǒng)計(jì)分析處理(單因素方差分析ANOVA和回歸分析)。
實(shí)驗(yàn)用僧帽牡蠣的生物學(xué)參數(shù)(Mean±SD):殼長(zhǎng)(37.61±6.21)mm,軟體濕重(8.86±2.63)g,軟體干重(1.96±0.86)g。
2 結(jié)果
2.1 溫度和鹽度對(duì)僧帽牡蠣耗氧率的影響
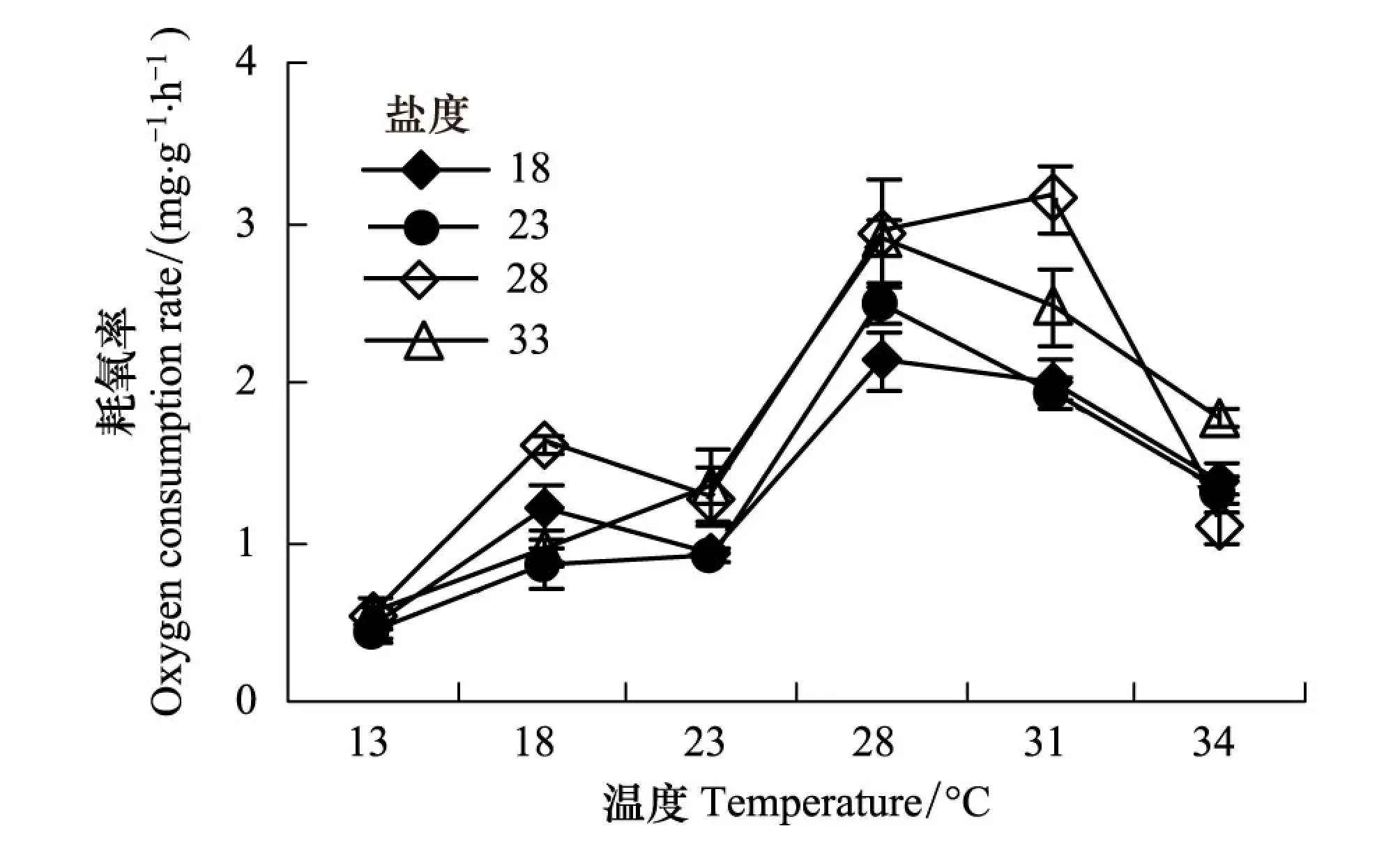
圖1 溫度對(duì)不同鹽度條件下泥蚶耗氧率的影響 Fig.1 The effect of temperature on the oxygen consumption rate of Saccostrea cucullata in different salinity
實(shí)驗(yàn)結(jié)果表明:在4個(gè)鹽度梯度下,僧帽牡蠣耗氧率隨溫度的變化趨勢(shì)較為一致,均在13℃時(shí)最低,隨著溫度的升高而升高,到28℃或31℃時(shí)出現(xiàn)峰值,溫度為34℃時(shí),耗氧率大幅下降。由圖1看出,僧帽牡蠣耗氧率的變化范圍為0.43—3.15 mg g-1h-1,最低值出現(xiàn)在13℃,最高值出現(xiàn)在31℃。不同溫度時(shí),僧帽牡蠣的耗氧率的變化均值為0.51—2.61 mg g-1h-1,峰值出現(xiàn)在28℃時(shí)。單因素方差分析顯示,僧帽牡蠣耗氧率受溫度的影響極顯著(F=60.440,P<0.01)。利用線性回歸分析發(fā)現(xiàn),在溫度13—31℃范圍內(nèi),僧帽牡蠣耗氧率與溫度呈顯著的指數(shù)相關(guān)lnY=lna+blnT(Y=aebT)(表1),其中a的取值范圍為0.1378—0.2117,b的取值范圍為0.0757—0.0912。
鹽度梯度的變化對(duì)僧帽牡蠣耗氧率也有影響,基本規(guī)律是隨鹽度升高耗氧率升高,在鹽度升高到一定范圍后,耗氧率又下降。不同鹽度梯度下,牡蠣耗氧率的值分別為1.36 mg g-1h-1、1.31 mg g-1h-1、1.77 mg g-1h-1和1.67 mg g-1h-1,單因素方差分析顯示,各鹽度梯度下耗氧率差異極不顯著(F=1.391,P=0.253)。
2.2 溫度和鹽度對(duì)僧帽牡蠣排氨率的影響
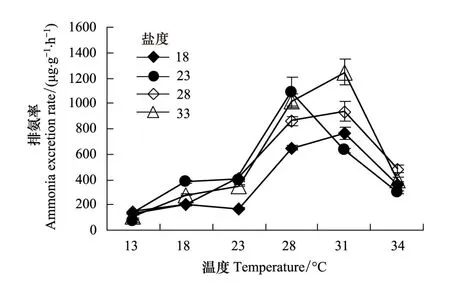
圖2 溫度對(duì)不同鹽度條件下僧帽牡蠣排氨率的影響 Fig.2 The effect of temperature on the ammonia excretion rate of Saccostrea cucullata in different salinity
由圖2可見,僧帽牡蠣排氨率與耗氧率的變化趨勢(shì)基本一致,溫度低時(shí)排氨率低,溫度在13—31℃的范圍內(nèi)升高時(shí),排氨率升高,溫度為34℃時(shí),排氨率下降。僧帽牡蠣排氨率的變化范圍72.62—1245.17μg g-1h-1,峰值出現(xiàn)在13℃和31℃時(shí)。不同溫度時(shí),僧帽牡蠣排氨率的變化范圍為114.08—904.99μg g-1h-1,峰值出現(xiàn)在28℃時(shí)。單因素方差分析結(jié)果顯示,溫度對(duì)僧帽牡蠣排氨率的影響極顯著(F=61.083,P<0.01)。利用線性回歸分析發(fā)現(xiàn),在適溫范圍內(nèi),僧帽牡蠣排氨率也與溫度呈顯著的指數(shù)相關(guān)(表1),其中a的取值范圍為20.364—30.634,b的取值范圍為0.1004—0.1341。
