金斑蝶成蟲行為學特征
2017-02-23 07:43:37唐宇翀陳曉鳴周成理
林業科學研究 2017年1期
唐宇翀,陳曉鳴,周成理
(1. 廣安職業技術學院,四川 廣安 638000; 2.中國林業科學研究院資源昆蟲研究所,云南 昆明 650224)
金斑蝶成蟲行為學特征
唐宇翀1,2,陳曉鳴2*,周成理2
(1. 廣安職業技術學院,四川 廣安 638000; 2.中國林業科學研究院資源昆蟲研究所,云南 昆明 650224)
[目的]為摸清金斑蝶成蟲的行為特征,[方法]在田間的網室內跟蹤觀察其飛行、取食、求偶、交配和產卵行為。[結果]表明:金斑蝶成蟲羽化主要集中在8:00—10:00,成蟲羽化當天基本處于停歇狀態;羽化第2天起至交配前,雌雄成蟲的日平均飛行次數及日平均飛行時間均逐漸增加,雄成蟲的日飛行高峰在13:00—15:00,雌成蟲的日飛行高峰期在9:00—11:00和15:00—17:00,雌雄成蟲的日平均飛行次數、平均飛行時間和日飛行規律的差異均不顯著(p>0.05);飛行次數與飛行時間均呈顯著正相關(雌蝶:r= 0.978,p<0.05;雄蝶:r=0.957,p<0.05)。金斑蝶雌雄成蟲羽化次日開始取食,自羽化第2天起,雌雄成蟲的日平均取食次數及日平均取食時間均逐漸增加,雄蟲的日取食活動高峰期在9:00—11:00,雌蟲日訪花高峰期在11:00—13:00和15:00—17:00,雌雄成蟲的日平均取食次數、平均取食時間和日取食規律的差異均不顯著(p>0.05);取食次數與取食時間均呈不顯著正相關(雌蝶:r= 0.949,p>0.05;雄蝶:r=0.853,p>0.05)。最早交配時間發生在羽化第4天,雌雄成蟲均可多次交配,交配主要發生在14:00—16:00。產卵時間主要在11:00—13:00,雌成蟲的產卵方式為散產,主要產卵于寄主植物葉片背面。[結論]金斑蝶成蟲期可分為運動器官成熟階段、補充營養和促進生殖器官發育階段、求偶和交配階段、產卵等4個階段,各階段內行為有交叉。
飛行行為;取食行為;求偶行為;交配行為;產卵行為
任何昆蟲的行為都具有周期性的節律特征[1],因此,掌握昆蟲行為活動節律,有利于更好地開展害蟲防治或益蟲保護工作。然而,目前關于蝴蝶成蟲行為的系統研究較少,僅王翻艷等[2]和Li等[3]分別對觀賞蝴種大帛斑蝶(Idealeuconoe(Erichson))和青斑蝶(Tirumalalimniace(Cramer))成蟲的飛行、訪花、求偶、交配和產卵行為作了系統觀察。其余關于蝶類成蟲行為的研究國內主要圍繞一些瀕危或保護蝶種的取食、交配或產卵等部分行為展開,以期通過對行為的研究找出影響其繁殖的因素或野生種群保護的措施[4-7]。國外則側重于從化學生態學角度尋找吸引蝴蝶取食[8-11]、交配[12-13]或產卵的化合物[14-15]以及研究對蝴蝶種群繁衍至關重要的產卵行為[16]。
