基于MaxEnt的西藏飛蝗在中國的適生區(qū)預測
2017-03-09 08:28:38王茹琳封傳紅石朝鵬
生態(tài)學報 2017年24期
王茹琳,李 慶,封傳紅,石朝鵬
1 四川農(nóng)業(yè)大學農(nóng)學院,成都 611130 2 四川省農(nóng)村經(jīng)濟綜合信息中心,成都 610072 3 四川省農(nóng)業(yè)廳植物保護站,成都 610041 4 山東省植物保護總站,濟南 250000
全世界約有蝗蟲12000余種,除南極洲外各大洲均有分布。飛蝗(LocutsamigratoriaL.)是世界范圍內(nèi)的重要害蟲,已知有1個種10個亞種,其中西藏飛蝗(LocustamigratoriatibetensisChen)為青藏高原特有且分布海拔最高的亞種[1],主要分布在西藏、四川及青海等省區(qū)[2],主要以青稞、牧草及雜草為食[3- 5]。據(jù)史料記載,西藏飛蝗曾在青藏高原多次成災,蝗災所到之處莊稼顆粒無收。20世紀90年代以來,西藏飛蝗在四川甘孜州和阿壩州,西藏普蘭、昌都、阿里及青海玉樹等地曾大面積爆發(fā),尤以2003—2006年該蟲發(fā)生極為嚴重,特別是雅魯藏布江、雅礱江以及金沙江等橫斷山脈河谷地帶,蝗災的爆發(fā)對當?shù)剞r(nóng)牧業(yè)正常生產(chǎn)造成極大破壞,導致難以估量的經(jīng)濟損失[6]。
預測物種適生分布是生態(tài)學的重要研究領域,物種分布模型(species distribution models,SDM)逐漸發(fā)展并成為研究物種適生性的一個重要工具[7]。此類模型根據(jù)物種已知的“存在”或“不存在”數(shù)據(jù),結(jié)合相應的環(huán)境變量,利用一定的數(shù)學理論計算物種在目標地區(qū)的分布概率,量化環(huán)境變量與物種分布之間的關系,尋找物種分布的限制因子及生境偏好等。目前,物種分布模型的常用領域主要為:預測物種潛在分布區(qū)、物種分布與氣候關系、瀕危及珍貴物種適生性預測和古生物地理研究等方面[8- 10],并起到了良好的效果[11]。目前常用的物種分布模型中,最大熵模型MaxEnt(maximum entropy model)的模擬精度要高于其他模型,加之該軟件具有運行時間短、操作簡便、運行結(jié)果穩(wěn)定和所需樣本量小等特點在業(yè)內(nèi)得到了廣泛的認可[12- 13],已被應用于多種病蟲害適生性的模擬預測,如柑橘黃龍病菌(CandidatusLiberibacterasiaticus)、葉緣焦枯病菌(Xylellafastidiosa)、柑橘木虱(DiaphorinacitriKuwayama)和稻水象甲(LissorhoptrusoryzophilusKuschel)等[14- 27],且模擬的結(jié)果與物種實際分布的吻合度高。
目前對西藏飛蝗的研究主要集中在生物學特性、環(huán)境脅迫、數(shù)量性狀及遺傳特征等方面[28],還未發(fā)現(xiàn)對其生態(tài)位預測的相關研究報道。本研究基于MaxEnt模型,研究西藏飛蝗分布與環(huán)境變量之間的聯(lián)系,揭示影響其分布的主導氣候因子,分析西藏飛蝗在中國的潛在分布區(qū),旨在為制定合理的防控措施提供重要參考和理論依據(jù)。
1 材料與方法
1.1 西藏飛蝗的分布數(shù)據(jù)與處理
MaxEnt模型模擬需要物種分布數(shù)據(jù)和環(huán)境數(shù)據(jù),分布數(shù)據(jù)即為“存在”點的經(jīng)緯度信息。西藏飛蝗的分布數(shù)據(jù)通過以下方式獲得:(1)實地考察,四川境內(nèi)的采樣點多為實地考察獲得,調(diào)查點主要位于西藏飛蝗高發(fā)區(qū)的甘孜州和阿壩州,發(fā)現(xiàn)西藏飛蝗若蟲或成蟲即記為“存在”,GPS記錄經(jīng)緯度。(2)查詢物種分布數(shù)據(jù)庫。本研究查詢的數(shù)據(jù)庫包括“國際農(nóng)業(yè)與生物科學中心(CABI,http://www.cabi.org/)”數(shù)據(jù)庫、“全球物種多樣性信息庫(GBIF,http://www.gbif.org/)”、中國西南地區(qū)動物資源數(shù)據(jù)庫(http://www.swanimal.csdb.cn)、教學標本資源共享平臺(http://mnh.scu.edu.cn/)。(3)檢索國內(nèi)外公開發(fā)表的西藏飛蝗相關的期刊論文。