紫萍休眠體的萌發誘導研究
2017-04-05 18:28:15李蘭芝吳坤鑫張家明
江蘇農業科學 2016年12期
李蘭芝++吳坤鑫++張家明
摘要:以紫萍DW2501-4和HB0301產生的休眠體為材料,利用單因子試驗和正交試驗研究不同蔗糖含量、不同的GA3濃度、不同的溫度和光照條件誘導紫萍休眠體萌發的最適條件。結果表明,在1%~5%質量濃度蔗糖范圍內,高的蔗糖質量濃度會抑制休眠體的萌發,質量濃度為1%、2%的蔗糖對DW2501-4和 HB0301休眠體萌發誘導效果最好。在20~32 ℃,DW2501-4休眠體在32 ℃下萌發效果最佳,HB0301休眠體在28 ℃下萌發效果最佳。在0~10 mg/L 范圍內GA3能促進DW2501-4和HB0301休眠體的萌發,最佳濃度為0.1 mg/L。DW2501-4和HB0301在光—暗周期為24 h—0 h和16 h—8 h下萌發勢最高,葉狀體數量最多。3種日照時間下萌發率差異不顯著,但長日照可縮短休眠體萌發時間,促進葉狀體繁殖。DW2501-4和HB0301-4休眠體的最佳萌發誘導條件是0.5倍 Hoaglands 培養基中添加1%蔗糖和0.1 mg/L的GA3,在28 ℃,光—暗周期為16 h—8 h的條件下培養。
關鍵詞:紫萍;休眠體;萌發;最佳條件
中圖分類號: Q945.1;S555+.901文獻標志碼: A
文章編號:1002-1302(2016)12-0532-05
收稿日期:2016-05-19
基金項目:國家國際科技合作專項(編號:2014DFA30680)。
作者簡介:李蘭芝(1990—),女,湖北廣水人,碩士研究生,研究方向為植物分子遺傳學,E-mail:317004873@qq.com;共同第一作者:吳坤鑫(1969—),男,福建永定人,博士,副研,研究方向作物遺傳育種,E-mail:wukunxin@itbb.org.cn。
通信作者:張家明,博士,研究員。研究方向為生物質能源。E-mail:jmzhang@vip.163.com。
紫萍(Spirodela polyrrhiza (L.) Schleid.)屬于浮萍科(Lemnaceae)紫萍屬(Spirodela)。自然界中,夏末生長季節結束時紫萍會產生休眠體(turion)[1]。紫萍產生的休眠體直徑大約為2 mm,呈橢圓形,灰褐色。上表面平滑,下表面微微凸出。新形成的休眠體會沉入水底進行冬眠,經歷一段時間的低溫冬眠后,春天隨著溫度上升,光照時間延長,細胞內碳水化合物的代謝發生變化,冬眠才會被打破,休眠體開始萌發[2-3]。休眠體萌發時會從2個分生側囊中長出新的營養生殖葉片[4]。通過消耗休眠體內積累的大量淀粉,新葉會快速生長[4]。寒冷處理對紫萍休眠體進行預催熟而更有利于休眠體的萌發,光照、充足的碳水化合物的供應是促進休眠體萌發的必要條件[5-7]。影響休眠體萌發的主要因子包括溫度、光照、營養條件、GA3等[7-10]。在實驗室內紫萍的種質資源也可利用休眠體進行保種,研究休眠體快速萌發的條件是實驗室紫萍保種研究的重要部分。本試驗通過研究不同蔗糖含量、不同的GA3濃度、不同的溫度和光照條件對紫萍休眠體萌發的影響,以期為在實驗室長期保存紫萍種質資源提供理論依據。
1材料和方法
1.1試驗材料
