果蠅鈉通道 DmNav22在雙氧木脂素A作用機理研究中的應用
2017-04-13 07:55:10閆秀芳郄杏桃吳文君胡兆農
西北農業學報 2017年4期
閆秀芳,郄杏桃,吳文君,胡兆農
(1. 西北農林科技大學 植物保護學院 農藥研究所,陜西楊凌 712100; 2. 西北農林科技大學 陜西省植物源農藥研究與開發重點實驗室,陜西楊凌 712100)
果蠅鈉通道 DmNav22在雙氧木脂素A作用機理研究中的應用
閆秀芳,郄杏桃,吳文君,胡兆農
(1. 西北農林科技大學 植物保護學院 農藥研究所,陜西楊凌 712100; 2. 西北農林科技大學 陜西省植物源農藥研究與開發重點實驗室,陜西楊凌 712100)
建立以爪蟾卵母細胞為載體的果蠅鈉通道表達系統,研究雙氧木脂素A的殺蟲分子機理。將果蠅鈉通道 DmNav22與TipE亞基基因分別進行體外轉錄,獲得 DmNav22與TipE亞基基因的cRNA,將 DmNav22與TipE的cRNA等質量混合,顯微注射至新鮮的非洲爪蟾卵母細胞,在ND96溶液、18 ℃培養12~18 h后采用雙電極電壓鉗技術記錄其電流,測定雙氧木脂素A對鈉電流的影響。結果表明,注射cRNA的非洲爪蟾卵母細胞,可記錄到明顯的果蠅鈉通道電壓門控內向電流,未注射的對照組細胞則未記錄到鈉電流;雙氧木脂素A使鈉通道失活電壓依賴性向超極化方向移動。說明,果蠅鈉通道在非洲爪蟾卵母細胞中成功表達,為進一步研究雙氧木脂素A的作用機理奠定基礎。
果蠅鈉通道;雙電極電壓鉗;非洲爪蟾卵母細胞;雙氧木脂素A
電壓門控鈉通道在電信號傳遞中起著關鍵作用,是許多天然神經毒素的靶標[1]。昆蟲鈉通道結構和哺乳動物相似,也由4個高度同源的結構域(DI-DIV)組成,每個結構域均含有6個跨膜α-螺旋片段(S1-S6)[2-3]。昆蟲第1個鈉通道基因是從果蠅的溫度敏感突變體克隆的para基因(命名為DmNav)[4]。TipE是昆蟲鈉通道的一個調節亞基基因,將para與TipE共同表達在爪蟾卵母細胞中不僅增強了表達的豐度,而且發現在不影響鈉電流的時間和電壓依賴性下,鋒電流增大,失活加快[5-6]。
非洲爪蟾卵母細胞被廣泛用于各型離子通道和受體的異源表達,用以研究受體及離子通道的生理學和藥理學特性,或探討藥物的作用機制和作用靶標[7]。迄今,已有4種DmNav同源基因在非洲爪蟾卵母細胞中功能性表達,即家蠅的Vssc1[8]、德國小蠊的BgNav[9-10]、瓦螨的VmNav[11]和黃熱病蚊伊蚊的aNav[12]基因。但在國內,爪蟾卵母細胞表達體系在昆蟲鈉通道研究方面的應用較少。
雙氧木脂素A(haedoxan A,HA)是日本學者Taniguchi等[13]從草本植物透骨草(PhrymaleptostachyaL.)中分離鑒定的一個二氧雙環辛烷木脂素類化合物。Taniguchi等[13]和肖新敏等[14-16]的殺蟲活性測定表明,HA具有超高的殺蟲活性,與合成擬除蟲菊酯類殺蟲劑的殺蟲活性處于同一水平。關于HA的殺蟲作用機理,Hu等[17]初步研究表明,HA能引起鈉通道失活曲線顯著地向超極化方向移動,推測其作用于電壓門控鈉通道。然而,HA作用鈉通道的機理及其作用位點如何,尚需進一步的研究。因此,本試驗建立以非洲爪蟾卵母細胞為載體的果蠅鈉通道表達系統,初步探討HA對昆蟲鈉通道作用的濃度效應,旨在為進一步研究HA的殺蟲作用機理奠定基礎。