數(shù)據(jù)處理:采用緩沖區(qū)分析法對獲得的分布點進行校對、篩選,排除空間關聯(lián)性較大造成的過擬合模擬的影響,由于環(huán)境變量空間分辨率為2.5 arc-minutes(約4.5 km2),故設置緩沖半徑為1.5 km,當分布點之間距離小于3 km時,只保留其中一點,共獲得分布點47個,通過Google earth確定每個分布點的經(jīng)緯度。參照MaxEnt軟件操作手冊,將經(jīng)緯度坐標輸入Excel,保存格式為*.CSV,字段包括物種名稱、經(jīng)度和緯度[29]。由于西藏飛蝗生活在交通不便、氣候惡劣的高海拔地區(qū),所獲得的分布點很難完全覆蓋其實際分布。研究表明,在樣本量很小的情況下,MaxEnt模型較其他模型預測效果更好、準確性更高,更適用于小樣本數(shù)據(jù)的預測。
1.2 預測環(huán)境變量
1.2.1 環(huán)境變量數(shù)據(jù)來源
本研究所用的環(huán)境因子均來源于Worldclim氣候數(shù)據(jù)庫,數(shù)據(jù)是以全球的氣象記錄信息為基礎數(shù)據(jù),整合插值生成的全球氣候柵格數(shù)據(jù)[30]。利用ArcGIS軟件提取研究區(qū)(西藏、四川、云南、貴州和青海)19個生物氣候因子(具有較強的生物學意義,可反映溫度與降水的特點及季節(jié)性變化特征)、月平均降水量、月平均最高及最低氣溫等氣候指標數(shù)據(jù)。另外還選取了海拔數(shù)據(jù)作為地形因子(表1)。上述數(shù)據(jù)的空間分辨2.5 arc-minutes(約4.5 km2)。

表1 西藏飛蝗潛在地理分布的評價指標
1.2.2 影響西藏飛蝗潛在分布關鍵環(huán)境變量的篩選
影響物種分布的環(huán)境因子多種多樣,包括氣候因子、土壤因子、植被分布等,本文中選取了worldclim提供的24個環(huán)境變量構(gòu)建初始模型。初始模型構(gòu)建過程中選擇MaxEnt軟件中的刀切法(jackknife test)檢驗來測定環(huán)境變量對模型預測的貢獻大小,以此剔除對MaxEnt模型預測結(jié)果貢獻較小的環(huán)境變量,對貢獻較大的關鍵限制因子進行Spearman相關分析,提高模型模擬的精度,在此基礎上重建西藏飛蝗分布的最大熵模型,并對模擬結(jié)果進行準確性評價[31]。
刀切法檢驗使用MaxEnt軟件,Spearman相關分析使用Spss分析軟件。
1.3 MaxEnt模型的操作與結(jié)果評價
MaxEnt模型來源:選用基于最大熵理論的MaxEnt模型軟件來預測西藏飛蝗在中國的適生區(qū)分布,該軟件可在MaxEnt主頁(http://www.cs.princeton.edu/~schapire/maxent/)上免費獲取[32],當前最新版本為3.3.3k版。
MaxEnt模型的使用:將上述1.1中47個西藏飛蝗分布點數(shù)據(jù)和1.2中的環(huán)境因子數(shù)據(jù)添加到軟件中,隨機選取75%的西藏飛蝗分布點作為訓練集(training data)建立預測模型,剩余25%的西藏飛蝗分布點作為測試集(test data)驗證模型,選擇刀切法(jackknife test)測定各變量權(quán)重,選擇創(chuàng)建環(huán)境變量響應曲線,其余參數(shù)均選擇模型的默認值[33]。
本研究采用受試者工作特征曲線(Receiver Operating Characteristic Curve,ROC曲線)下面積值,即AUC值(Area Under Curve, AUC)為判據(jù)來評價模型模擬結(jié)果。ROC曲線,又稱為感受性曲線,該曲線根據(jù)一系列不同的二分類方式,分別以假陽性率(1-特異率)和真陽性率(1-遺漏率)為橫縱坐標繪制而成,曲線下面積值即AUC值,取值范圍為[0,1],AUC值越接近1表明環(huán)境變量與分布模型之間相關性越大,預測結(jié)果精度越高[34]。ROC曲線可使用Spss分析軟件獲得,本研究中MaxEnt模型可自動生成ROC曲線,并給出相應的AUC值。具體評價標準[35- 36]見表2。
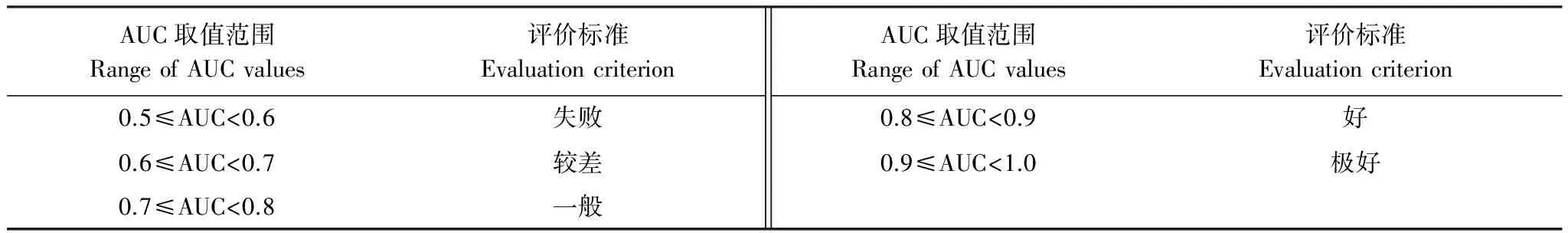
表2 AUC評價標準表
AUC:ROC曲線下面積值,即Area Under Curve
1.4 西藏飛蝗適生等級分區(qū)