試驗材料選用本實驗室保存的采自海南的紫萍 DW2501-4和湖北的紫萍HB0301。試驗于2015年1—12月在中國熱帶農業科學院熱帶生物技術研究所試驗基地溫室內進行。利用其無菌無藻的葉狀體,在含0.005 mg/L ABA的0.5倍缺氮的Hoaglands培養基,24 ℃,16 h—8 h光—暗周期條件下,誘導休眠體。將誘導出的休眠體放在4 ℃冰箱中黑暗處理7 d,再接種到固體培養基中進行萌發誘導。本試驗用含瓊脂8 g/L的0.5倍Hoaglands固體培養基,培養基pH值為5.8~6.0。
1.2試驗方法
1.2.1單因子誘導試驗
不同蔗糖含量誘導試驗中,0.5倍Hoaglands 固體培養基作為對照組,用添加1%、2%、3%、5%(質量濃度)蔗糖的0.5倍Hoaglands固體培養基進行萌發誘導,培養條件為28 ℃,24 h光照。溫度試驗中,溫度設置為20、24、28、32 ℃,添加1%蔗糖的0.5倍Hoaglands固體培養基中,24 h光照進行萌發誘導。光照試驗中,光—暗周期為8 h—16 h、16 h—8 h、24 h—0 h,添加1%蔗糖的0.5倍Hoaglands固體培養基中,28 ℃下進行萌發誘導。GA3誘導試驗中,在添加不同濃度的GA3(濃度設置為0.001、0.010、0.100、1.000、10.000 mg/L)的含1%蔗糖的0.5倍Hoaglands固體培養基中進行誘導,培養條件為28 ℃,24 h光照。每個培養皿中接種20個休眠體,重復3次,置于光溫培養箱中培養。
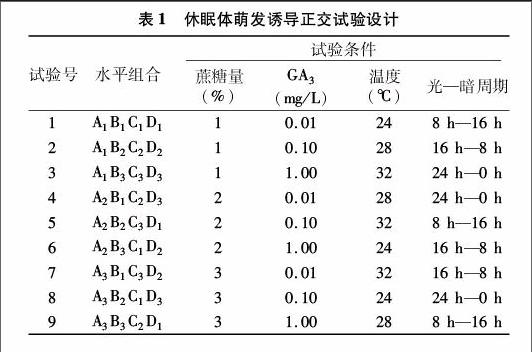
1.2.2正交試驗
進行4因素3水平的L9(34)正交試驗,各因素水平見表1。A因素是蔗糖質量濃度,設1%、2%、3% 3個水平,B因素是GA3,設0.01、0.10、1.00 mg/L 3個水平,C因素是溫度,設24、28、32 ℃ 3個水平,D因素是光—暗周期,設8 h—16 h、16 h—8 h、24 h—0 h 3個水平。
每個皿接種20個休眠體,重復3次。根長1 mm或者葉狀體直徑1 mm均視為萌發,每24 h觀察記錄休眠體萌發情況,記錄休眠體萌發情況和葉狀體生長狀態,得到休眠體萌發誘導最佳條件。
1.2.3萌發測定指標
萌發時滯,即萌發啟動時間,指從萌發誘導試驗開始到第1個休眠體開始萌發所需要的時間;萌發高峰:日萌發數達到最大時的天數;萌發持續時間,從休眠體開始萌發到最后1個休眠體萌發所需的總天數;萌發勢=日萌發數最大時的總數/休眠體總數×100%;萌發率=7 d時休眠體萌發總數/休眠體總數×100%;葉狀體數:7 d時葉狀體總數。
1.3數據統計與分析
采用Excel進行數據處理,用SPSS 19.0統計分析軟件進行數據的方差分析與差異顯著性檢驗。
2結果與分析
2.1蔗糖含量對紫萍休眠體萌發的影響
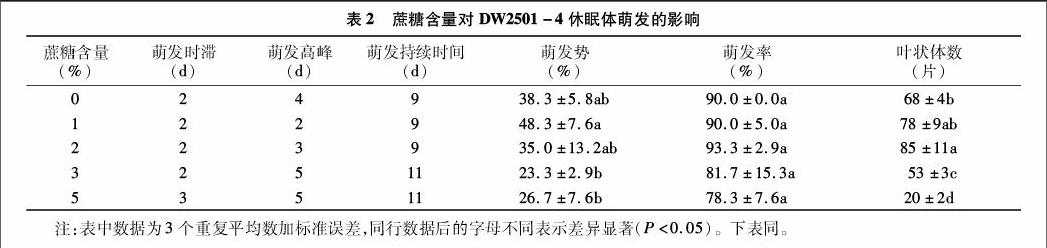
蔗糖含量對DW2501-4休眠體萌發的影響:如表2所示,在蔗糖含量為0、1%、2%、3%條件下,DW2501-4休眠體在第2 d開始萌發;在蔗糖含量為5%條件下,休眠體在第3 d開始萌發。在蔗糖含量為0條件下,休眠體在第4 d萌發達到高峰期,在蔗糖含量1%、2%、3%、5%條件下,依次在第 2 d、第3 d、第5 d、第5 d休眠體萌發達到高峰期;在蔗糖含量為0、1%、2%條件下,休眠體萌發持續9 d,3%、5%條件下,休眠體萌發持續11 d;在蔗糖含量為1%、2%條件下,葉狀體數顯著高于3%、5%。說明在蔗糖含量為1%、2%條件下,能有效誘導DW2501-4休眠體萌發,3%和5%的蔗糖抑制DW2501-4休眠體萌發。