1 材料與方法
1.1 供試材料
雙氧木脂素A(HA):從透骨草中分離獲得,純度(質量百分數)≥98%。其分子式C33H34O14,相對分子質量654,結構如圖1[13]。將HA溶于DMSO中制備成30 mmol/L的母液,4 ℃儲存備用,試驗時將其溶解于相應的ND96溶液中稀釋得到所需藥物終濃度。
菌種、質粒、試驗動物及培養基:大腸桿菌Stbl3購自廣州易錦生物技術有限公司;攜帶 DmNav22、TipE編碼序列的質粒由美國密歇根州立大學Dong Ke教授惠贈,表達載體為pGH19;性成熟的雌性非洲爪蟾購自中科院上海生命科學院生化細胞所干細胞技術平臺;大腸桿菌的培養基為LB液體或固體培養基。
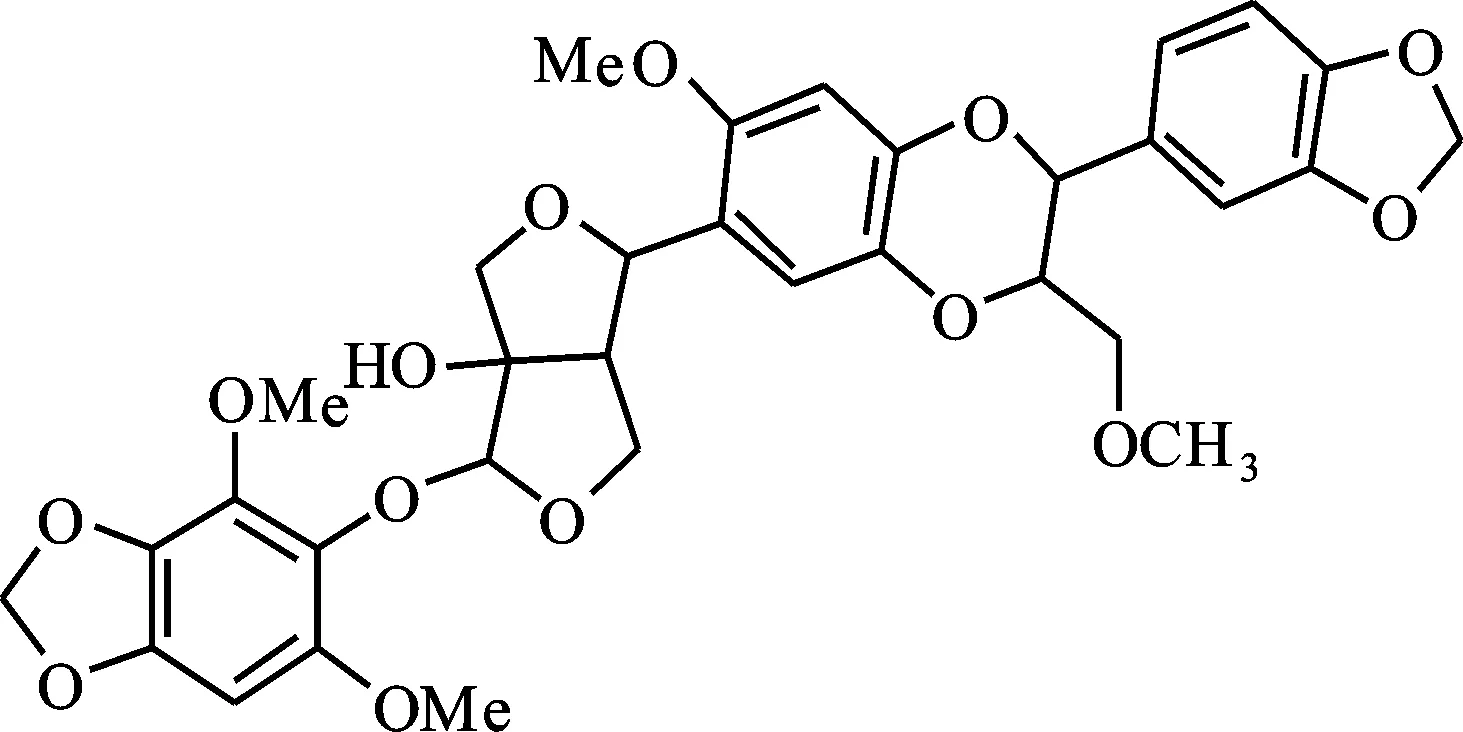
圖1 雙氧木脂素A結構Fig.1 Structure of haedoxan A
1.2 主要試劑及儀器
質粒小量抽提試劑盒、IA型膠原酶、限制性內切酶NotⅠ等常規分子生物學試劑購自Sigma公司;cRNA合成試劑盒mMESSAGE mMACHINE T7 Kit購自美國Ambion公司;檢測電流的電生理儀器為雙電極電壓鉗(Molecular Devices Corporation,型號:AXOCLAMP900A);微電極拉制儀(美國Sutter公司,P-97);體視顯微鏡(重慶重光實驗有限公司,型號:ZSA300);顯微注射儀(美國Sutter公司,Nanoliter2000)等。
1.3 記錄及培養液的配制
細胞浴液(ND96)成分:96 mmol/L NaCl,2 mmol/L KCl,1.8 mmol/L CaCl2,1 mmol/L MgCl2及5 mmol/L HEPES,用NaOH調節pH至7.5。
卵母細胞洗滌液:去除CaCl2的ND96液,用于洗滌及消化細胞。
卵母細胞培養液:加入2.5 mmol/L丙酮酸鈉、0.5 mmol/L茶堿以及50 μg/mL慶大霉素的ND96液,用于培養卵母細胞。
卵母細胞記錄液:ND96液作為電流記錄時的記錄液。
1.4 質粒轉化、提取及線性化
將含有 DmNav22、TipE基因的表達載體pGH19轉化大腸桿菌Stbl3,涂于含有氨芐(100 μg/mL)的固體LB培養基平板上,倒置平板,37 ℃培養箱中培養17~24 h,挑選單克隆置于液體LB培養基中,250 r/min振蕩培養12~16 h,提取質粒,經雙酶切鑒定正確后,送上海立菲生物測序有限公司鑒定,測序結果與已知數據比對正確。取 DmNav22和TipE的cDNA用NotⅠ限制性內切酶線性化2 h,線性化質粒電泳檢測并純化后,用作體外轉錄合成cRNA的模板。酶切反應體系參照說明書進行設定。
1.5 cRNA的制備及純化