MaxEnt模型輸出結(jié)果為西藏飛蝗在世界范圍內(nèi)的存在概率,數(shù)據(jù)為ASCⅡ格式,首先使用ArcGIS的ArcToolbox的格式轉(zhuǎn)換工具,將該數(shù)據(jù)轉(zhuǎn)為Raster格式,使該結(jié)果可在ArcGIS中顯示,而后使用“提取分析”功能得到該蟲在中國的存在概率分布圖[37- 38]。MaxEnt軟件模擬輸出的結(jié)果值在0—1之間,值越接近1表示物種越可能存在。參考IPCC報告[39]關于評估可能性的劃分方法,結(jié)合西藏飛蝗的實際情況,利用“Reclassify”功能,劃分分布值等級及相應分布范圍,并使用不同顏色表示,劃分標準為:存在概率<0.05為不適生區(qū);0.05≤存在概率<0.33為低適生區(qū);0.33≤存在概率<0.66為中適生區(qū);存在概率≥0.66為高適生區(qū)。
2 研究結(jié)果
2.1 主導環(huán)境因子篩選及模型檢驗
2.1.1 影響西藏飛蝗潛在分布主導環(huán)境變量篩選
MaxEnt模型預測過程中需使用物種空間分布點數(shù)據(jù)和環(huán)境變量數(shù)據(jù),并不是每個環(huán)境變量對物種潛在分布預測都是必需的,因此需要剔除對MaxEnt 模型預測結(jié)果貢獻較小的環(huán)境變量,篩選得到貢獻較大的關鍵限制因子[40]。本研究參考雷軍成等[41- 42]方法,利用MaxEnt軟件本身提供的模塊,即刀切法測定變量的重要性,通過比較每個環(huán)境變量對模型結(jié)果的影響程度來剔除影響較小的變量,最終獲得對模型模擬貢獻率較大的變量作為主導變量。本研究中,依據(jù)MaxEnt的運算結(jié)果,不同環(huán)境變量對西藏飛蝗潛在分布的累積貢獻率見表3。海拔高度為限制西藏飛蝗分布的主要變量,單因子貢獻率達到了52%,其次,8月份平均雨量、1月份平均雨量、等溫性、1月份平均溫度、12月份平均溫度和2月份平均溫度對西藏飛蝗潛在分布的累積貢獻率為42.6%。
本研究使用Spearman相關系數(shù)法計算7個環(huán)境因子之間的相關性,以消除共線性對模型建模過程和結(jié)果解釋的影響,環(huán)境因子之間的相關系數(shù)絕對值大于0.8[43- 44],則認為兩者具有很強的相關性。由表4可以看出,7個環(huán)境因子兩兩之間相關系數(shù)均未超過0.8,因此本研究選定上述7種環(huán)境變量作為影響西藏飛蝗分布的主導變量,在此基礎上重建西藏飛蝗在中國的分布的最大熵模型,并進行模擬結(jié)果的精度評價,以獲得最優(yōu)秀的預測模型。
表3影響西藏飛蝗分布的環(huán)境變量貢獻百分率和累積貢獻百分率
Table3TheaccumulatedcontributionofeachenvironmentalvariabletothepotentialdistributionofLocustamigratoriatibetensisdefinedbyMaxEnt
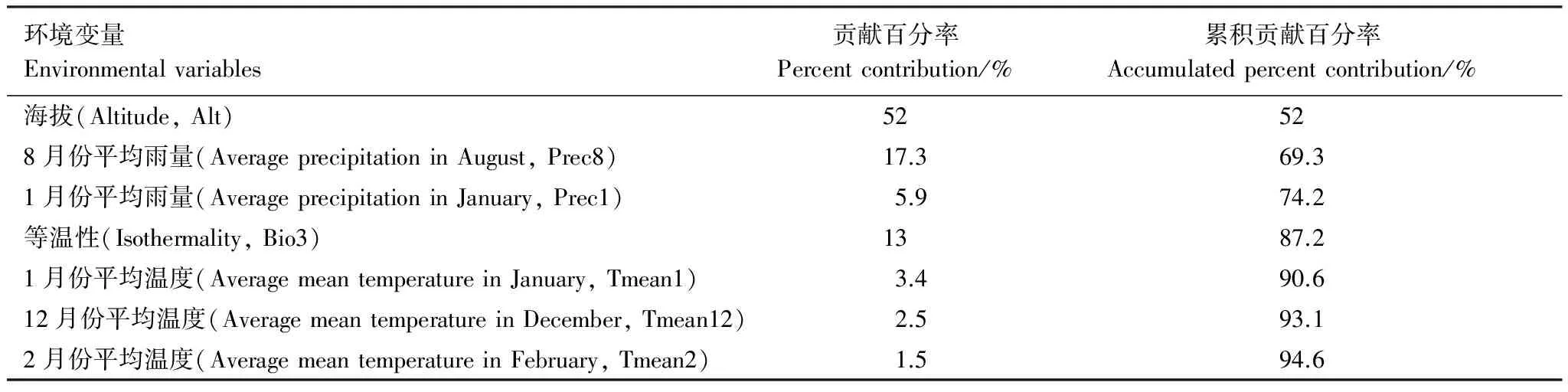
環(huán)境變量Environmentalvariables貢獻百分率Percentcontribution/%累積貢獻百分率Accumulatedpercentcontribution/%海拔(Altitude,Alt)52528月份平均雨量(AverageprecipitationinAugust,Prec8)17.369.31月份平均雨量(AverageprecipitationinJanuary,Prec1)5.974.2等溫性(Isothermality,Bio3)1387.21月份平均溫度(AveragemeantemperatureinJanuary,Tmean1)3.490.612月份平均溫度(AveragemeantemperatureinDecember,Tmean12)2.593.12月份平均溫度(AveragemeantemperatureinFebruary,Tmean2)1.594.6
累積貢獻百分率為貢獻百分率之和

表4 環(huán)境因子之間的Spearman相關系數(shù)
Alt:海拔,Altitude;Bio3:等溫性,Isothermality;Prec1:1月份平均雨量,Average precipitation in January;Prec8:8月份平均雨量,Average precipitation in August;Tmean1:1月份平均溫度,Average mean temperature in January;Tmean2:2月份平均溫度,Average mean temperature in February;Tmean12:12月份平均溫度,Average mean temperature in December;*表示在α=0.05水平上顯著相關
2.1.2 ROC曲線和AUC值對模型準確性檢驗
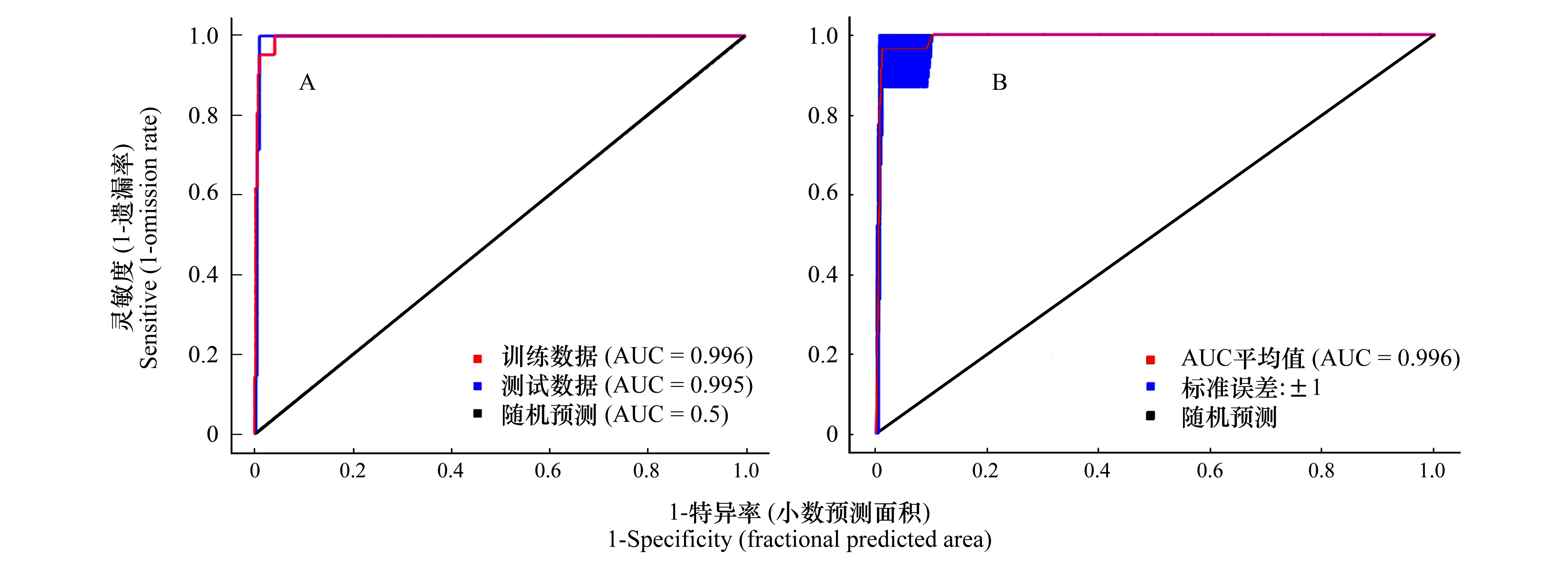