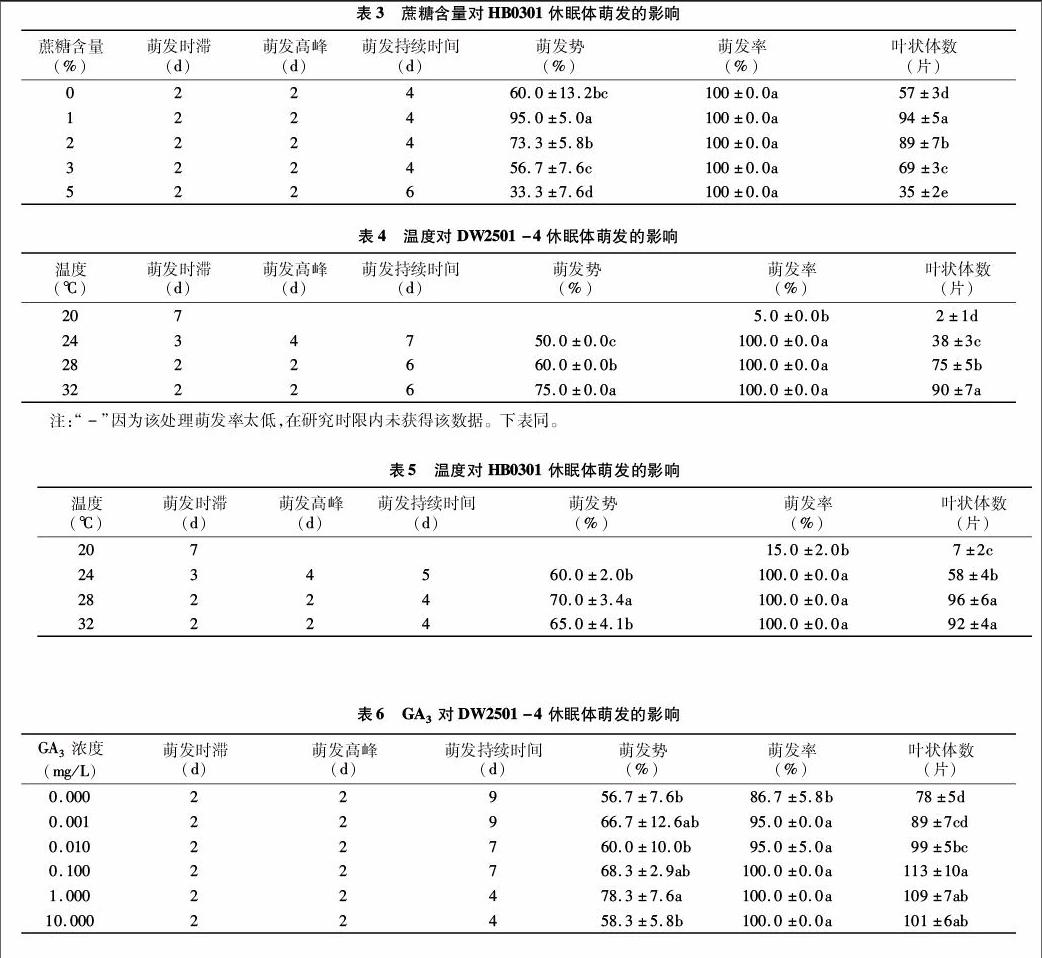
蔗糖含量對HB0301休眠體萌發的影響:如表3所示,在蔗糖含量為0、1%、2%、3%、5%條件下,HB0301休眠體均在在第2 d開始萌發并達到高峰期;在蔗糖含量為0、1%、2%、3%條件下,休眠體全部萌發持續4 d,5%條件下,休眠體萌發持續6 d;在蔗糖含量為1%條件下,萌發勢、葉狀體數均顯著高于其他組,在2%條件下葉狀體數顯著高于0、3%、5%條件下,而在5%蔗糖下萌發勢和葉狀體數則顯著低于不含蔗糖的。說明在蔗糖含量為1%、2%條件下,能有效誘導HB0301休眠體萌發;5%的蔗糖抑制HB0301休眠體萌發。
在1%~5%蔗糖含量范圍內,高的蔗糖含量會抑制休眠體的萌發;含量為1%、2%的蔗糖對DW2501-4和 HB0301休眠體萌發誘導效果最好;在2%~5%蔗糖下,HB0301休眠體萌發高峰期早于DW2501-4,且萌發率和萌發勢高于DW2501-4。
2.2溫度的對紫萍休眠體萌發的影響
溫度對DW2501-4休眠體萌發的影響:如表4所示,在20 ℃下,DW2501-4休眠體在7 d時開始萌發,在24 ℃下,休眠體在3 d時開始萌發,在28、32 ℃下,休眠體在2 d時開始萌發;在24、28、32 ℃下,休眠體依次在4、2、2 d達到休眠體[CM(25]萌發高峰期,在20[KG*3]℃下,萌發遲緩,誘導7[KG*3]d未出現高峰期;在24、28、32 ℃下,休眠體萌發持續時間依次為7、6、6 d;在32 ℃下萌發勢、葉狀體數均顯著高于其他組。由表5可見,在20 ℃下,HB0301休眠體在7 d時開始萌發,在24 ℃下,休眠體在3 d時開始萌發,在28、32 ℃下,休眠體在2 d時開始萌發;在24、28、32 ℃下,休眠體依次在4、2、2 d達到休眠體萌發高峰期,在20 ℃下,萌發遲緩,誘導7 d未出現高峰期;在24、28、32 ℃下,休眠體萌發持續依次為5、4、4 d;HB0301在28、32 ℃下葉狀體數均顯著高于20、24 ℃。
[FK(W+32mm][HT6H][WTHZ][JZ]表5溫度對HB0301休眠體萌發的影響[HTSS]
[HJ*5][BG(!][BHDFG3,WK6,WK7。3,WK13。2,WK7W]溫度(℃)萌發時滯(d)萌發高峰(d)萌發持續時間(d)萌發勢(%)萌發率(%)葉狀體數(片)
[BHDG1*2,WK6,WK7。3,WK13。2DW,WK7W]20715.0±2.0b7±2c
[BHDW]2434560.0±2.0b100.0±0.0a58±4b
[BH]2822470.0±3.4a100.0±0.0a96±6a
[BH]3222465.0±4.1b100.0±0.0a92±4a[BG)F]
在20~32 ℃,大體上隨著溫度升高,休眠體萌發加快,葉狀體繁殖增多;雖然24~32 ℃萌發率一樣,但是28、32 ℃的萌發時滯和萌發高峰都要比24 ℃快;DW2501-4休眠體在32 ℃下萌發效果最佳,HB0301休眠體在28 ℃下萌發效果最佳。在24、28 ℃下,HB0301比DW2501-4所需萌發持續時間短,萌發勢高。
2.3GA3濃度對紫萍休眠體萌發的影響