金斑蝶(Danauschrysippus(Linnaeus))為鱗翅目(Lepidoptera)斑蝶科(Danaidae)斑蝶屬(DanausKluk),國內分布于海南、云南、廣東、廣西、福建、臺灣、四川、江西、湖北和陜西等地[17]。成蟲喜訪馬利筋,性情溫和,飛行低緩,姿態優雅,非常適應人工繁殖園內小生境。對經濟和園林植物無害,是集工藝、生態觀賞和喜慶放飛三用蝶種,具較高的開發利用價值,在南方各地廣為養殖。在云南元江地區,1年可發生10代以上[18]。
在金斑蝶的行為方面,王文明[7]研究了燕山地區金斑蝶的生物學特征,并對其交尾節律和產卵行為做了詳細觀察,發現6—7月為其適宜的繁殖月份。Tang 等[19]通過對金斑蝶訪花行為選擇發現,其在訪花過程中,以視覺信號為主,嗅覺信號為輔;但關于金斑蝶成蟲行為學研究還未見系統報道。作為優良的三用蝶種,系統地觀察金斑蝶成蟲的飛行、取食、交配和產卵行為,不僅可掌握其成蟲的活動特點與規律,了解成蟲的生物學習性,還可對金斑蝶的人工規模化繁育及野生種群的保護提供理論支撐,并為金斑蝶飛舞景觀營造提供理論依據。
1 材料與實驗場地
1.1 材料
供試蝴蝶為人工養殖的臨近羽化的金斑蝶蛹。蜜源植物為馬利筋。
1.2 實驗場地
中國云南元江縣干熱河谷區中國林業科學研究院資源昆蟲研究所試驗站,海拔約 400 m,實驗在網室內(4 m×8 m×4 m)進行。網室透光良好,光照均勻。
2 研究方法
參照周成理[5]對枯葉蛺蝶成蟲習性的觀察方法和Li等[3]對青斑蝶成蟲行為的研究方法,于6月中旬每日6:30—19:00在田間的網室內持續觀察金斑蝶成蟲的羽化、飛行、取食、求偶、交配和產卵行為,并記載其羽化、飛行和取食時間。
2.1 羽化
將掛有臨近羽化的金斑蝶蛹的塑料盆放入網室內的支架上。次日早晨成蟲羽化前(6:30)開始觀察記錄成蟲的羽化時間。
2.2 飛行

2.3 取食
跟蹤觀察標記成蟲的日取食起始時間及其取食行為。
2.4 求偶和交配
跟蹤觀察標記成蟲的交配起始時間及其求偶和交配行為。
2.5 產卵
跟蹤觀察標記成蟲的產卵行為以及產卵地點和方式。