圖1 模型適用性檢驗AUC值Fig.1 AUC value of test on applicability of the MaxEnt modelA代表基于所有環(huán)境變量計算的ROC曲線,B代表基于主導環(huán)境變量ROC曲線
圖1是MaxEnt模型模擬輸出的ROC曲線圖,圖中表明基于所有環(huán)境變量和主導環(huán)境變量構(gòu)建的西藏飛蝗在中國的地理分布模型的AUC值分別為0.996和0.993,依據(jù)表1的AUC值評價標準表,本次構(gòu)建模型的預測準確性達到“極好”的標準,說明此結(jié)果可用于研究西藏飛蝗的適生區(qū)劃。
2.2 西藏飛蝗在中國的地理分布的預測
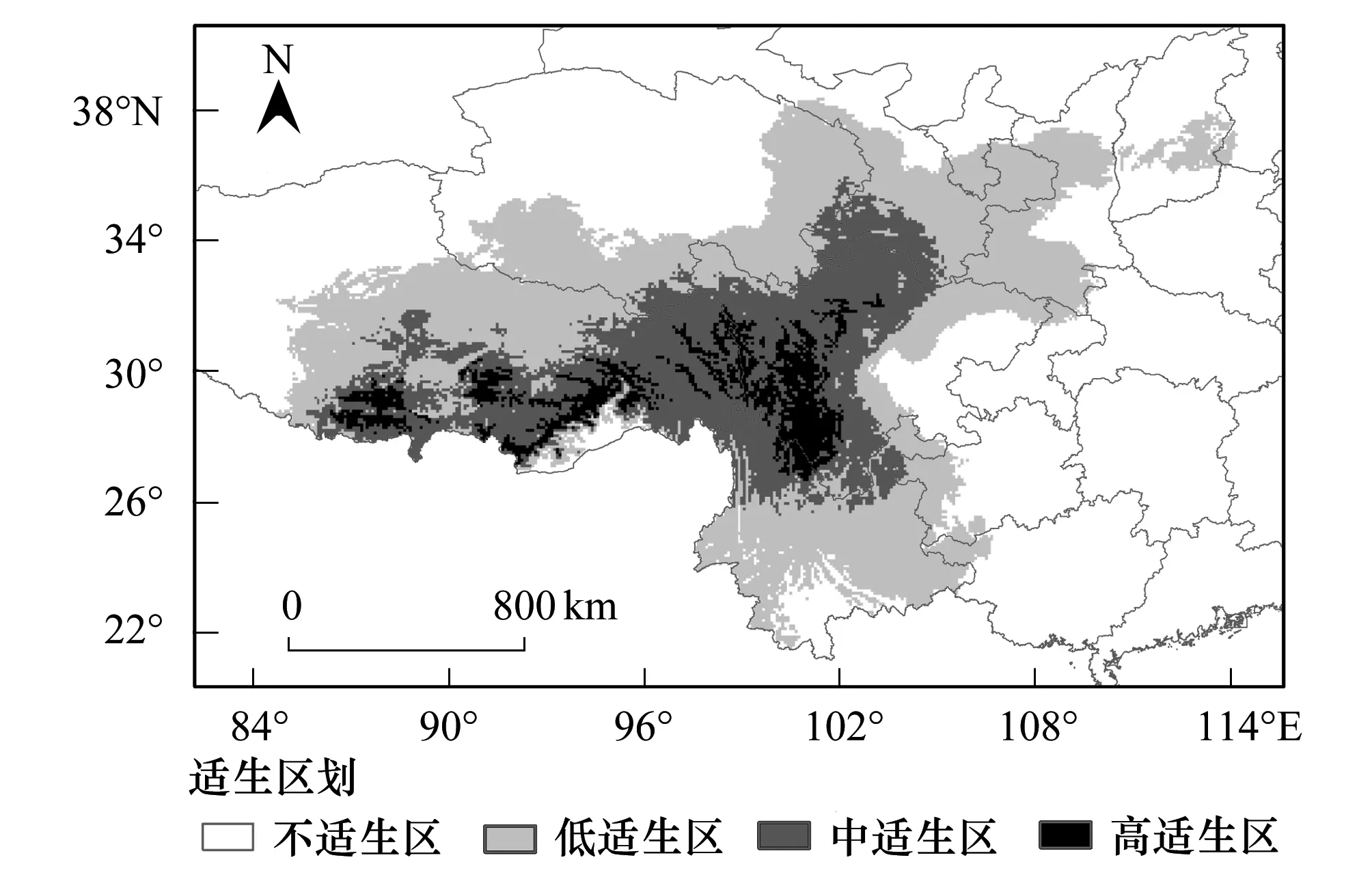
圖2 基于MaxEnt模型預測的西藏飛蝗在中國的適生分析圖 Fig.2 Potential suitable distribution of Locusta migratoria tibetensis in China based on MaxEnt model
根據(jù)7個主導環(huán)境變量與西藏飛蝗的分布數(shù)據(jù)構(gòu)建的最大熵模型模擬結(jié)果,按照高適生區(qū)、中適生區(qū)、低適生區(qū)和不適生區(qū)4個等級標準劃分,最終獲得西藏飛蝗在中國的適生區(qū)劃圖(圖2)。預測結(jié)果顯示,西藏飛蝗在當代氣候條件下預測的適生區(qū)范圍為88.79°—113.57°E,21.52°—38.33°N。其中高適生區(qū)主要位于四川和西藏兩省區(qū),包括四川的甘孜州,西藏的昌都地區(qū)、林芝地區(qū)、山南地區(qū)和拉薩市。中適生區(qū)沿高適生區(qū)周圍分布,主要集中青藏高原東部地區(qū),包括四川西部、西藏東部、云南北部等地。
西藏飛蝗當代適生區(qū)總面積為170.55×104km2,占中國國土面積的17.77%。其中,高、中、低適生區(qū)面積分別為22.51×104km2、63.04×104km2和85.01×104km2,分別占總適生區(qū)面積的13.2%、36.96%和49.84%。四川和西藏高適生區(qū)面積最大,分別為10.19×104km2和11.05×104km2(表5)。
2.3 西藏飛蝗地理分布與環(huán)境變量之間的關系
2.3.1 刀切法檢驗結(jié)果
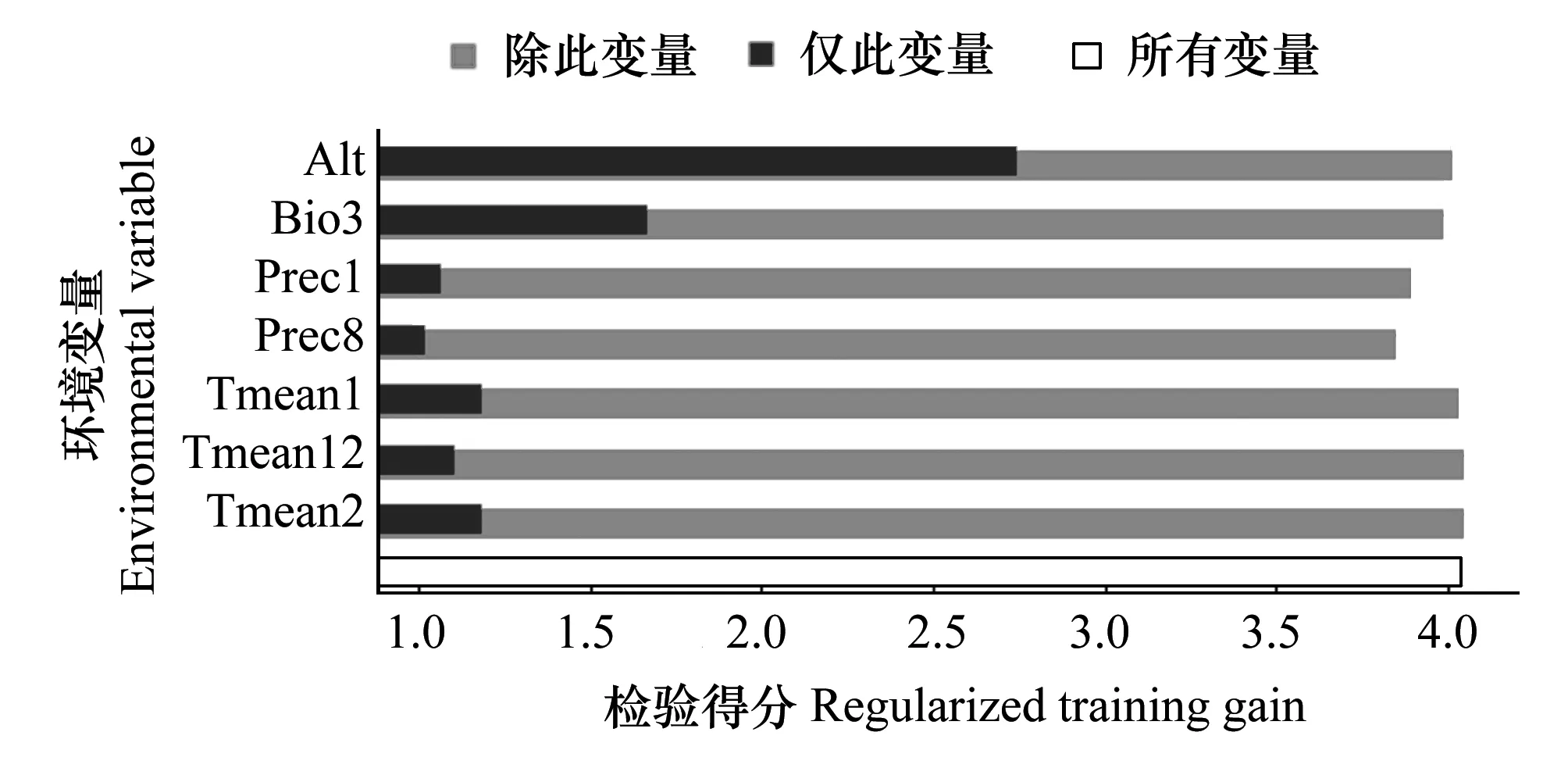
圖3 環(huán)境變量對西藏飛蝗分布的重要性(jackknife) Fig.3 Importance of environmental variables to Locusta migratoria tibetensis by jackknife analysis
刀切法檢驗結(jié)果可反映不同環(huán)境變量對于分布增益的貢獻大小,該方法分別計算“僅此變量”、“除此變量”和“所有變量”模擬時的訓練得分情況,判定標準為:“僅此變量”時得分較高,說明該因子具有較高的預測能力,對物種分布貢獻較大;“除此變量”時模型訓練得分能力降低較多,說明該變量具有較多的獨特信息,對物種分布較為重要。圖3為使用MaxEnt軟件自帶的刀切法分析各環(huán)境變量對西藏飛蝗分布重要性的結(jié)果圖,由圖中可見海拔(Alt)是影響西藏飛蝗分布的關鍵環(huán)境變量,其訓練增益超過2.5;等溫性(Bio3)也是較為重要的環(huán)境變量,其單獨訓練得分超過1.5;另外5種主導變量的重要性排序為2月份平均溫度(Tmean2)>1月份平均溫度(Ttmean1)>12月份平均溫度(Tmean12)>1月份平均雨量(Prec1)>8月份平均雨量(Prec8),這5種環(huán)境變量的訓練得分也均超過1.0。
2.3.2 環(huán)境變量對MaxEnt模型預測的影響
圖4是MaxEnt模型繪制的主導環(huán)境變量與分布概率之間的反饋曲線,該圖可反映不同閾值下環(huán)境變量的取值范圍。參照1.4中的分級方法,本文以0.33為閾來劃分適合西藏飛蝗分布的環(huán)境變量的范圍。結(jié)果顯示,海拔的適宜范圍為2870—5188 m,最適高度為3941 m,海拔2870—3941 m時,分布概率隨海拔升高而增加,3941—5188 m時,隨海拔升高而降低。等溫性的適宜值為38.5—47.2,最適値為42.9,在38.5—42.9,隨等溫性上升分布概率增大,在42.9—47.2時,隨等溫性升高分布概率降低。1、2、12月份平均溫度和8月份平均雨量與分布概率之間的反饋曲線的變化趨勢與上述2種變量類似,只是在變化幅度及取值范圍存在差異,具體見表6。在適宜范圍內(nèi),各主導環(huán)境變量的變化對西藏飛蝗的存在概率均有一定的影響,而取值在適宜范圍之外時對該蟲分布影響的概率逐漸減小。