GA3濃度對DW2501-4休眠體萌發的影響見表6。在0.000~10.000 mg/L范圍內的幾個GA3試驗濃度下,DW2501-4休眠體均在2 d時開始萌發,并且在2 d時達到萌發高峰期;在GA3濃度為0.000 mg/L和0.001 mg/L時,休眠體萌發持續9 d,在GA3濃度為0.010 mg/L和0.100 mg/L時,休眠體萌發持續7 d,在GA3濃度為1.000 mg/L和 1000 mg/L 時,休眠體萌發持續4 d;在0.010~10.000 mg/L范圍內的幾個GA3試驗濃度下,萌發率和葉狀體數均顯著高于GA3濃度為0.000 mg/L時,說明這幾個GA3濃度均對休眠體萌發起誘導作用。其中在10.000 mg/L濃度下,雖然萌發率高但葉狀體分化幾天后玻璃化。在GA3濃度為 0.100 mg/L 和1.000 mg/L時DW2501-4休眠體萌發勢、萌發率高,葉狀體總數多,生長狀態最好。
GA3濃度對HB0301休眠體萌發的影響見表7。在0000~10.000 mg/L范圍內的幾個GA3試驗濃度下,HB0301休眠體均在2 d時開始萌發,并且在2 d時達到萌發高峰期;在GA3濃度為0.000、0.010、0.100、1.000 mg/L時,休眠體萌發持續3 d,在GA3濃度為10.000 mg/L時,休眠體萌發持續4 d,葉狀體生長受抑制且幾天后玻璃化,在GA3濃度為0.001 mg/L時,休眠體萌發持續5 d;不同GA3濃度下,萌發率無差異,但GA3濃度為0.010 mg/L和0.100 mg/L時萌發勢、 葉狀體數均顯著高于 1.000 mg/L 和 10.000 mg/L時[CM(25]。說明HB0301在GA3濃度為 0.100 mg/L 時萌發誘導效果最佳。
與不添加GA3相比在0.001~10.000 mg/L范圍內GA3能促進DW2501-4和HB0301休眠體的萌發,在GA3濃度為 0.100 mg/L 時休眠體萌發率高,葉狀體數最多,萌發效果最佳。
2.4光照時間對紫萍休眠體萌發的影響
由表8可見,在28 ℃,光—暗周期為24 h—0 h、16 h—8 h、8 h—16 h 下,DW2501-4休眠體在2 d時開始萌發,且在2 d時達到休眠體萌發高峰期;光—暗周期為24 h—0 h時和16 h—8 h時休眠體萌發持續時間都是6 d,小于8 h—16 h的9 d,而在16 h—8 h時萌發勢最高,在24 h—0 h時產生的葉狀體數最多。從表9可知,在28 ℃,光—暗周期為24 h—0 h、16 h—8 h、8 h—16 h 下,HB0301休眠體在2 d時開始萌發,且在2 d時達到休眠體萌發高峰期;在光—暗周期為 24 h—0 h和16 h—8 h下休眠體萌發持續時間都是4 d,小于8 h—16 h的5 d,而在 24 h—0 h 和16 h—8 h下萌發勢和葉狀體繁殖數量都要比 8 h—16 h 下高。總之,在上述3個光—暗周期下DW2501-4和HB0301中休眠體萌發率沒有顯著差異,但長日照可縮短休眠體萌發持續時間,并促進葉狀體繁殖。
2.5正交試驗結果分析
由表10可知,在1~6號試驗條件下,誘導7 d DW2501-4休眠體的萌發率均為100%,但在2號和3號試驗條件下,萌發完全所需時間最少,3號試驗條件下萌發勢最高,2號試驗條件下葉狀體最多。從表11可知,在1~7號試驗條件下,誘導7 d HB0301-4休眠體的萌發率均為100%,但在2號試驗條件下,萌發完全所需時間少,萌發勢最高,葉狀體最多。