鹽度對(duì)僧帽牡蠣排氨率的影響不明顯。不同鹽度梯度下,僧帽牡蠣耗氧率的值分別為380.58 μg g-1h-1、477.54 μg g-1h-1、503.85 μg g-1h-1和556.48 μg g-1h-1,單因素方差分析顯示,各鹽度梯度下排氨率差異不顯著(F=0.928,P=0.432)。
2.3 溫度和鹽度的交互作用對(duì)對(duì)耗氧率與排氨率的影響
利用單變量線性模型(Univariate)分析表明:在一定溫度范圍(13—31℃)和鹽度范圍(18—28)內(nèi),溫度和鹽度的交互作用對(duì)僧帽牡蠣的耗氧率(F=4.907,P<0.01)和排氨率(F=11.412,P<0.01)影響均極顯著(表2)。
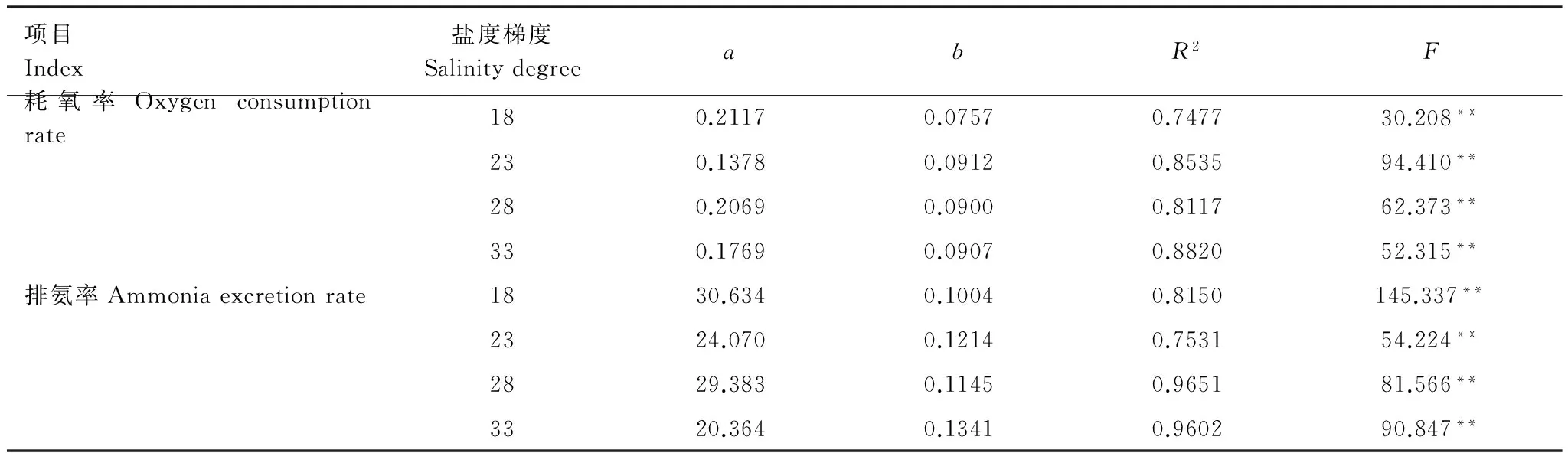
表1 僧帽牡蠣耗氧率、排氨率與溫度的回歸方程
* 代表差異顯著,**代表差異極顯著
表2 溫度、鹽度對(duì)僧帽牡蠣耗氧率與排氨率影響的交互作用檢驗(yàn)
Table 2 The test of interaction of temperature and salinity to oxygen consumption rate and ammonia excretion rate ofSaccostreacucullata

來源Soucres耗氧率Oxygenconsumptionrate排氨率Ammoniaexcretionrate平方和類型(III)TypeIIISumofSquares均方MeansquareFP平方和類型(III)TypeIIISumofSquares均方MeansquareFP模型校正CorrectedModel32.7932.34253.7150.0004575223.597326801.68566.4090.000溫度Watertemperature27.8246.956159.5190.0003964428.735991107.184201.4030.000鹽度Salinity3.2571.62837.3430.000161507.00780753.50316.4100.000溫度×鹽度Watertemperature×sa-linity1.7120.2144.9070.001449287.85656160.98211.4120.000誤差Error1.3080.044147630.3084921.010總和校正CorrectedTo-tal34.1014722853.905
在上述條件下,利用線性回歸(Linear Regression)分析,耗氧率(OR)、排氨率 (NR)與溫度(T)和鹽度(S)的二元線性回歸方程分別為:OR=0.631-0.070T-0.016S+0.006T×S(R2=0.883);NR=-133.376-15.305T+14.014S+1.227T×S(R2=0.872)。
2.4 溫度、鹽度與僧帽牡蠣呼吸排泄O∶N的關(guān)系
氧氮比是動(dòng)物呼吸排泄的重要生理指標(biāo),由表3可以看出,僧帽牡蠣呼吸排泄的O∶N比在2.20—7.78之間,平均值為3.81。其中O∶N比隨溫度的上升成凹陷的弧形變化,而隨鹽度的上升變化趨勢(shì)不甚明顯。方差分析表明,不同溫度范圍內(nèi)的O∶N比有極顯著差異(F=4.459,P<0.01),差異的來源于低溫實(shí)驗(yàn)條件下(13℃和18℃)的O∶N比。鹽度對(duì)僧帽牡蠣呼吸排泄O∶N比的影響不顯著(F=0.800,P=0.498)。