表5 西藏飛蝗在當代氣候條件下的適生區(qū)面積預測
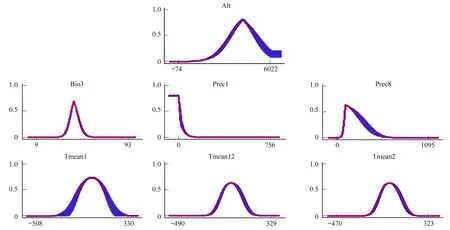
圖4 MaxEnt模型中研究物種對各環(huán)境變量的反饋曲線Fig.4 Response of curves of environmental variables in MaxEnt models
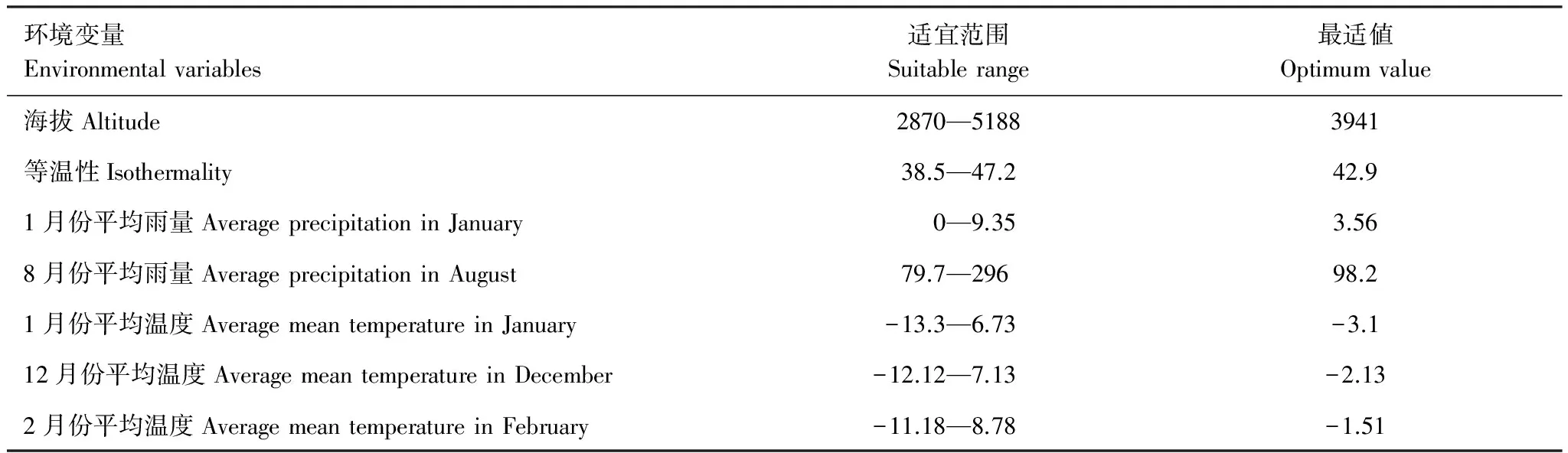
環(huán)境變量Environmentalvariables適宜范圍Suitablerange最適値Optimumvalue海拔Altitude2870—51883941等溫性Isothermality38.5—47.242.91月份平均雨量AverageprecipitationinJanuary0—9.353.568月份平均雨量AverageprecipitationinAugust79.7—29698.21月份平均溫度AveragemeantemperatureinJanuary-13.3—6.73-3.112月份平均溫度AveragemeantemperatureinDecember-12.12—7.13-2.132月份平均溫度AveragemeantemperatureinFebruary-11.18—8.78-1.51
3 討論
研究表明[45],環(huán)境變量的選擇會對生態(tài)位模型的預測結(jié)果造成一定的影響。應用MaxEnt模型預測物種地理分布的很多研究,采用世界氣候數(shù)據(jù)庫(http://www.worldclim.org)的Bioclim數(shù)據(jù)集中的19個生物氣候因子作為全部或主要環(huán)境變量。19個環(huán)境變量是以溫度和雨量為基礎數(shù)據(jù),根據(jù)不同需求計算演生而來,因此這些變量之間存在不可避免的自相關及多重線性重復等問題[46- 47]。研究業(yè)已證實,這些高度相關的變量在模型預測過程中會引入冗余信息,影響預測結(jié)果。在建模過程中,為避免上述問題,應首先對環(huán)境變量進行相關分析和有效篩選。本研究中,參考孫敬松等[48- 50]方法,以各因子對物種分布的貢獻率來評價變量的重要性,有效剔除了貢獻率較小的環(huán)境變量并對篩選的變量進行了相關性分析,獲得了7種主導環(huán)境變量并對模型進行重建,減少了冗余信息對模擬結(jié)果的影響,提高了預測結(jié)果的準確性。
目前對模型精度評價應用最廣泛的方法為ROC曲線法(即AUC法)[51],由于AUC不受診斷閾值的影響,并可提供所有閾值范圍上的性能評價結(jié)果,因此目前被公認為生態(tài)位模型評價領域的最佳評價指標。AUC的取值范圍在0.5—1,值越接近于1模型精度越高。本研究中基于主導環(huán)境變量的模擬的訓練集AUC平均值為0.993(非常接近1),預測結(jié)果達到“極好”水平,說明此次模型預測的地理分布與西藏飛蝗實際分布擬合度較高。另外,本研究利用地理信息系統(tǒng)軟件ArcGIS對MaxEnt輸出的柵格文件進行后期處理,使目標物種的分布數(shù)據(jù)與環(huán)境變量數(shù)據(jù)在柵格單元上相對應,有效的降低了系統(tǒng)誤差,進一步提高了數(shù)據(jù)的準確性。
本研究預測結(jié)果顯示,西藏飛蝗在中國的適生范圍為88.79°—113.57°E,21.52°—38.33°N,總面積達170.55×104km2。其中高適生區(qū)主要位于四川和西藏兩省區(qū),中適生區(qū)則主要集中青藏高原東部地區(qū),包括四川西部、西藏東部、云南北部等地。上述區(qū)域地理上包含了四川的多數(shù)草原(若爾蓋草原、紅原草原、塔公草原、龍燈草原和毛埡草原等)以及西藏雅魯藏布江及雅礱江河谷。李慶等[3]研究發(fā)現(xiàn),西藏飛蝗在草地環(huán)境下產(chǎn)卵塊數(shù)最多、草地環(huán)境更適合其生活,西藏飛蝗可對草地造成嚴重危害。因此西藏飛蝗在上述地區(qū)的危險系數(shù)極高,有關部門應引起高度重視,嚴防西藏飛蝗聚集成災。封傳紅等[52]建立了有效積溫與地理位置的模型,獲得了西藏飛蝗的潛在分布面積,結(jié)果表明,1998年西藏飛蝗分布面積最大,為14.3×104km2。本文中高適生區(qū)面積為22.51×104km2,中適生區(qū)面積則達63.04×104km2,與封傳紅的研究結(jié)果相差較大,這可能是因為使用的模型及變量不同所導致。