DW2501-4和HB0301-4休眠體的最佳誘導條件是,05倍Hoaglands固體培養基中添加1%蔗糖和0.100 mg/L的GA3,在28 ℃,光—暗周期為16 h—8 h的條件下培養。
3結論與討論
可能會抑制休眠體的分化萌發。DW2501-4在蔗糖質量濃度為1%條件下萌發勢最高,葉狀體數在2%蔗糖下產生最多,其次是1%蔗糖。HB0301[JP+1]在蔗糖質量濃度為1%條件下萌發勢最高,葉狀體數也最多,其次是2%蔗糖。在3%蔗糖和5%蔗糖下,DW2501-4休眠體萌發完全所需時間相對較長,萌發勢和萌發率相對較低。隨著培養基營養消耗,蔗糖含量降低,對分化的抑制作用降低,剩余的蔗糖可以繼續為葉狀體的分化和生長供能。因此含2%蔗糖的培養基營養供給比含1%蔗糖的多,對于葉狀體的持續分化和生長繁殖更有利。
在0.001~1.000 mg/L范圍內,總體上隨著GA3濃度升高,DW2501-4休眠體分化萌發持續時間越短,在 10.000 mg/L 濃度下葉狀體數量較多,但葉狀體分化幾天后玻璃化。DW2501-4在GA3濃度為1.000 mg/L時休眠體萌發勢最高,但在0.100 mg/L濃度下葉狀體總數最多,生長狀態最好。HB0301在GA3濃度為0.100 mg/L時,萌發所需時間短,萌發勢和萌發率高、葉狀體數多。GA3在一定濃度范圍內對休眠體萌發有誘導作用,且有最佳誘導濃度,因此GA3是誘導休眠體萌發的有效因子。
在20~28 ℃,總體隨著溫度升高,DW2501-4 和HB0301休眠體萌發時滯逐步減少,葉狀體繁殖增多。休眠體的萌發率從20 ℃的低位快速提升到24 ℃的100.0%。20 ℃下休眠體也可以萌發,只是萌發遲緩,說明溫度條件對休眠體萌發起重要作用;32 ℃下,雖然休眠體的萌發率和葉狀體數量也多,但 28 ℃ 下葉狀體生長狀態更好。更低或者更高的溫度都不利于HB0301和DW2501-4休眠體的萌發和葉狀體生長。在24 ℃和28 ℃,HB0301休眠體比DW2501-4休眠體萌發持續時間短,萌發勢高,說明相同條件下HB0301休眠體比DW2501-4休眠體更易萌發。[JP+1]
DW2501-4 在光—暗周期為16 h—8 h下萌發勢最高,24 h—0 h [JP+1]時產生葉狀體數量最多;HB0301在光—暗周期為 24 h—0 h 下萌發勢最高,葉狀體數量最多;3個光—暗周期下萌發率沒有明顯差異,但長日照可縮短休眠體萌發時間。
參考文獻:
[1]Appenroth K J,Nickel G. Turion formation in Spirodela polyrhiza:The environmental signals that induce the developmental process in nature[J]. Physiologia Plantarum,2010,138(3):312-320.
[2]Appenroth K J,Palharini L,Ziegler P. Low-molecular weight carbohydrates modulate dormancy and are required for post-germination growth in turions of Spirodela polyrhiza[J]. Plant Biology,2013,15(2):284-291.
