太湖浮游植物初級生產力的生態模擬及其時空分布
2017-03-21 08:14:04宋玉芝王錦旗鄭建偉
江蘇農業科學 2016年11期
宋玉芝+王錦旗+鄭建偉
摘要:為了了解太湖浮游植物初級生產力的時空變化,利用3種常用模型[初級生產力(vertically generalized production model,簡稱VGPM)、Cadée模型、Talling模型]分別對江蘇太湖梅梁灣浮游植物日光合生產力進行估算,并用實測值驗證3種模型在該區域的適用性,從中選用模擬效果最佳的模型來估算2001—2011年太湖不同湖區的初級生產力,分析其時空變化規律。結果表明,VGPM模型、Cadée模型、Talling模型均適用于太湖梅梁灣浮游植物初級生產力的估算,VGPM模型的估算精度略高于其他2個模型,更接近真實值;VGPM模型估算結果表明,梅梁灣、湖心區及東太湖3個湖區的浮游植物初級生產力有明顯的季節性變化,以夏季最高,春秋季次之,冬季最低。從多年日均初級生產力的變化來看,3個湖區均表現為先提高后降低的趨勢;從空間上看,太湖浮游植物初級生產力呈現出明顯的空間異質性,太湖北部梅梁灣浮游植物初級生產力最高,湖心區次之,東太湖最低。
關鍵詞:浮游植物;初級生產力;生態模擬;太湖;梅梁灣
中圖分類號: Q178.1;S181.6 文獻標志碼: A
文章編號:1002-1302(2016)11-0419-04
隨著經濟的發展,進入水體氮、磷等營養鹽增多,湖泊富營養化程度日益加劇。而進入湖泊的氮、磷等營養鹽,通過浮游植物吸收同化作用進入食物鏈,在一定程度上決定了湖泊漁業產量[1]。事實上,湖泊富營養化的最終表現是水體浮游植物生長旺盛,初級生產力大幅度增加。因此,了解浮游植物初級生產力的變化規律,不僅有利于估算漁產潛力,而且可用于評價水體營養類型、指示湖泊環境特征[2-5]。目前,已有大量的關于湖泊等淡水水體初級生產特征和影響因素的報道。已有的研究表明,由于在不同區域的湖泊生態系統中,浮游植物初級生產力的大小既受生物量的影響,又與水環境中的光、營養鹽、溫度等多種因素密切相關[6-8]。以往用來估算浮游植物初級生產力的方法主要有黑白瓶測氧法、碳同位素示蹤法等,這些估算結果雖然準確,但費時費力。因此,近年來國內外學者相繼提出計算浮游植物初級生產力的一些經驗、半經驗半理論或者解析模式的計算模型[9-13]。其中Behrenfeld 等的初級生產力(vertically generalized production model,簡稱VGPM)、Cadée的簡化模型以及Talling的模型考慮水溫、光合有效輻射、湖泊葉綠素濃度和真光層深度等因素,能較準確地模擬水柱初級生產力,應用最為廣泛[12-14]。