同時通過攝像機記錄其余放飛的金斑蝶,作為行為分析的補充。網室中央集中擺放處于盛花期的馬利筋盆栽16盆。
2.6 數據分析與處理
采用 SPSS23.0 統計分析軟件對數據進行單因素方差分析和相關分析。
3 結果與分析
3.1 羽化
金斑蝶羽化時,首先是蛹體背部裂開,然后成蟲的胸足慢慢從裂口伸出,逐漸爬出蛹殼,接著用足抓住蛹殼,倒掛身體。隨著身體的倒掛,翅膀由皺縮不斷變大變硬,并伴有數滴黑褐色液體—蛹便從腹部排出。約10 min后翅膀完全舒展開,但成蟲不立即飛行,而是在原處抖動拍打翅膀或向上爬行。供試金斑蝶成蟲羽化均在上午完成,最早羽化時間為6:46,主要集中在8:00—10:00。雄蝶羽化時間早于雌蝶,但在羽化時間分布上雌蝶更為集中。
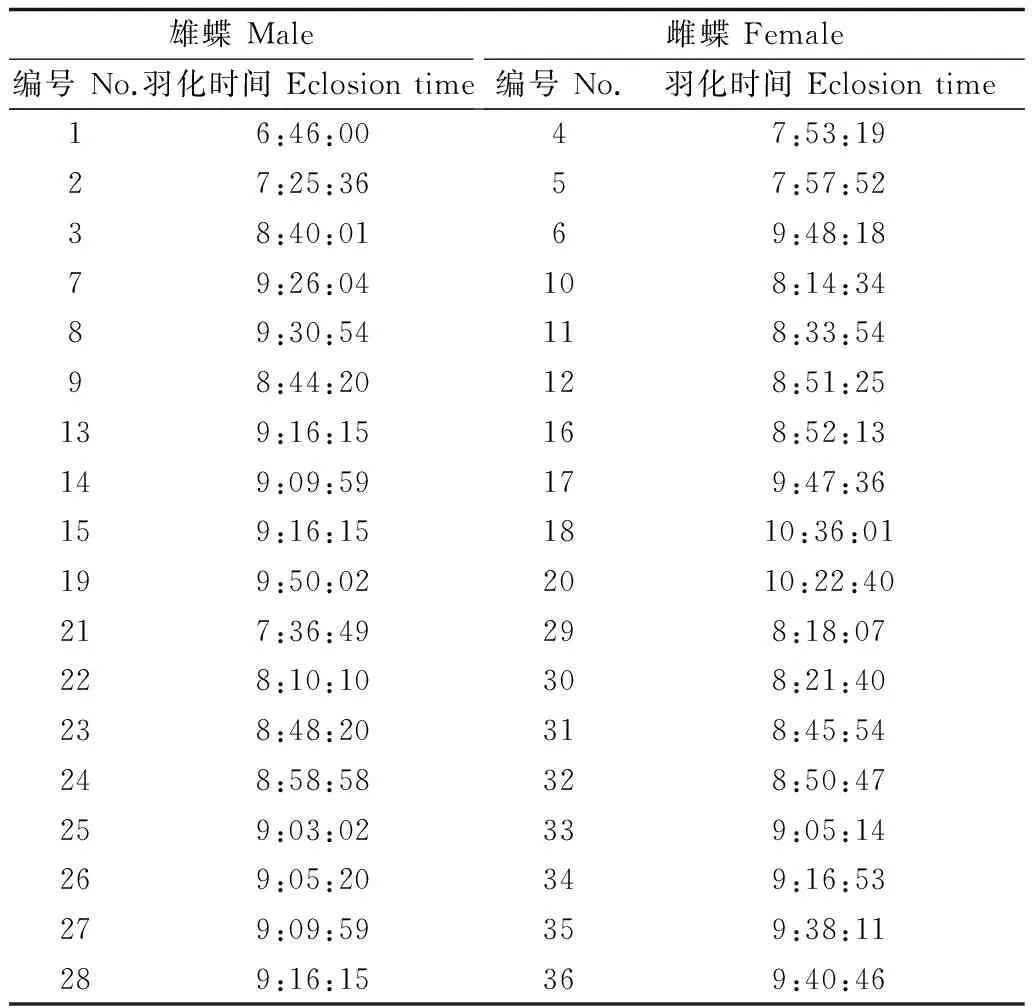
表1 金斑蝶羽化時間Table 1 The eclosion time of Danaus chrysippus
3.2 飛行行為
成蟲羽化當天基本處于停歇狀態,偶有幾次轉換停歇位置在網室壁上的爬行行為。自羽化第2天起,雌雄成蟲的日平均飛行次數及日平均飛行時間均逐漸增加(圖1)。金斑蝶的雌雄成蟲在7:00之前就開始少量的飛行活動,雄成蟲的日飛行高峰在13:00—15:00,雌成蟲的日飛行高峰為9:00—11:00和15:00—17:00(圖2)。金斑蝶在羽化后至產卵前,雌蟲飛行總次數為176次,飛行總時間為84.90 min,平均每天飛行44次,飛行21.23 min,單次平均飛行時間0.48 min;雄蟲的飛行總次數為99次,飛行總時間為54.10 min,平均每天飛行25次,飛行13.53 min,單次平均飛行時間0.54 min。雌雄成蟲的日平均飛行次數、平均飛行時間和日飛行規律均無顯著差異(p>0.05);飛行次數與飛行時間均呈顯著正相關(雌蝶:r=0.978,p<0.05;雄蝶:r=0.957,p<0.05)。
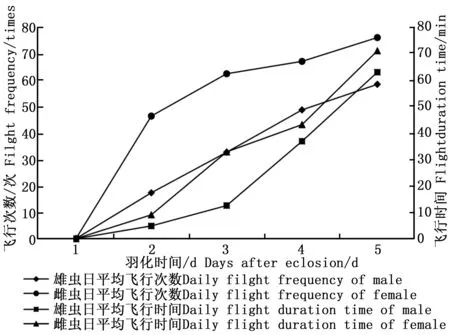
圖1 雌雄成蟲羽化后至產卵前的日飛行次數與時間Fig.1 The daily flight frequency and duration time of female and male from eclosion to oviposition
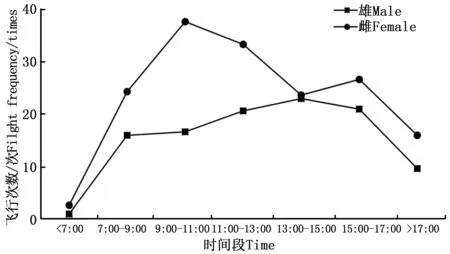
圖2 雌雄成蟲羽化后至產卵前的日飛行節律Fig.2 The daily flight rhythm of female and male from eclosion to oviposition