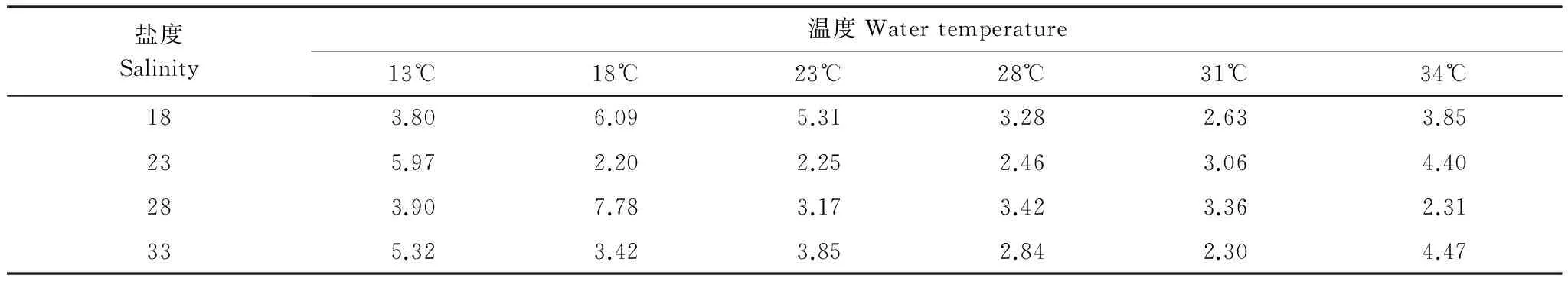
表3 不同溫度、鹽度梯度下僧帽牡蠣呼吸排泄的氧氮比值
3 討論
3.1 溫度對(duì)貝類呼吸排泄的影響
耗氧率和排氨率是動(dòng)物有氧代謝的重要指標(biāo),研究貝類的耗氧率和排氨率有助于了解它們新陳代謝活動(dòng)的規(guī)律和變化特點(diǎn),也能夠反映環(huán)境條件對(duì)其生理活動(dòng)的影響[20-21]。貝類是變溫動(dòng)物,溫度是影響貝類生理活動(dòng)的重要環(huán)境因素,研究者普遍認(rèn)為溫度對(duì)貝類耗氧率和排氨率有顯著影響[22-26]。董志國(guó)等[22]研究彩虹明櫻蛤發(fā)現(xiàn),在水溫低于28℃時(shí)耗氧率隨著水溫的上升呈線型上升,排氨率也與溫度呈正相關(guān)。李金碧等[14]對(duì)櫛江珧研究發(fā)現(xiàn),溫度對(duì)耗氧率和排氨率影響也呈線性關(guān)系,耗氧率和排氨率在18℃最低,34℃最高。張媛等[17]對(duì)橄欖蚶的研究也表明,耗氧率和排氨率隨著溫度的升高而上升。然而,溫度對(duì)耗氧率和排氨率的影響趨勢(shì)并不一致。楊紅生等[11]對(duì)墨西哥灣扇貝研究表明,在一定溫度范圍內(nèi),耗氧率隨溫度的升高而上升,當(dāng)水溫超過30℃后,耗氧率下降,排氨率是隨著溫度的升高持續(xù)增加。王俊等[12]對(duì)櫛孔扇貝研究發(fā)現(xiàn),在8—23℃間耗氧率和排氨率均隨水溫升高而增大,當(dāng)溫度超過28℃時(shí)耗氧率下降,排氨率增加。隨溫度上升耗氧率出現(xiàn)下行拐點(diǎn),排氨率卻上升的現(xiàn)象也出現(xiàn)在方斑東風(fēng)螺中[23]。然而,文海翔等[13]研究硬殼蛤發(fā)現(xiàn),耗氧率和排氨率隨溫度的升高而升高,峰值出現(xiàn)在20—30℃之間,在30℃以后隨著溫度上升,兩者均下降。孟學(xué)平等[24]研究西施舌也表明,溫度對(duì)西施舌耗氧率影響顯著,在10—20℃,耗氧率隨水溫的升高而升高,當(dāng)超過25℃以后,耗氧率則下降。排氨率在10—20℃間上升緩慢,在25℃后氨代謝急劇增加,之后下降。徐東等[25]對(duì)蝦夷扇貝的研究表明,耗氧率和排氨率均隨溫度的緩慢升高而增加,在15℃達(dá)到最大值,之后隨溫度的升高而減小。王雅倩等[26]對(duì)中國(guó)蛤蜊研究表明,耗氧率先隨著溫度的升高而上升,當(dāng)溫度超過23℃后,隨著溫度的升高而降低,排氨率隨溫度的變化趨勢(shì)與耗氧率的變化趨勢(shì)一致。
綜合分析表明,溫度對(duì)貝類耗氧率和排氨率的影響可以表現(xiàn)為3種形式:一是耗氧率和排氨率都與溫度呈線型關(guān)系,隨著溫度上升而上升;二是排氨率與溫度呈線型關(guān)系,而耗氧率對(duì)溫度變化有最大響應(yīng)區(qū),耗氧率先隨溫度上升而上升,待上升到峰值后,隨著溫度的上升而下降;三是耗氧率和排氨率均對(duì)溫度的變化有最大響應(yīng),先上升后下降。