太湖是我國第3大淡水湖泊,也是富營養化較為嚴重的湖泊[15-18]。近幾十年來,越來越多的學者運用模型來模擬湖泊的浮游植物初級生產力[12,19],然而關于太湖浮游植物初級生產力的時空變化還有待于進一步的研究。因此,本研究利用VGPM模型、Cadée模型及Talling模型3種常用的估算浮游植物初級生產力的模型,分別對太湖梅梁灣浮游植物日光合生產力進行估算,從中選用模擬效果最佳的模型來估算2001—2011年太湖不同湖區的初級生產力,分析其變化規律,為深入了解太湖生態系統提供科學依據。
1 數據來源及研究方法
1.1 研究區域概況
太湖是一個大型淺水湖泊,湖灣、沿岸及湖心等區域受地形影響,湖流結構及水土界面水力要素均有顯著差異。本研究選取太湖3個不同生態特征的湖區:藍藻水華頻繁暴發富營養化嚴重的梅梁灣(1#)、受外源影響較小但水動力擾動頻繁的湖心區(2#)、水生植物覆蓋度高且水質好的東太湖(3#)(圖1),以此估算不同類型湖區的初級生產力。
1.2 數據資料
2001—2011年氣象資料來自蘇州市東山氣象站;研究湖區各站點的水質資料主要來自《湖泊濕地海灣生態系統卷(江蘇太湖站)》[20]。
1.3 模型簡介
1.3.1 VGPM模型 Behrefeld 等收集了1971—1994年一類、二類水體共1 698個站點的11 283個實測數據,將葉綠素含量、真光層深度、光照周期標準化后發現,水體初級生產力垂直分布均呈相同形式,在此基礎上建立了初級生產力深度垂向歸納模型[9],最初用于海洋,但是由于該模型是在長時間、大范圍、不同水域上千萬個站點和實測數據的基礎上建立起來的,所以同樣適用于內陸水域[9],其表達式如下:
1.4 數據處理方法
將所獲的數據分類整理,用Excel 2003、SPSS 16.0軟件進行統計分析。
2 結果與分析
2.1 3種模型對梅梁灣浮游植物初級生產力的模擬及其適用性分析
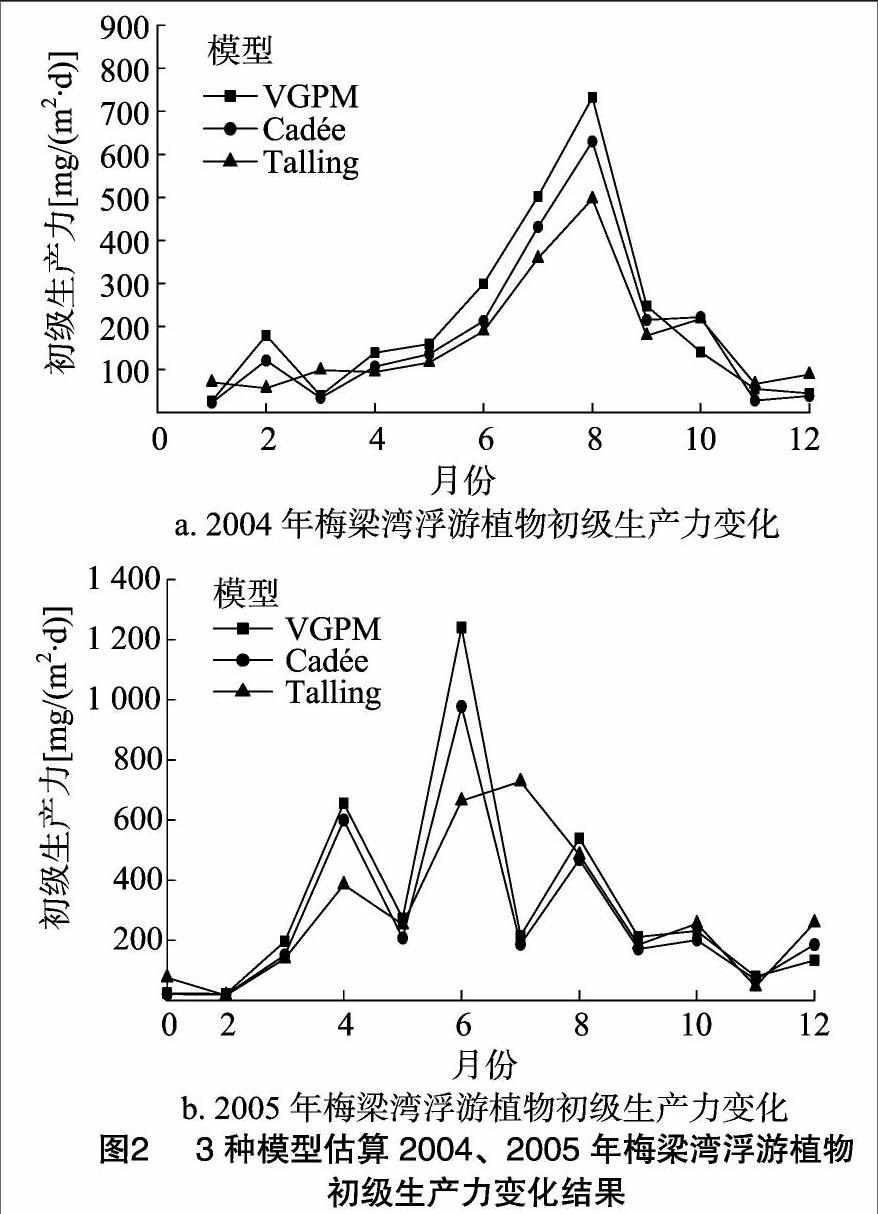
根據太湖梅梁灣2004—2005年的水質數據及東山氣象觀測站的氣象數據,用VGPM、Cadée、Talling模型對太湖梅梁灣浮游植物初級生產力進行模擬,將模擬結果與太湖梅梁灣浮游植物初級生產實力的實測值進行對比。
由圖2可以看出,3種模型所模擬的梅梁灣2004、2005年浮游植物初級生產力隨時間變化的趨勢基本相同,即隨時間的變化呈先增加后下降的趨勢。在不同年份,浮游植物初級生產力的變化較大。2004年浮游植物初級生產力最大值出現在8月份,最小值出現在1—2月份。2005年,用VGPM、Cadée 模型模擬的浮游植物初級生產力最大值出現在6月份,Talling模型所獲得浮游植物初級生產力最大值出現在7月份,3種模型模擬的最低值均出現2月份(圖2-b)。
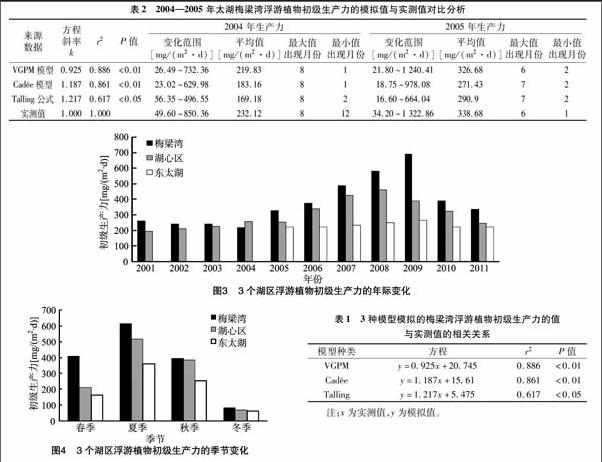