3.3 取食行為
觀察發現,金斑蝶雌雄成蟲羽化當日均不取食,次日才開始取食。自羽化第2天起,雌雄成蟲的日平均取食次數及日平均取食時間均逐漸增加(圖3)。
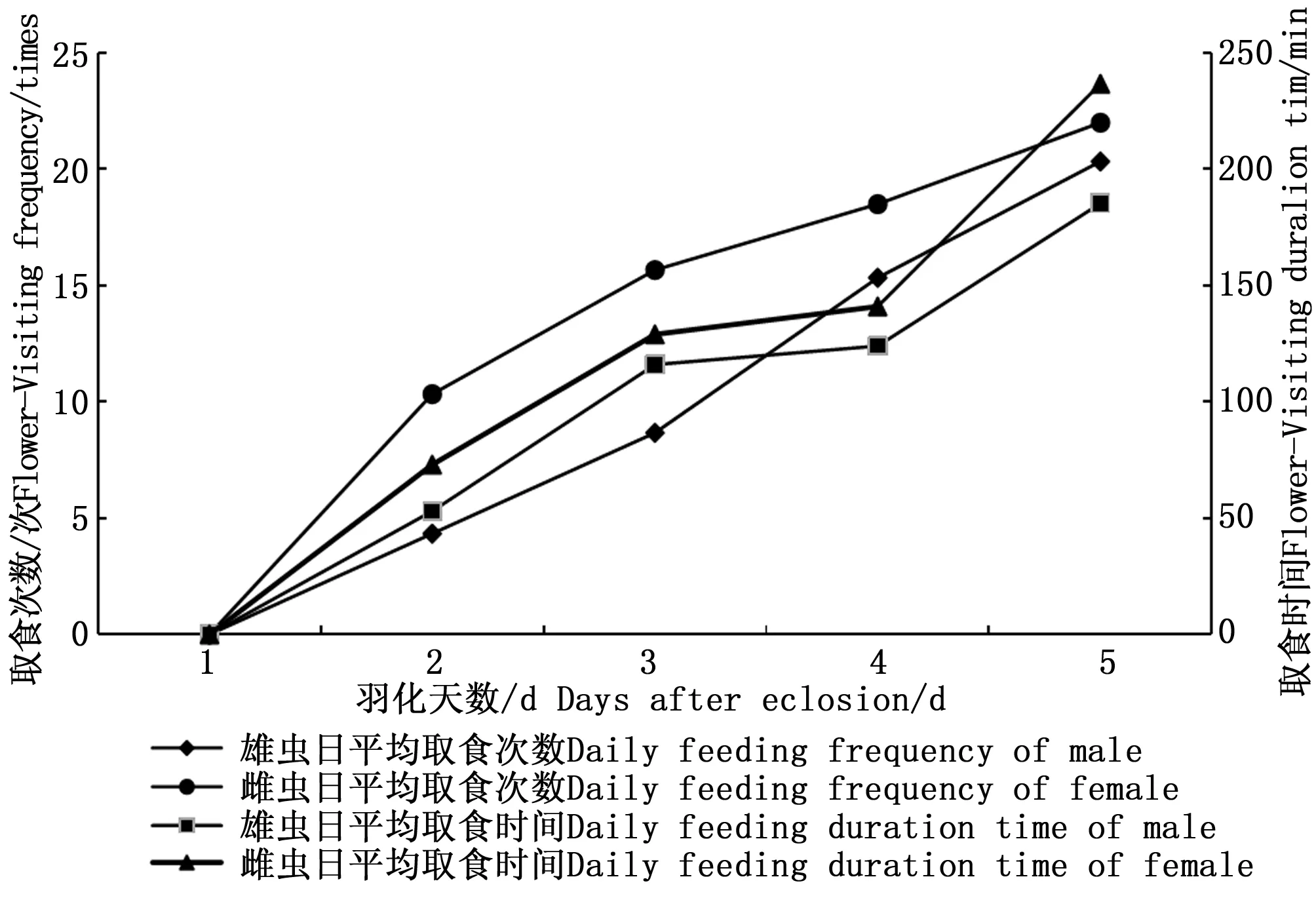
圖3 雌雄成蟲羽化后至產卵前的日取食次數與時間Fig.3 The daily feeding frequency and duration time of female and male from eclosion to oviposition
金斑蝶接近7:00開始取食活動,之后取食次數逐漸增多,交配前,雄蟲的日取食活動有一個高峰期在9:00—11:00,之后取食次數下降;雌蟲有2個取食高峰期,分別在11:00—13:00和15:00—17:00,17:00后取食次數逐漸減少,直至停止活動(圖4)。
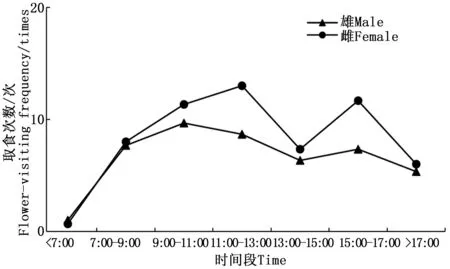
圖4 雌雄成蟲羽化后至產卵前的日取食節律Fig.4 The daily feeding rhythm of female and male from eclosion to oviposition
金斑蝶在羽化后至產卵前,雌蟲的取食總次數為134次,取食總時間為343.00 min,平均每天取食34次,取食85.75 min,單次平均取食時間2.52 min;雄蟲的取食總次數為85次,取食總時間為293.00 min,平均每天取食21次,取食73.25 min,單次平均取食時間3.49 min。
雌雄成蟲的日平均取食次數、平均取食時間和日取食規律均無顯著差異(p>0.05);取食次數與取食時間均無顯著正相關(雌蝶:r= 0.949,p>0.05;雄蝶:r=0.853,p>0.05)。
3.4 求偶、交配與產卵行為
金斑蝶雄成蟲的交配求偶屬于巡游型,雄成蟲在網室內巡回飛行,主動尋找雌成蟲交配,當遇到有交配意愿的雌成蟲時,雙雙在空中漫游,然后雌成蟲停落在網室壁、地面或植物枝葉上,緊接著雄成蟲也停落在雌成蟲旁并將腹部伸出彎曲至雌成蟲腹末開始交配。
最早交配時間發生在羽化第4天。雌雄成蟲均可多次交配,交配主要發生在14:00—16:00;最早交配開始于11:03,止于16:00—17:00,最晚開始于17:56,過夜后于次日6:30前結束。在觀察到的4對成蟲共7次交配時間中,交配持續時間最短16 min,最長357 min,平均交配時間(174.71±152.07) min。