本文中,僧帽牡蠣的耗氧率和排氨率均隨著溫度的上升表現(xiàn)出最大響應(yīng)現(xiàn)象,在溫度13—28℃時(shí),耗氧率隨溫度上升而上升,但溫度超過28℃后,耗氧率和排氨率均下降明顯,這說明溫度超過28℃后,牡蠣處于不適狀態(tài),代謝出現(xiàn)了抑制甚至停止。一般認(rèn)為溫度升高,動(dòng)物組織器官活性提高,生物體內(nèi)的酶活性也隨著溫度升高而加強(qiáng),從而加速了物質(zhì)代謝水平,致使呼吸和代謝加快;當(dāng)超過極限溫度后,就會(huì)引起生理功能的紊亂,呼吸和代謝活動(dòng)下降,這也是變溫動(dòng)物的普遍特征[20]。因此,不同貝類呼吸代謝的規(guī)律雖然受溫度影響顯著,但不同種類之間的變化規(guī)律也不盡相同,這種差異性還需要深入研究。
3.2 鹽度對(duì)貝類呼吸排泄的影響
鹽度是海洋生態(tài)系統(tǒng)中重要的生態(tài)因子, 影響著貝類的分布和其生理代謝,鹽度變化對(duì)貝類耗氧率和排氨率也有明顯的影響。例如,趙文等[27]研究了鹽度對(duì)中國(guó)蛤蜊耗氧率和排氨率的影響,結(jié)果發(fā)現(xiàn),在鹽度升高的過程中,耗氧率和排氨率都是增加的,峰值出現(xiàn)在18時(shí),然后隨著鹽度的升高而下降。Stickle等[28]對(duì)紫貽貝(Mytilusedulis)的研究表明,在鹽度10—30范圍內(nèi),耗氧率隨鹽度的降低而增加。羅杰等[29]對(duì)管角螺研究發(fā)現(xiàn),耗氧率隨鹽度的升高而增加,在27時(shí)出現(xiàn)峰值,隨后耗氧率隨鹽度的升高而下降鹽度對(duì)排氨率的影響與對(duì)耗氧率的影響趨勢(shì)基本一致。郭海燕等[30]在大西洋浪蛤(Spisulasolidissima)的研究中發(fā)現(xiàn), 在鹽度 19—43 范圍內(nèi), 它的耗氧率和排氨率都隨鹽度升高而增大, 達(dá)到最大值后逐漸下降。黃洋等[31]研究尖紫蛤(Soletellinaacuta)表明,耗氧率隨著鹽度的升高而增加,在鹽度為10時(shí)達(dá)到最大值,當(dāng)鹽度超過10時(shí),耗氧率就下降。李曉燕[32]研究三倍體和二倍體太平洋牡蠣表明,隨著鹽度的降低,耗氧率均降低,在鹽度為20時(shí)達(dá)到最低,然后隨著鹽度的繼續(xù)降低,耗氧率稍有上升。隨著鹽度的降低,兩種牡蠣排氨率出現(xiàn)不同程度的升高,在鹽度20時(shí)達(dá)到最高,然后隨著鹽度的持續(xù)降低,排氨率大幅度降低。李金碧等[14]研究櫛江珧發(fā)現(xiàn),在18℃耗氧率和排氨率最低,在34℃時(shí)最高,耗氧率和排氨率隨鹽度升高先降低后升高,在鹽度為31時(shí)為最小值,在鹽度為41時(shí)為最大值。Farmer等[33]研究貝類Acartiatonsa表明,耗氧率和排氨率在適宜鹽度條件下最低,當(dāng)鹽度升高或降低時(shí)耗氧率和排氨率均升高。本研究發(fā)現(xiàn),鹽度對(duì)僧帽牡蠣耗氧率和排氨率影響的變化趨勢(shì)基本一致,均存在最大鹽度響應(yīng)區(qū)。但是同是牡蠣,也有不同的規(guī)律,如王剛等[15]對(duì)近江牡蠣的研究表明,在低鹽環(huán)境下(5—15),隨著鹽度的升高,耗氧率、排氨率均下降,在鹽度為15時(shí)降至最低,然后隨著鹽度的升高,耗氧率和排氨率升高很快,且幅度較大。