物種-環(huán)境關系是研究物種生態(tài)學需求和空間分布的重要方面,本研究分析了西藏飛蝗存在概率與主導環(huán)境變量之間的關系,獲得了相關的反饋曲線。結(jié)果表明,西藏飛蝗的存在概率隨著主導環(huán)境變量(海拔、等溫性、1月份及8月份平均雨量、1月份2月份及12月份平均溫度)的變化而改變。7個主導變量中,海拔和等溫性是影響該蟲分布最為重要的環(huán)境變量,表明西藏飛蝗擴散和繁殖過程中,一方面受當?shù)睾0畏秶闹萍s,另一方面也受當?shù)氐葴匦缘挠绊憽1狙芯恐形鞑仫w蝗適宜的海拔范圍為2870—5188m,最適為3941 m。目前已知西藏飛蝗的分布范圍僅限于青藏高原地區(qū),王思忠等[53]雖曾在四川雅安(海拔560 m左右)室內(nèi)成功飼養(yǎng)繁殖,但死亡率高,且眾多研究證明至今未發(fā)現(xiàn)該蟲在低海拔地區(qū)分布,這與本實驗的結(jié)果相吻合。等溫性反映了溫度變化的遲早及幅度,如當年氣溫轉(zhuǎn)變遲緩,則利于西藏飛蝗的胚胎發(fā)育,西藏飛蝗存在概率較高的等溫性指標在42.9左右,這表明該蟲在溫差較大情況下存在概率較高,這與上述的研究結(jié)果相近。李慶等研究了西藏飛蝗各發(fā)育階段的耐寒性,結(jié)果表明卵期的過冷卻點為-22.02℃,4齡蝗蝻的過冷卻點為-6.46℃,西藏飛蝗以卵越冬。西藏飛蝗蝗卵越冬期長達200 d,冬季低溫會影響其胚胎發(fā)育及孵化率。封傳紅等[54]研究發(fā)現(xiàn),溫度為限制西藏飛蝗分布的主要限制因子,夏季高溫不是限制因素,冬季低溫可影響西藏飛蝗的安全越冬,這些都說明冬季低溫對蝗卵的越冬至關重要,本研究認為西藏飛蝗的存在概率與越冬期12月及次年1、2月平均溫度有關,在較低的溫度條件下,該蟲仍有較高的存在概率,但過低的溫度將導致其存在概率下降;8月份平均雨量也是影響西藏飛蝗分布的重要因素,最適值為98.2 mm。研究發(fā)現(xiàn),西藏飛蝗與飛蝗其他亞種類似,主要棲居在較低濕的禾本科雜草地帶,該蟲一般在8月開始交配、產(chǎn)卵,適宜的濕度有利于該蟲順利的完成這些生命活動。此處需要注意,物種存在概率對環(huán)境變量的反饋曲線反映的是單一環(huán)境變量的作用,但昆蟲的生命活動受到各種環(huán)境變量(包括氣候因子、寄主條件、天敵種類、植被條件等)的綜合影響,因此此結(jié)果可作為判斷西藏飛蝗與環(huán)境變量之間關系的參考,但不能完全準確解釋二者之間的關系。
盡管MaxEnt模型預測具有操作簡便、樣本需求量小和預測精度高等優(yōu)勢,但同其他生態(tài)位預測模型類似不可避免的存在一定的局限性。(1)用MaxEnt模型進行預測時所涉及的環(huán)境變量除海拔外,其余變量均為氣候變量,19個生物氣候變量為西藏飛蝗實際分布地的氣候極值,即最大值和最小值,因而,MaxEnt模型所顯示的是物種分布的最大可能性,無法準備表達物種在現(xiàn)實分布的主要地區(qū)。上述預測結(jié)果更側(cè)重于理解和展現(xiàn)西藏飛蝗潛在地理分布以及揭示適于該物種分布的氣候特征。(2)基礎生態(tài)位是一種理想生態(tài)位,指某一物種在理想的生存條件而不存在其他物種競爭時所占據(jù)的最大生態(tài)位,該理論只需考慮非生物因素的影響[55]。MaxEnt模型對適生區(qū)的預測就是基于物種對基礎生態(tài)位的需求,但在實際的生存環(huán)境中,生物因子(如物種之間的相互作用、植被類型、地貌特征、物種自身擴散能力和土壤類型等)同樣會對預測物種的潛在分布產(chǎn)生重要影響[56]。例如,李慶等[3]研究了植被及土壤對西藏飛蝗產(chǎn)卵及分布的影響,結(jié)果表明,植被類型、土壤含鹽量、含水量和PH值能顯著影響西藏飛蝗的產(chǎn)卵及分布。基于上述原因,可以推測,MaxEnt模型預測的生態(tài)位比西藏飛蝗所占據(jù)的實際生態(tài)位要寬。對此,在下一步工作中,除考慮氣候因素影響外,還應注重考量物種間相互作用等生物因子的可信表達,以改善模型的預測效果。(3)過去30年來,隨著溫室氣體的不斷排放和保持,全球氣候正異常變暖,而氣候變化會引起物種生長及分布格局的變化[57- 59]。本研究中所使用的環(huán)境變量來源于世界氣候數(shù)據(jù)庫worldclim,時間跨度為1950—2000s,缺失了近期10多年的氣候數(shù)據(jù)。因此,在以后的工作中,應充實缺失的數(shù)據(jù),使預測結(jié)果更為準備可靠。
[1] 陳永林. 飛蝗新亞種—西藏飛蝗Locustamigratoriatibetensissubsp. n. 昆蟲學報, 1963, 12(4): 463- 475.