對比分析發現,3種模型模擬的梅梁灣浮游植物初級生產力與實測值均呈顯著或極顯著的線性相關(表1),這說明3種模型均可用來預測太湖梅梁灣浮游植物初級生產力。相比較而言,VGPM模型的模擬值與實測值相關性更好。進一步分析模擬值和實測值發現,VGPM模型的模擬值大于Cadée模型的估算值,Cadée模型的估算值大于Talling模型的估算值,但總體都小于實測值。由VGPM模型模擬的浮游植物初級生產力與實測值比較接近,而Cadée、Talling模型所獲得的浮游植物初級生力的模擬值相對來說略差(表2)。
2.2 太湖浮游植物初級生產力的時空分布特性
浮游植物初級生產力受光照周期、真光層深度、葉綠素濃度等因素的影響,而這些因素又受水文、水質、氣象等因素的綜合作用,因此浮游植物初級生產力是一個不斷變化的復雜過程。太湖水域面積約2 338km2,不同區域水質狀況、水生動植物群落組成差異很大,因此用VGPM模型對梅梁灣、東太湖、湖心區2001—2011年浮游植物的初級生產力進行估算,以了解太湖浮游植物初級生產力的時空分布規律。
2.2.1 太湖不同湖區浮游植物初級生產力年際變化及季節變化 從圖3可見,2001—2004年梅梁灣與湖心區的浮游植物年平均初級生產力較為穩定,變化不大,其中東太湖水質資料缺失;從2004年開始,梅梁灣與湖心區的初級生產力先逐年增加后逐年下降,而東太湖的浮游植物初級生產力沒有明顯的年際變化。
從季節變化來看,3個湖區初級生產力有明顯的季節變化,夏季最高,冬季最低,春秋季次之;還可以看出,梅梁灣的初級生產力在春季及秋季相當,而湖心區與東太湖秋季初級生產力卻高于春季(圖4)。
2.2.2 太湖浮游植物初級生產力空間變化 進一步比較梅梁灣、湖心區、東太湖3個湖區浮游植物初級生產力的狀況可知,2001—2004年,梅梁灣與湖心區的初級生產力相差不大,而2005—2011年間,3個湖區浮游植物初級生產力間具有明顯差異,梅梁灣的初級生產力最大,湖心區次之,東太湖最小(圖3、圖4)。從季節上看,在冬季,梅梁灣初級生產力雖高于湖心區及東太湖,但3個湖區浮游植物初級生產力差異不大;在夏季及春季,梅梁灣初級生產力明顯高于湖心區,湖心區高于東太湖;在秋季,梅梁灣與湖心區相差不大,而東太湖卻明顯低于梅梁灣及湖心區(圖4)。
3 討論
VGPM、Cadée及Talling模型均考慮了水溫、光照等因子對水柱初級生產力的影響,在實際運用中比較廣泛。通過3種模型對梅梁灣初級生產力模擬結果與實測值的對比分析發現,3種模型的模擬值與實測值均呈較高的線性相關關系,都可以反映浮游植物初級生產力的變化情況,但是VGPM模型估算的浮游植物初級生產力最接近實測值,Cadée、Talling模型略差。曾臺衡等利用模型對長江中下游湖區浮游植物初級生產力進行估算時也得出相似的結果[13]。這可能是Cadée模型實際考慮的要素有所精簡,而太湖水環境比較復雜,可能是導致Cadée模型估算值略差的原因之一;而Talling公式中缺少真光層深度這一重要因子,并且式中垂直消光系數取的是經驗值[13-14],可能存在誤差,從而影響其模擬的準確性。