鹽度對(duì)貝類呼吸排泄的影響較為復(fù)雜,其規(guī)律性不顯著,除上述幾種典型的規(guī)律外,Barber等[34]對(duì)海灣扇貝的研究表明,在鹽度17—34范圍內(nèi),排氨率隨鹽度的升高而下降。有研究者認(rèn)為貝類在其適宜鹽度時(shí)耗氧率和排氨率較低,是因?yàn)樗鼈兲幵邴}度等滲點(diǎn),只需要消耗較少的能量便能維持自身的生理活動(dòng),當(dāng)鹽度不適時(shí),體內(nèi)滲透壓不平衡,為維持滲透壓需要消耗一定的能量,導(dǎo)致呼吸排泄活躍。也有學(xué)者認(rèn)為,低鹽環(huán)境導(dǎo)致貝類關(guān)閉貝殼或者水管,從而減低了有機(jī)體與外界的水流交換,以保護(hù)貝類不受低鹽造成的傷害。生活在潮間帶的生物要適應(yīng)周圍環(huán)境鹽度的變化就要調(diào)整其生理代謝活動(dòng)[12],目前普遍認(rèn)為,貝類屬于滲透壓調(diào)節(jié)者,它們體內(nèi)存在著一個(gè)游離氨基酸庫(kù),當(dāng)外界環(huán)境中鹽度變化時(shí),它們可以通過調(diào)節(jié)機(jī)體內(nèi)游離氨基酸或者其它小分子有機(jī)物濃度等來調(diào)整體內(nèi)滲透壓的變化,從而適應(yīng)環(huán)境條件[33,35]。
綜合分析發(fā)現(xiàn),溫度和鹽度對(duì)僧帽牡蠣耗氧率(F=4.907,P<0.01)和排氨率(F=11.412,P<0.01)的影響存在著極顯著交互作用。除溫度和鹽度外,影響貝類呼吸和排泄的因素還很多,如體重、餌料濃度、pH、溶解氧及個(gè)體大小、性腺發(fā)育程度等,王芳等[36]研究發(fā)現(xiàn),濕重和溫度的交互作用對(duì)海灣扇貝和太平洋牡蠣的耗氧率影響也較為顯著。因此,貝類的呼吸排泄是復(fù)雜的生命過程之一,環(huán)境因素的影響也不一而足,要探明環(huán)境因子對(duì)貝類呼吸代謝的的影響規(guī)律或機(jī)制,需要從多學(xué)科深入探討。
3.3 氧氮比
貝類是以體內(nèi)碳水化合物、 脂肪和蛋白質(zhì)為供能來源,氧氮比(O∶N)可以表征動(dòng)物呼吸底物的類別,反映動(dòng)物在特定狀態(tài)下體內(nèi)的脂肪、糖類和蛋白質(zhì)代謝的比例關(guān)系[37]。不同氧氮比代表不同供能底物的研究已經(jīng)非常廣泛[38-39]。另外,氧氮比與環(huán)境對(duì)有機(jī)體的壓力相關(guān),其值的高低可以作為生物體適應(yīng)環(huán)境壓力的指標(biāo)之一[21]。本文中僧帽牡蠣的呼吸排泄氧氮比較低,明顯低于墨西哥灣扇貝[11]、硬殼蛤[13]、橄欖蚶[16]以及其它貝類[19-30],也低于近江牡蠣[15],略低于僧帽牡蠣周年呼吸排泄氧氮比,其呼吸排泄周年氧氮比的變化范圍為3.04—10.27[6]。產(chǎn)生上述差異的原因,推測(cè)是因?yàn)樯蹦迪牉槌遍g帶棲息的種類,經(jīng)常處于干露狀態(tài),它們的生理活動(dòng)狀態(tài)長(zhǎng)期受到外界環(huán)境的脅迫,故呼吸排泄速率較低,氧氮比不高。本文還發(fā)現(xiàn):在溫度較低時(shí),僧帽牡蠣的氧氮比較高,隨著溫度的上升,氧氮比先下降后上升;而隨著鹽度的上升,氧氮比下降。這與僧帽牡蠣呼吸排泄氧氮比周年變化規(guī)律較為一致,均為低溫時(shí)氧氮比高、高溫時(shí)氧氮比低[6]。總體來看,僧帽牡蠣活動(dòng)較為緩慢,日常代謝中需要消耗蛋白質(zhì)來滿足能量需求[6],當(dāng)它們?cè)馐軄碜酝饨绲拿{迫時(shí),其氧氮比更低,這與貝類遭受外界脅迫從而降解體內(nèi)蛋白質(zhì)形成氨基酸的生理適應(yīng)相吻合[32],也與Marcela等[40]認(rèn)為氧氮比可以作為評(píng)估生理學(xué)調(diào)節(jié)工具的推斷相吻合。