大多數雌成蟲交配次日便開始產卵,也有個別雌成蟲交配當天即開始產卵。產卵時,雌成蟲搜尋到產卵地后首先停落在要產卵的部位上,然后用腹部接觸摩擦幾次產卵部位,最后將腹部彎曲,產出卵1枚。產卵最早開始于8:31,最晚止于18:12,產卵時間主要在11:00—13:00。金斑蝶雌成蟲的產卵方式為散產,主要產卵于寄主植物葉片背面,其次為莖桿,少數產于寄主植物花朵、寄主植物葉片正面、雜草和花盆上。
4 討論
蝶類成蟲經過羽化后才具備飛行能力,目前蝶類的羽化節律已有較多報道,且結果表明不同種類之間的羽化時間有差異,多數發生在4:00—6:00和上午[20-24]。本研究中,金斑蝶最早羽化時間為6:46,主要集中在8:00—10:00,與王文明[7]對燕山地區金斑蝶的羽化節律相吻合;在羽化時間上,雄蝶羽化時間早于雌蝶,這也符合在外界條件相同的情況下,雄蝶比雌蝶先羽化的研究結果[20, 23, 25-26]。金斑蝶與枯葉蛺蝶、大帛斑蝶和青斑蝶一樣[2-3, 5],羽化當天基本不活動,為停歇狀態,說明其處于運動器官成熟階段。通過飛行肌的鍛煉,待其發育成熟后為飛行做準備[27]。
飛行是蝶類開展其他活動的前提,金斑蝶也不例外,其成蟲的飛行活動與取食、交配和產卵等行為密切相連,不取食、交配或產卵時,成蟲基本都處于停歇狀態。蝴蝶為變溫動物,通過外界溫度調節蝴蝶飛行期身體的溫度影響其飛行能力[8],從而影響其在飛行、繁殖、休息、取食的時間分配[28]。蝴蝶在陰雨天一般停止飛行[29]。金斑蝶的雌雄成蟲在7:00之前就開始少量的飛行活動,雄成蟲的日飛行高峰在13:00—15:00,雌成蟲的日飛行高峰期為9:00—11:00和15:00—17:00。四川絹蝶(Parnassiidaszechenyii(Frivaldszk))10點開始有少量活動,12:00—13:00達到高峰[29];曲紋紫灰蝶(Chiladespandava(Horsfield))7:30開始少量活動,9:30—10:30達到日活動高峰[30]。夏季,枯葉蛺蝶(Kallimainachus(Doyére))大多在11:00—13:00 和15:00—17:00活動[5]。說明不同蝶種、不同性別和不同環境溫度都可以對蝴蝶的飛行產生影響[31]。
昆蟲的訪花活動節律主要受環境溫度、昆蟲自身的生理特征、蜜源植物等多種因素的影響,因此,不同蝴蝶的訪花時間和訪花高峰期有所差異[29, 32]。本研究中,從羽化后到交配前金斑蝶的取食次數逐漸增加,與大帛斑蝶和青斑蝶類似[2-3],說明其同樣處于補充營養和促進生殖器官發育階段。通過補充營養不僅能促進生殖系統的發育成熟,還能為日常生命活動提供活動能量以及延長成蟲的產卵期,對于蝴蝶生殖具有重要意義[33-34]。