太湖梅梁灣、湖心區以及東太湖3個湖區的浮游植物初級生產力均存在明顯的季節性差異,夏季最高,春秋季次之,冬季最低。但不同湖區,秋季、春季浮游植物初級生產力狀況有所不同,湖心區及東太湖,秋季浮游植物初級生產力高于春季,而梅梁灣秋季與春季差別不大。這是由于浮游植物初級生產力的影響因素較為復雜,不同季節最直觀的差異即溫度不同,夏季、秋季氣溫較高,是浮游植物活動及生長繁殖較為旺盛的季節,初級生產力也達全年的最高值;而冬季藍藻等喜溫浮游植物還處于休眠狀態,初級生產力比較低,對于春季浮游植物處于復蘇的階段,生產力也相對較低。然而,梅梁灣春季與秋季初級生產力相當,一方面可能是梅梁灣藻種源基數比較高,另一方面也可能是夏季藻類發生水華后,這部分藻類大量死亡,水質急劇下降,也會影響秋季浮游植物的初級生產力[21]。再加上不同水域浮游植物群落結構不同,優勢種不同,它們對環境因子的響應程度也不相同,從而影響到湖泊水體的初級生產力。從3個湖區的浮游植物年平均初級生產力來看,梅梁灣和湖心區在2001—2011年總體表現為先增長后下降的趨勢,在2007—2009年達到最高值,隨后逐步降低。這可能是由于人為對太湖周邊進行治理整頓,減少氮磷等污染物的輸入 [15,21],使得水體葉綠素a含量發生變化,進而影響浮游植物的初級生產力。
太湖浮游植物初級生產力存在空間變化規律,總的表現為北部高,湖心次之,南北低。具體而言,梅梁灣浮游植物初級生產力明顯高于其他湖區,湖心區次之,東太湖的浮游植物初級生產力相對而言最低。這是由于梅梁灣位于太湖北端,氮磷濃度高,水體葉綠素濃度高[19],當氣象、水文、水質達到最佳條件時浮游植物便開始迅速生長繁殖致使水體初級生產力不斷增長。夏季開始出現藍藻水華現象,草型湖區已向藻型湖區轉變,浮游植物初級生產力也隨即增長;東太湖屬草型湖區,沉水植物占優勢,真光層深度相對較高,風浪等作用相對較弱,浮游植物種類和數量較少[12],其初級生產力與其他湖區相比也稍低些;而在湖心區,營養鹽濃度相對較高,由于受風浪的擾動,水下光衰減迅速,進而影響浮游植物初級生產力[12,20-21]。總之,浮游植物初級生產力的變化是一個復雜的過程,不僅受浮游植物群落結構、藻的種類和數量的影響,同時受氣溫、氮的供給、浮游動物選擇性牧食等多種因素的影響。
4 結論
(1)VGPM、Cadée、Talling模型均可以反映太湖梅梁灣浮游植物初級生產力的變化情況,但VGPM模型的估算精度略高于其他2個模型,更接近實測值。
(2)太湖梅梁灣、湖心區及東太湖浮游植物初級生產力存在明顯的季節性差異,夏季最高,冬季最低,春秋季次之;年平均初級生產力在2001—2011年總體表現為先增長后下降的趨勢。
(3)太湖北部梅梁灣浮游植物初級生產力最高,湖心區次之,太湖南部東太湖浮游植物初級生產力最低。
參考文獻:
[1]王 驥,梁彥齡.用浮游植物的生產量估算武昌東湖鰱鳙生產潛力與魚種放養量的探討[J]. 水產學報,1981,5(4):343-350.