[3] Appenroth K J,Augsten H. Photophysiology of turion germination in Spirodela polyrhiza (L.) Schleiden.Ⅴ. Demonstration of a calcium-requiring phase during phytochrome-mediated germination[J]. Photochemic Photobiology,1990,52(1):61-65.
[4] Newton R J,Shelton D R,Disharoon S,et al. Turion formation and germination in Spirodela polyrhiza[J]. American Journal of Botany,1978,65(4):421-428.
[5]Appenroth K J,Ziegler P. Light-induced degradation of storage starch in turions of Spirodela polyrhiza depends on nitrate[J]. Plant,Cell and Environment,2008,31(10):1460-1469.
[6] Appenroth K J,Keresztes A,Krzysztofowicz E,et al. Light-induced degradation of starch granules in turions of Spirodela polyrhiza studied by electron microscopy[J]. Plant and Cell Physiology,2011,52(2):384-391.
[7] Reimann R,Hippler M,Machelett B,et al. Light induces phosphorylation of glucan water dikinase,which precedes starch degradation in turions of the duckweed Spirodela polyrhiza[J]. Plant Physiology,2004,135(1):121-128.
[8] Perry T O. Dormancy,turion formation,and germination by different clones of Spirodela polyrrhiza[J]. Plant Physiology,1968,43(11):1866-1869.
[9] Lacor M A M. On the influence of gibberellic acid and kinetin on the germination of turions of Spirodela polyrhiza (L.) Schleiden [J]. Acta Botanica Neerlandica,1969,18(4):550-557.
[10]Malek L,Oda Y. Germination of Spirodela polyrhiza turions:the role of culture conditions during turion development[J]. Plant and Cell Physiology,1980,21( 2):357-361.