蝴蝶的求偶有2種類型:巡游型和等候型[18]。金斑蝶與大帛斑蝶、青斑蝶、四川絹蝶[2-3, 8]都屬于巡游型,枯葉蛺蝶為等候型[5],說明不同蝶種的求偶方式有差異,巡游型主要見于鳳蝶科和粉蝶科[18]。王文明[7]對燕山地區金斑蝶的生物學研究表明,交配發生在10:00—16:00,時長(94.6±22.39) min,交配后雌蟲將大部分卵散產于寄主植物葉背面。在元江地區,金斑蝶的羽化規律、產卵方式和產卵部位與燕山地區相同,但成蟲交配發生在14:00—16:00,平均交配時間為(174.71±152.07) min,這可能與元江的氣溫較燕山地區高,蝴蝶成蟲活動受溫度影響有關。一旦出現求偶和交配行為,成蟲即進入求偶和交配階段。
交配完成后,成蟲進入產卵階段。多數情況下,蝴蝶將卵主要產于寄主植物的嫩葉嫩梢的背面,有利于減少卵受日曬、雨淋以及天敵的侵害[35],如玉帶鳳蝶(Papiliopolytes(Linnaeus))、青斑蝶、大帛斑蝶[2-3, 24],有的種類則喜產卵在寄主植物附近或寄主植物葉片正面,如枯葉峽蝶[5]。雌蝶的產卵方式因蝶種不同有所差異,金斑喙鳳蝶(Teinopalpusaureus(Mell))、青斑蝶、大帛斑蝶、枯葉蛺蝶等為散產,絲帶鳳蝶(Sericinusmontelus(Gray))、紅鋸蛺蝶(Cethosiabiblis(Drury))、白帶鋸蛺蝶(C.cyane(Drury))和文蛺蝶(Vindulaerota(Fabricius))等為聚產[18]。散產可有效避免后代對食物的競爭,聚產則可發揮集體防御功能。因此,不管何種產卵方式,均是成蟲為使后代成功繁衍所采取的適應性措施。
5 結論
[1] 吳少會,向 群,薛芳森. 昆蟲的行為節律[J]. 江西植保,2006,29(4):147-157.
[2] 王翻艷,李承哲,陳曉鳴,等. 大帛斑蝶成蟲行為學特征[J]. 生態學報,2015,35(23):1-8.
[3] Li C Z, Wang F Y, Chen X M,etal. Adult Behavior ofTirumalalimniace(Lepidoptera: Danaidae) [J]. Journal of Insect Science, 2015, 15(1): 1-7.