[2]韓德舉,吳生桂,鄒 清,等. 陸水水庫的浮游生物及營養類型[J]. 湖泊科學,1996,8(4):351-358.
[3]王信海,姜愛蘭,丁辰龍,等. 龍虎泡秋季浮游生物組成與水質理化特征分析[J]. 江蘇農業科學,2009(3):363-366.
[4]朱愛民,劉家壽,胡傳林,等. 湖北浮橋河水庫浮游植物初級生產力及其管理[J]. 湖泊科學,2007,19(3):340-344.
[5]蘇潔瓊,王烜,楊志峰. 考慮氣候因子變化的湖泊富營養化模型研究進展[J]. 應用生態學報,2012,23(11):3197-3206.
[6]Chen Y W,Qin B Q,Teubner K,et al. Long-term dynamics of phytoplankton assemblages:microcystis-domination in Lake Taihu,a large shallow lake in China[J]. Journal of Plankton Research,2003,25(4):445-453.
[7]Vaillancourt R D,Sambrotto R N,Green S,et al. Phytoplankton biomass and photosynthetic competency in the summer time Mertz Glacier Region of East Antarctica[J]. Deep Sea Research PartⅡ:Topical Studies in Oceanography,2003,50(8/9):1415-1440.
[8]汪益嬪,張維硯,徐春燕,等. 淀山湖浮游植物初級生產力及其影響因子[J]. 環境科學,2011,32(5):1249-1256.
[9]Behrenfeld M J,Falkowski P G. Photosynthetic rates derived from satellite-based chlorophyll concentration[J]. Limnology and Oceanography,1997,42(1):1-20.
[10]Cadée G C. Primary production off the Guyana coast [J]. Netherlands Journal of Sea Research,1975,9(1):128-143.[HJ1.67mm]
[11]Talling J F. The phytoplankton population as a compound photosynthetic system[J]. New Phytologist,1957,56(2):133-149.
[12]張運林,馮 勝,馬榮華,等. 太湖秋季真光層深度空間分布及浮游植物初級生產力的估算[J]. 湖泊科學,2008,20(3):380-388.
[13]曾臺衡,劉國祥,胡征宇. 長江中下游湖區浮游植物初級生產力估算[J]. 長江流域資源與環境,2011,20(6):717-722.
[14]易齊濤,徐 鑫,曲喜杰,等. 兩淮采煤沉陷積水區水體初級生產特征[J]. 中國環境科學,2014,34(8):2101-2110.
[15]朱廣偉. 太湖富營養化現狀及原因分析[J]. 湖泊科學,2008,20(1):21-26.
[16]Song Y Z,Wang J Q,Gao Y X,et al. The physiological responses of Vallisneria natans,to epiphytic algae with the increase of N and P concentrations in water bodies[J]. Environmental Science & Pollution Research International,2015,22(11):8480-8487.
[17]席運官,田 偉,李 妍,等. 太湖地區稻麥輪作系統氮、磷徑流排放規律及流失系數[J]. 江蘇農業學報,2014,30(3):534-540.
[18]亢志華,劉華周. 基于成本-收益分析法的太湖氮磷污染物去除生態補償[J]. 江蘇農業學報,2015,31(4):942-945.
[19]殷 燕,張運林,時志強,等. 基于VGPM模型和MODIS數據估算梅梁灣浮游植物初級生產力[J]. 生態學報,2012,32(11):3528-3537.
[20]秦伯強,胡春華. 湖泊濕地海灣生態系統卷:江蘇太湖站(1991—2006) [M]. 北京:中國農業出版社,2010.
[21]蔡琳琳,朱廣偉,李向陽. 太湖湖岸帶浮游植物初級生產力特征及影響因素[J]. 生態學報,2013,33(22):7250-7258.