[1] 陳和鵬. 僧帽牡蠣海區(qū)半人工采苗及棚架式養(yǎng)殖. 福建水產(chǎn), 1992, (1): 6-9.
[2] 柯才煥, 李少菁, 李復(fù)雪, 鄭重. 僧帽牡蠣幼體附著和變態(tài)的誘導(dǎo). 水產(chǎn)學(xué)報(bào), 2000, 24(3): 229-234.
[3] 尤仲杰, 徐善良, 謝起浪. 浙江沿岸的貝類資源及其增養(yǎng)殖. 東海海洋, 2000, 18(1): 50-56.
[4] 王寶強(qiáng), 薛俊增, 莊驊, 吳惠仙. 洋山港海域大型污損生物生態(tài)特點(diǎn). 海洋學(xué)報(bào), 2012, 34(3): 155-162.
[5] 張璽, 樓子康. 僧帽牡蠣的繁殖和生長(zhǎng)的研究. 海洋與湖沼, 1957, 1(1): 123-138.
[6] 焦海峰, 項(xiàng)翔, 尤仲杰, 嚴(yán)小軍. 泥蚶、縊蟶和僧帽牡蠣呼吸與排泄的周年變化. 海洋學(xué)報(bào), 2013, 35(6): 147-153.
[7] 林元燒, 曹文清, 羅文新, 郭東暉, 鄭愛榕, 黃長(zhǎng)江. 幾種主要養(yǎng)殖貝類濾水率的研究. 海洋學(xué)報(bào), 2003, 25(1): 86-92.
[8] 冉福, 曲憲成, 沈和定, 陳宇昕, 徐偉. 塔瑪亞歷山大藻對(duì)文蛤呼吸和排泄的影響. 海洋環(huán)境科學(xué), 2009, 28(1): 30-33.
[9] Newell R I E. Ecosystem influences of natural and cultivated populations of suspension-feeding bivalve molluscs: a review. Journal of Shellfish Research, 2004, 23(1): 51-61.
[10] Prins T C, Smaal A C, Dame R F. A review of the feedbacks between bivalve grazing and ecosystem processes. Aquatic Ecology, 1998, 31(4): 349-359.
[11] 楊紅生, 張濤, 王萍, 何義朝, 張福綏. 溫度對(duì)墨西哥灣扇貝耗氧率及排泄率的影響. 海洋學(xué)報(bào), 1998, 20(4): 91-96.
[12] 王俊, 姜祖輝, 唐啟升. 櫛孔扇貝耗氧率和排氨率的研究. 應(yīng)用生態(tài)學(xué)報(bào), 2002, 13(9): 1157-1160.
[13] 文海翔, 張濤, 楊紅生, 劉保忠, 周毅, 毛玉澤, 張福綏. 溫度對(duì)硬殼蛤Mercenariamercenaria(Linnaeus, 1758)呼吸排泄的影響. 海洋與湖沼, 2004, 35(6): 549-554.