[4] 曾菊平,周善義,丁 健,等. 瀕危物種金斑喙鳳蝶的行為特征及其對生境的適應性[J]. 生態學報,2012,32(20):6527-6534.
[5] 周成理. 枯葉蛺蝶生物學特征及種內分化的分子遺傳研究[D]. 北京:中國林業科學研究院,2008.
[6] 包正云. 青海祁連山地區四川絹蝶的訪花行為和繁殖生態學研究[D]. 西寧:青海師范大學,2010.
[7] 王文明. 中華虎鳳蝶和金斑蝶在燕山地區的生物學特性研究[D]. 南昌:南昌大學,2011.
[8] Andersson S, Dobson H E M. Behavioral foraging responses by the butterflyHeliconiusmelpomenetoLantanacamarafloral scent [J]. Journal of Chemical Ecology, 2003, 29(10): 2303-2318.
[10] Lewis Z, Wedell N. Effect of adult feeding on male mating behaviour in the butterfly,Bicyclusanynana(Lepidoptera: Nymphalidae).[J]. Journal of Insect Behavior, 2007, 20(2): 201-213..
[12] Costanzo K, Monteiro A. The use of chemical and visual cues in female choice in the butterflyBicyclusanynana[J]. Proceedings Biological Sciences, 2007, 274(1611): 845-851.
[13] Nishida R, Schulz S, Kim C S,etal. Male sex pheromone of a giant danaine butterfly,Idealeuconoe[J]. Journal of Chemical Ecology, 1996, 22(5): 949-972.
[14] Bruinsma M, Van Dam N M, Van Loon J J A,etal. Jasmonic acid-induced changes inBrassicaoleraceaaffect oviposition preference of two specialist herbivores [J]. Journal of chemical ecology, 2007, 33(4): 655-668.
[15] Stefanescu C, Penuelas J, Sardans J,etal. Females of the specialist butterflyEuphydryasaurinia(Lepidoptera: Nymphalinae: Melitaeini) select the greenest leaves ofLoniceraimplexa(Caprifoliaceae) for oviposition [J]. European Journal of Entomology, 2006, 103(3): 569-574.
[16] Catta-Preta P D, Zucoloto F S. Oviposition behavior and performance aspects ofAsciamonuste(Godart, 1919) (Lepidoptera, Pieridae) on kale (Brassicaoleraceavar. acephala) [J]. Revista Brasileira de Entomologia, 2003, 47(2): 169-174.
[17] 周 堯. 中國蝶類志[M]. 鄭州:河南科學技術出版社,1994.
[18] 陳曉鳴,石 雷,周成理,等.中國觀賞蝴蝶[M]. 北京:中國林業出版社,2008.
[19] Tang Y C, Zhou C L, Chen X M,etal. Visual and Olfactory Responses of Seven Butterfly Species During Foraging[J]. Journal of Insect Behavior, 2013, 26(3): 387-401.
[20] 張軍生,滕文霞,康爾年. 紅線蛺蝶生物學特性初步研究[J]. 內蒙古林業科技,2002(1):20-22.
[21] 佘德松,馮福娟. 木蘭青鳳蝶生物學特性研究[J]. 中國森林病蟲,2003,22(6):17-20.
[22] 周成理,史軍義,易傳輝,等. 枯葉蛺蝶Kallimainachus的生物學研究[J]. 四川動物,2005,24(4):445-450.
[23] 任志勇,朱道弘,王洪建. 花椒鳳蝶生物學特性觀察及防治試驗初報[J]. 甘肅林業科技,2007,32(4):54-56.
[24] 廖 宇. 玉帶鳳蝶生物學特性研究[D]. 福州:福建農林大學, 2009.
[25] 袁德成,買國慶,薛大勇,等. 中華虎鳳蝶棲息地、生物學和保護現狀[J]. 生物多樣性,1998,6(2):105-115.
[26] 羅志文,李世震,李春豐,等. 絲帶鳳蝶的生物學特性研究初報[J]. 佳木斯大學學報:自然科學版,2005,23(3):437-442.
[27] Fernanda C F, Carminda C L. Differential flight muscle development in workers, queens and males of the eusocial bees,ApismelliferaandScaptotrigonapostica[J]. Journal of Insect Science, 2010, 10(85): 1-9.