[2] 朱昱翰, 李慶, 楊剛, 白馬吞珠, 匡健康, 蔣春先, 王海建, 楊群芳. 低溫和光周期對西藏飛蝗體內(nèi)物質(zhì)的影響. 應用生態(tài)學報, 2016, 27(2): 629- 633.
[3] 李慶, 廖志昌, 楊剛, 封傳紅, 楊群芳, 羅懷海, 蔣春先, 王海建. 植被及土壤對西藏飛蝗產(chǎn)卵的影響. 中國農(nóng)業(yè)科學, 2012, 45(4): 656- 665.
[4] 陳永林. 蝗蟲和蝗災. 生物學通報, 1991, 26(11): 9- 12.
[5] 李慶, 吳蕾, 楊剛, 匡健康, 封傳紅, 羅懷海, 楊群芳, 蔣春先, 王海建. 溫度和紫外輻射脅迫對西藏飛蝗抗氧化系統(tǒng)的影響. 生態(tài)學報, 2012, 32(10): 3189- 3197.
[6] 王翠玲, 姚小波, 覃榮, 席永士, 王保海, 王文峰, 扎羅, 王考昌. 西藏飛蝗的發(fā)生規(guī)律與綜合防治技術(shù)探討. 西藏農(nóng)業(yè)科技, 2008, 30(4): 34- 40.
[7] 許仲林, 彭煥華, 彭守璋. 物種分布模型的發(fā)展及評價方法. 生態(tài)學報, 2015, 35(2): 557- 567.
[8] 朱耿平, 劉國卿, 卜文俊, 高玉葆. 生態(tài)位模型的基本原理及其在生物多樣性保護中的應用. 生物多樣性, 2013, 21(1): 90- 98.
[9] 張琳娜, 樊雋軒, Melchin M J, 陳清, Wu S Y, Goldman D, Mitchell C M, Sheets H D. 物種分布模型在古生物學研究中的應用. 古生物學報, 2013, 52(2): 146- 160.
[10] 吳慶明, 王磊, 朱瑞萍, 楊宇博, 金洪陽, 鄒紅菲. 基于MAXENT模型的丹頂鶴營巢生境適宜性分析——以扎龍保護區(qū)為例. 生態(tài)學報, 2016, 36(12): 3758- 3764.
[11] Anderson R P, Raza A. The effect of the extent of the study region on GIS models of species geographic distributions and estimates of niche evolution: preliminary tests with montane rodents (genusNephelomys) in Venezuela. Journal of Biogeography, 2010, 37(7): 1378- 1393.
[12] 張路. MAXENT最大熵模型在預測物種潛在分布范圍方面的應用. 生物學通報, 2015, 50(11): 9- 12.
[13] 李國慶, 劉長成, 劉玉國, 楊軍, 張新時, 郭柯. 物種分布模型理論研究進展. 生態(tài)學報, 2013, 33(16): 4827- 4835.
[14] Narouei-Khandan H A, Halbert S E, Worner S P, van Bruggen A H C. Global climate suitability of citrus huanglongbing and its vector, the Asian citrus psyllid, using two correlative species distribution modeling approaches, with emphasis on the USA. European Journal of Plant Pathology, 2016, 144(3): 655- 670.
[15] De Meyer M, Robertson M P, Mansell M W, Ekesi S, Tsuruta K, Mwaiko W, Vayssières J F, Peterson A T. Ecological niche and potential geographic distribution of the invasive fruit flyBactrocerainvadens(Diptera, Tephritidae). Bulletin of Entomological Research, 2010, 100(1): 35- 48.
[16] Hernández-Baz F, Romo H, González J M, de Jesús M, Hernández M, Gámez Pastrana R. Maximum entropy niche-based modeling (Maxent) of potential geographical distribution ofCoreuraalbicosta(Lepidoptera: Erebidae: Ctenuchina) in Mexico. Florida Entomologist, 2016, 99(3): 376- 380.
[17] Gill N S, Sangermano F. Africanized honeybee habitat suitability: a comparison between models for southern Utah and southern California. Applied Geography, 2016, 76: 14- 21.
[18] Bosso L, Di Febbraro M, Cristinzio G, Zoina A, Russo D. Shedding light on the effects of climate change on the potential distribution ofXylellafastidiosain the Mediterranean basin. Biological Invasions, 2016, 18(6): 1759- 1768.
[19] 齊國君, 高燕, 黃德超, 呂利華. 基于MAXENT的稻水象甲在中國的入侵擴散動態(tài)及適生性分析. 植物保護學報, 2012, 39(2): 129- 136.
[20] 楊會楓, 鄭江華, 吳秀蘭, 穆晨, 林俊, 許仲林. 基于MaxEnt模型的西伯利亞蝗蟲在新疆潛在分布預測研究. 新疆農(nóng)業(yè)科學, 2016, 53(1): 43- 50.
[21] 李宗亮, 于向鵬, 陸晨晨, 金海如. 基于物種預測分布模型對桔小實蠅在全球適生區(qū)的評估. 浙江師范大學學報: 自然科學版, 2011, 34(2): 207- 210.
[22] 馮益明, 劉洪霞. 基于Maxent與GIS的銹色棕櫚象在中國潛在的適生性分析. 華中農(nóng)業(yè)大學學報, 2010, 29(5): 552- 556.
[23] 董丹丹, 劉崇懷, 樊秀彩, 孫海生, 張國海, 王忠躍. 葡萄根瘤蚜在中國的風險性分析. 植物檢疫, 2011, 25(1): 21- 26.
[24] 趙文娟, 陳林, 丁克堅, 段霞瑜, 周益林. 利用MAXENT預測玉米霜霉病在中國的適生區(qū). 植物保護, 2009, 35(2): 32- 38.
[25] 曹學仁, 陳林, 周益林, 段霞瑜. 基于MaxEnt的麥瘟病在全球及中國的潛在分布區(qū)預測. 植物保護, 2011, 37(3): 80- 83.
[26] 韓陽陽, 王焱, 項楊, 葉建仁. 基于Maxent生態(tài)位模型的松材線蟲在中國的適生區(qū)預測分析. 南京林業(yè)大學學報: 自然科學版, 2015, 39(1): 6- 10.
[27] 曾輝, 黃冠勝, 林偉, 梁憶冰, 李志紅. 利用MaxEnt預測橡膠南美葉疫病菌在全球的潛在地理分布. 植物保護, 2008, 34(3): 88- 92.
[28] 楊群芳, 廖志昌, 李慶, 楊剛, 封傳紅, 蔣春先. 西藏飛蝗食性及其防治指標. 植物保護學報, 2008, 35(5): 399- 404.
[29] 葉永昌, 周廣勝, 殷曉潔. 1961- 2010年內(nèi)蒙古草原植被分布和生產(chǎn)力變化——基于MaxEnt模型和綜合模型的模擬分析. 生態(tài)學報, 2016, 36(15): 4718- 4728.