[14] 李金碧, 龔世園, 喻達(dá)輝. 溫度和鹽度對(duì)櫛江珧耗氧率和排氨率的影響. 安徽農(nóng)業(yè)科學(xué), 2009, 37(5): 2016-2018.
[15] 王剛, 張其中, 喻子牛, 林小濤. 鹽度對(duì)近江牡蠣耗氧率、排氨率、O∶N和吸收率的影響. 海洋科學(xué), 2013, 37(2): 101-106.
[16] Hao Z L, Tang X J, Ding J, Ben Y, Chang Y Q. Effect of high temperature on survival, oxygen consumption, behavior, ammonia-N excretion, and related immune indicators of the Japanese scallopMizuhopectenyessoensis. Aquaculture International, 2014, 22(6): 1863-1876.
[17] 張媛, 方建光, 毛玉澤, 張繼紅, 薛素燕. 溫度和鹽度對(duì)橄欖蚶耗氧率和排氨率的影響. 中國(guó)水產(chǎn)科學(xué), 2007, 14(4): 690-694.
[18] 唐保軍, 尹飛, 歸從時(shí). 海水鹽度對(duì)紫血蛤耗氧率和排氨率的影響. 臺(tái)灣海峽, 2012, 31(1): 89-94.
[19] Yu Z H, Qi Z H, Hu C Q, Liu W G, Huang H H. Effects of salinity on ingestion, oxygen consumption and ammonium excretion rates of the sea cucumberHolothurialeucospilota. Aquaculture Research, 2013, 44(11): 1760-1767.
[20] Hammen C S. Aminotransferase activities and amino acid excretion of bivalve mollusks and brachiopods. Comparative Biochemistry & Physiology, 1968, 26(2): 697-705.
[21] Widdows J, Bayne B L. Temperature acclimation ofMytilusEduliswith reference to its energy budget. Journal of the Marine Biological Association of the United Kingdom, 1971, 51(4): 827-843.
[22] 董志國(guó), 李曉英, 孟學(xué)平, 程漢良, 閻斌倫, 陳漢春, 王美珍. 彩虹明櫻蛤的耗氧率與排氨率研究. 臺(tái)灣海峽, 2006, 25(4): 503-508.
[23] 劉建勇, 鄧興朝, 紹杰. 體重和溫度對(duì)方斑東風(fēng)螺耗氧率和排氨率的影響. 中國(guó)水產(chǎn)科學(xué), 2005, 12(3): 239-244.
[24] 孟學(xué)平, 董志國(guó), 程漢良, 李曉英, 李家樂. 西施舌的耗氧率與排氨率研究. 應(yīng)用生態(tài)學(xué)報(bào), 2005, 16(12): 2435-2438.
[25] 徐東, 張繼紅, 王文琪, 杜美榮, 任黎華, 王忠. 溫度變化對(duì)蝦夷扇貝耗氧率和排氨率的影響. 中國(guó)水產(chǎn)科學(xué), 2010, 17(5): 1101-1106.
[26] 王雅倩, 趙文, 程芳晉, 楊增強(qiáng). 體重和溫度對(duì)中國(guó)蛤蜊耗氧率和排氨率的影響. 大連水產(chǎn)學(xué)院學(xué)報(bào), 2009, 24(6): 544-548.
[27] 趙文, 王雅倩, 魏杰, 王麗. 體重和鹽度對(duì)中國(guó)蛤蜊耗氧率和排氨率的影響. 生態(tài)學(xué)報(bào), 2011, 31(7): 2040-2045.
[28] Stickle W B, Sabourin T D. Effects of salinity on the respiration and heart rate of the common mussel,MytilusedulisL., and the black chiton,Katherinatunicata(Wood). Journal of Experimental Marine Biology and Ecology, 1979, 41(3): 257-268.
[29] 羅杰, 劉楚吾, 李鋒, 曾理想, 陳加輝. 鹽度及規(guī)格對(duì)管角螺耗氧率和排氨率的影響. 海洋科學(xué), 2008, 32(5): 46-50.
[30] 郭海燕, 王昭萍, 于瑞海, 王芳, 林志華. 溫度、鹽度對(duì)大西洋浪蛤耗氧率和排氨率的影響. 中國(guó)海洋大學(xué)學(xué)報(bào), 2007, 31(增刊): 185-188.
[31] 黃洋, 黃海立, 林國(guó)游, 孫成波, 劉志剛. 鹽度、pH和規(guī)格對(duì)尖紫蛤(Soletellinaacuta)耗氧率和排氨率的影響. 海洋與湖沼, 2013, 44(1): 120-125.
[32] 李曉艷. 鹽度對(duì)三倍體和二倍體太平洋牡蠣代謝影響的比較. 大連大學(xué)學(xué)報(bào), 2014, 35(3): 88-91.
[33] Farmer L, Reeve M R. Role of the free amino acid pool of the copepodAcartiatonsain adjustment to salinity change. Marine Biology, 1978, 48(4): 311-316.
[34] Barber B J, Blake N J. Substrate catabolism related to reproduction in the bay scallopArgopectenirradiansconcentricus, as determined by O/N and RQ physiological indexes. Marine Biology, 1985, 87(1): 13-18.