[28] Hirota T, Hamano K, Obara Y. The Influence of Female Post-Emergence Behavior on the Time Schedule of Male Mate-Locating inPierisrapaecrucivora[J]. Zoologicalence, 2001, 18(4): 475-482.
[29] 霍曉杰. 青海祁連山地區四川絹蝶的生物學特性及各因素對其活動影響的研究[D]. 西寧:青海師范大學,2010.
[30] 劉光華, 陸永躍, 甘詠紅,等. 曲紋紫灰蝶的生物學特性和發生動態研究[J]. 應用昆蟲學報,2003,40(5):426-428.
[31] 王翻艷. 大帛斑蝶成蟲行為學觀察及其求偶機制研究[D]. 北京:中國林業科學研究院,2015.
[32] 邵長芬,關文靈,李得發,等. 大葉醉魚草訪花昆蟲行為與活動規律[J]. 亞熱帶植物科學,2014(3):227-231.
[33] O’Brien D M, Boggs C L, Fogel M L. Making eggs from nectar: connections between butterfly life history and the importance of nectar carbon in reproduction[J]. Oikos, 2004, 105(2): 279-291.
[34] Mevi-Schütz J, Erhardt A. Amino acids in nectar enhance butterfly fecundity: a long-awaited link[J]. American Naturalist, 2005, 165(4): 411-419.
[35] Baguette M, Schtickzelle N. Local population dynamics are important to the conservation of metapopulations in highly fragmented landscapes[J]. Journal of Applied Ecology, 2003, 40(2): 404-412.
(責任編輯:張 玲)
Adult Behavior Feature ofDanauschrysippus
TANGYu-chong1,2,CHENXiao-ming2,ZHOUCheng-li2
(1.Guang’an Vocational and Technical College, Guang’an 638000, Sichuan, China;2.Research Institute of Resource Insects, Chinese Academy of Forestry, Kunming 650224, Yunnan, China)
[Objective]To study the behavior characteristics of the adultDanauschrysippus. [Method]The adult butterfliesD.chrysippuswere observed by tracking the flying, feeding, courtship, mating and ovipositing behavior in the field of the net-house.[Result]The eclosion mainly occurred at the 8:00—10:00. The adults were in the stop or rest status at the day of emergence. The daily average flying time and frequencies gradually increased from the second day of eclosion to the day of mating. The flying peak time of males were at 13:00—15:00, while two flying peak were observed in females at 9:00—11:00 and 15:00—17:00. There was no significant difference between male and female adults in the daily average flight number, times and the daily flight rule (p>0.05). However, there was a significant positive correlation between the number of flights and the time of flight in both sex (p<0.05). The feeding behavior began at the second day of eclosion. The daily average feeding time and frequencies also gradually increased. The daily feeding activity peak of male is 9:00—11:00, on the female flower visiting peak is 11:00—13:00 and 15:00—17:00. There was no significant difference between male and female adults in the daily average feeding number, times and the daily feeding rule (p>0.05), and no significant positive correlation was found between the feeding frequency and feeding time in both sex (p>0.05). The first mating behavior occurred at the fourth day after eclosion. The male and female can mate more than once in their whole lives. Mating occurred mainly at 14:00—16:00, while egg-laying occurred mainly at 11:00—13:00. Most of eggs were deposited in a dispersed manner on the back of host leaves. [Conclusion]The adult stage ofD.chrysippuscan be divided into four, i.e. motion organ mature stage, nutritional supplements and promote reproductive organ development stage, courtship and mating stage, and oviposition stage. In each stage, the behavior has a cross.
flight behavior;feeding behavior;courtship behavior;mating behavior;egg-laying behavior
10.13275/j.cnki.lykxyj.2017.01.018
2016-05-18
林業公益性行業科研專項經費(201504305);云南省社會發展重大項目:大理蝴蝶泉蝴蝶飛舞自然景觀構建重現技術研究及應用(2-351);四川省教育廳一般項目(15ZB0398);廣安職業技術學院院級課題(KT-2016-20)
唐宇翀,博士,講師,從事園林植物病蟲害防治和資源昆蟲研究. E-mail:tangyuchong@126.com
* 通訊作者.
S899
A
1001-1498(2017)01-0131-06