[30] 柳曉燕, 李俊生, 趙彩云, 全占軍, 趙相健, 宮璐. 基于MAXENT模型和ArcGIS預測豚草在中國的潛在適生區(qū). 植物保護學報, 2016, 43(6):1041- 1048
[31] Phillips S J, Dudík M, Schapire R E. A maximum entropy approach to species distribution modeling//Proceedings of the Twenty-First International Conference on Machine Learning. New York: ACM Press, 2004.
[32] 朱耿平, 劉晨, 李敏, 劉強. 基于Maxent和GARP模型的日本雙棘長蠹在中國的潛在地理分布分析. 昆蟲學報, 2014, 57(5): 581- 586.
[33] 趙晶晶, 高丹, 馮紀年. 基于MaxEnt模型的葡萄根瘤蚜在中國的適生性分析. 西北農(nóng)林科技大學學報: 自然科學版, 2015, 43(11): 99- 104, 112- 112.
[34] 王運生, 謝丙炎, 萬方浩, 肖啟明, 戴良英. ROC曲線分析在評價入侵物種分布模型中的應用. 生物多樣性, 2007, 15(4): 365- 372.
[35] 王雷宏, 楊俊仙, 徐小牛. 基于MaxEnt分析金錢松適生的生物氣候特征. 林業(yè)科學, 2015, 51(1): 127- 131.
[36] 趙力, 朱耿平, 李敏, 劉強. 入侵害蟲西部喙緣蝽和紅肩美姬緣蝽在中國的潛在分布. 天津師范大學學報: 自然科學版, 2015, 35(1): 75- 78.
[37] Elith J, Graham H C, Anderson R P, Dudík M, Ferrier S, Guisan A, Hijmans R J, Huettmann F, Leathwick J R, Lehmann A, Li J, Lohmann L G, Loiselle B A, Manion G, Moritz C, Nakamura M, Nakazawa Y, Overton J M, Peterson A T, Phillips S J, Richardson K, Scachetti-Pereira R, Schapire R E, Soberón J, Williams S, Wisz M S, Zimmermann N E, Araujo M. Novel methods improve prediction of species′ distributions from occurrence data. Ecography, 2006, 29(2): 129- 151.
[38] 應凌霄, 劉曄, 陳紹田, 沈澤昊. 氣候變化情景下基于最大熵模型的中國西南地區(qū)清香木潛在分布格局模擬. 生物多樣性, 2016, 24(4):453- 461.
[39] 段居琦, 周廣勝. 中國水稻潛在分布及其氣候特征. 生態(tài)學報, 2011, 31(22): 6659- 6668.
[40] 邢丁亮, 郝占慶. 最大熵原理及其在生態(tài)學研究中的應用. 生物多樣性, 2011, 19(3): 295- 302.
[41] 雷軍成, 徐海根, 吳軍, 關慶偉. 基于IPCC AR5的我國常綠闊葉林潛在適宜生境變化分析. 生態(tài)與農(nóng)村環(huán)境學報, 2015, 31(1): 69- 76.
[42] 殷曉潔, 周廣勝, 隋興華, 何奇瑾, 李榮平. 蒙古櫟地理分布的主導氣候因子及其閾值. 生態(tài)學報, 2013, 33(1): 103- 109.
[43] Lemke D, Hulme P E, Brown J A, Tadesse W. Distribution modelling of Japanese honeysuckle (Lonicerajaponica) invasion in the cumberland plateau and mountain region, USA. Forest Ecology and Management, 2011, 262(2): 139- 149.
[44] Yang X Q, Kushwaha S P S, Saran S, Xu J C, Roy P S. Maxent modeling for predicting the potential distribution of medicinal plant, Justicia adhatoda, L. in Lesser Himalayan foothills. Ecological Engineering, 2013, 51(1): 83- 87.
[45] 陳新美, 雷淵才, 張雄清, 賈宏炎. 樣本量對MaxEnt模型預測物種分布精度和穩(wěn)定性的影響. 林業(yè)科學, 2012, 48(1): 53- 59.
[46] 朱耿平, 劉強, 高玉葆. 提高生態(tài)位模型轉(zhuǎn)移能力來模擬入侵物種的潛在分布. 生物多樣性, 2014, 22(2): 223- 230.
[47] 王運生, 謝丙炎, 萬方浩, 肖啟明, 戴良英. 應用生態(tài)位模型研究外來入侵物種生態(tài)位漂移. 生態(tài)學報, 2008, 28(10): 4974- 4981.
[48] 孫敬松, 周廣勝. 利用最大熵法(MaxEnt)模擬中國冬小麥分布區(qū)的年代際動態(tài)變化. 中國農(nóng)業(yè)氣象, 2012, 33(4): 481- 487.
[49] 常志隆, 周益林, 趙遵田, 段霞瑜. 基于MaxEnt模型的小麥印度腥黑穗病在中國的適生性分析. 植物保護, 2010, 36(3): 110- 112, 129- 129.
[50] 何奇瑾, 周廣勝. 我國夏玉米潛在種植分布區(qū)的氣候適宜性研究. 地理學報, 2011, 66(11): 1443- 1450.
[51] 宋花玲. ROC曲線的評價研究及應用[D]. 上海: 第二軍醫(yī)大學, 2006.
[52] 封傳紅, 單緒南, 郭聰, 羅林明. 1961- 2005年西藏飛蝗潛在分布的變化. 昆蟲學報, 2011, 54(6): 694- 700.
[53] 王思忠. 西藏飛蝗生物學特性及耐寒性研究[D]. 雅安: 四川農(nóng)業(yè)大學, 2006.
[54] 封傳紅, 王思忠, 蔣凡, 楊剛, 游超, 李慶, 張敏, 羅林明. 溫度對四川省甘孜州西藏飛蝗分布的影響. 植物保護, 2008, 34(1): 67- 71.
[55] 蔡靜蕓, 張明明, 粟海軍, 張海波. 生態(tài)位模型在物種生境選擇中的應用研究. 經(jīng)濟動物學報, 2014, 18(1): 47- 52.
[56] 喬慧捷, 胡軍華, 黃繼紅. 生態(tài)位模型的理論基礎、發(fā)展方向與挑戰(zhàn). 中國科學: 生命科學, 2013, 43(11): 915- 927.
[57] 霍治國, 李茂松, 王麗, 溫泉沛, 肖晶晶, 黃大鵬, 王春艷. 氣候變暖對中國農(nóng)作物病蟲害的影響. 中國農(nóng)業(yè)科學, 2012, 45(10): 1926- 1934.
[58] 陳瑜, 馬春森. 氣候變暖對昆蟲影響研究進展. 生態(tài)學報, 2010, 30(8): 2159- 2172.
[59] 徐小鋒, 田漢勤, 萬師強. 氣候變暖對陸地生態(tài)系統(tǒng)碳循環(huán)的影響. 植物生態(tài)學報, 2007, 31(2): 175- 188.
猜你喜歡
課堂內(nèi)外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
英語世界(2023年10期)2023-11-17 09:18:18
中學生數(shù)理化·七年級數(shù)學人教版(2020年10期)2020-11-26 08:24:50
數(shù)學物理學報(2020年2期)2020-06-02 11:29:24
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
光學精密工程(2016年6期)2016-11-07 09:07:19
核科學與工程(2015年4期)2015-09-26 11:59:03