[35] Zurburg W, De Zwaan A. The role of amino acids in anaerobiosis and osmoregulation in bivalves. Journal of Experimental Zoology, 1981, 215(3): 315-325.
[36] 王芳, 董雙林, 張碩, 潘克厚. 海灣扇貝(Argopectenirradians)和太平洋牡蠣(Crassostreagigas)呼吸和排泄的研究. 青島海洋大學(xué)學(xué)報(bào), 1998, 28(2): 233-239.
[37] Ruyet J P L, Mahé K, Le Bayon N, Le Delliou H. Effects of temperature on growth and metabolism in a Mediterranean population of European sea bass,Dicentrarchuslabrax. Aquaculture, 2004, 237(1/4): 269-280.
[38] Ikeda T. Nutritional ecology of marine zooplankton. Memoirs of the Faculty of Fisheries Hokkaido University, 1974, 22(1): 1-97.
[39] Mayzaud P. Respiration and nitrogen excretion of zooplankton. Ⅳ. the influence of starvation on the metabolism and biochemical composition of some species. Marine Biology, 1976, 37(1): 47-58.
[40] Montagna M C, Collins P A. Oxygen consumption and ammonia excretion of the freshwater crabTrichodactylusborellianusexposed to chlorpyrifos and endosulfan insecticides. Pesticide Biochemistry and Physiology, 2008, 92(3): 150-155.
Effects of water temperature and salinity on oxygen consumption rate and ammonia excretion rate ofSaccostreacucullata
JIAO Haifeng1,2, ZHENG Dan1,2, YAN Qiaona3, YOU Zhongjie1,2,*, YAN Xiaojun2
1NingboAcademyofOceanologyandFishery,Ningbo315012,China2OceancollegeofNingboUniversity,Ningbo315211,China3JiangbeiOceanandFisheryBureau,Ningbo315020,China
Intertidal oyster (Saccostreacucullata) is an important cultivated species and macro-fouling organism in the southeastern China Sea, and it is an important model organism for studies on shellfish physiology and marine ecology. Oxygen consumption rate and ammonia excretion rate are important parameters of shellfish physiology and metabolism, and can be measured as sensitive indicators of pollutant toxicity. However, little is known about the effects of environmental factors affect oxygen consumption rate and ammonia excretion rate ofSaccostreacucullata. The effects of water temperature (13, 18, 23, 28, 31, 34℃) and salinity (18, 23, 28, and 33) on oxygen consumption rate and ammonia excretion rate inSaccostreacucullatawere analyzed using experimental ecological methods. Oxygen consumption rate varied from 0.43 mg g-1h-1to 3.15 mg g-1h-1and the ammonia excretion rate varied from 72.62 μg g-1h-1to 1245.77 μg g-1h-1in different water temperature. Water temperature and salinity significantly affected oxygen consumption rate (F=60.440) and ammonia excretion rate (F=61.083); additionally, the combination of water temperature and salinity affected oxygen consumption rate (F=4.907) and ammonia excretion rateF=11.412). Temperature varied from 13 to 31℃ and salinity varied from 18 to 28, and the relationship between oxygen consumption or ammonia excretion rate (Y) and temperature (T) and salinity (S) can be represented by the equationY=a+bT+cS+dT×S. The relationship between oxygen consumption rate (Y) and water temperature (T) can be described by the regression equation:Y=aebT, wherearanges from 0.1378 to 0.2117, andbranges from 0.0757 to 0.0912. The relationship between ammonia excretion rate (Y) and water temperature (T) can also be described by the regression equation:Y=aebT, wherearanges from 20.364 to 30.634, andbranges from 0.1044 to 0.1341. The atomic ratio of O and N varied from 2.20 to 7.78, with a mean of 3.81 to maintain standard metabolism inSaccostreacucullata.
Saccostreacucullata; water temperature; salinity; oxygen consumption rate; ammonia excretion rate
公益性行業(yè)(農(nóng)業(yè))科研專項(xiàng)項(xiàng)目(201303047);寧波市科技計(jì)劃重大項(xiàng)目(2014C10018);歐盟第六框架項(xiàng)目(INCO-CT-2004-510706)
2015-07-27;
2016-06-13
10.5846/stxb201507271570
* 通訊作者Corresponding author.E-mail: zuiyou@163.com
焦海峰,鄭丹,嚴(yán)巧娜,尤仲杰,嚴(yán)小軍.溫度、鹽度及交互作用對(duì)僧帽牡蠣(Saccostreacucullata)呼吸排泄的影響.生態(tài)學(xué)報(bào),2017,37(2):692-699.
Jiao H F, Zheng D, Yan Q N, You Z J, Yan X J.Effects of water temperature and salinity on oxygen consumption rate and ammonia excretion rate ofSaccostreacucullata.Acta Ecologica Sinica,2017,37(2):692-699.
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2022年3期)2022-03-16 05:55:08
天津外國(guó)語(yǔ)大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
當(dāng)代陜西(2021年2期)2021-03-29 07:41:24
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
汽車工程學(xué)報(bào)(2017年2期)2017-07-05 08:13:02
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國(guó)塑料(2016年3期)2016-06-15 20:30:00