純化過的線性化 DmNav22、TipE質粒,用T7體外轉錄試劑盒合成cRNA,操作步驟參照說明書進行,37 ℃反應2 h。轉錄后的 DmNav22、TipEcRNA用氯化鋰沉淀法進行純化。用分光光度計測定純化后cRNA的質量濃度和A260/A280,然后分裝,于-80 ℃保存。
1.6 非洲爪蟾卵母細胞的分離及cRNA的顯微注射
將6個月以上未進行試驗的性成熟雌性非洲爪蟾浸在2 g/L的三卡因溶液中10~30 min使之麻醉,在麻醉狀態下,于下腹部正中偏外0.5 cm左右縱向切開一約1 cm的小口,剪下3~5個卵巢瓣。在含1 mg/mL IA型膠原酶的卵母細胞洗滌液中18 ℃下消化40 min。用卵母細胞洗滌液反復清洗后置于卵母細胞培養液中。在解剖顯微鏡下挑出黑白半球界限分明、發育階段處于Ⅴ~Ⅵ期且表面光滑、顆粒飽滿的卵母細胞備用。用微量注射器將等質量混合的 DmNav22、TipE的cRNA,注入挑選好的卵母細胞內。注射后的卵母細胞置于細胞培養液中,18 ℃恒溫培養12~18 h后用于電流檢測。試驗過程中所有器具需要預先進行滅菌處理,卵母細胞培養液每12 h 更換1次,且需及時在解剖顯微鏡下將不健康或死去的細胞挑出丟棄。
1.7 電流記錄
電流記錄及數據分析參照Du等[18]的方法。利用雙電極電壓鉗系統測定果蠅鈉通道電流。使用微電極拉制儀拉制玻璃微電極,灌充含瓊脂粉(5 g/L)的3 mol/L KCl溶液,電極電阻小于0.5 MΩ。將卵母細胞置于容積約1 mL底部正中粘有一小片網狀薄膜的記錄槽中,使用微操動儀分別將電流電極與電壓電極同時插入細胞中,設置鉗制電壓為-120 mV,給予細胞去極化刺激,室溫下記錄果蠅鈉通道電流,同時以沒有注射cRNA的卵母細胞為對照。
HA母液(30 mmol/L)用ND96記錄液稀釋成終濃度為0.3和3 μmol/L測試溶液。φ(DMSO)<0.5%,這樣試驗時不影響鈉通道功能特征。藥劑應用參照Tan等[19]所描述的方法。HA加入10 min后開始記錄。
穩態激活電壓依賴性測定,卵母細胞鉗置在-120 mV,從-80 mV去極化至65 mV,階躍電壓是5 mV。以經過標準化的電導G/Gmax來反映通道激活情況。G=I/(U-Urev),其中I是電流值,U是膜電壓(也就是記錄到I時的測試電壓),Urev是鈉電流的反轉電壓。通道激活數據擬合兩元Boltzmann方程G/Gmax=[1+exp(U-U1/2)/K]-1,其中U是膜電壓,U1/2是半激活電壓,K是斜率。
穩態失活電壓依賴性測定,卵母細胞先施予一組從-120 mV至40 mV的預前刺激,時間200 ms,階躍電壓是5 mV,接著再給予一組到-10 mV的測試刺激,時間20 ms。以經過標準化的電流值I/Imax來反應通道失活情況。通道失活數據擬合兩元Boltzmann方程I/Imax=[1+exp(U-U1/2)/K]-1,其中I是電流值,Imax是最大電流值,U是失活電壓,U1/2是半失活電壓,K是斜率[17]。
1.8 數據采集和分析
數據分析采用SPSS 12.0(version 12.0,SPSS Inc.,Chicago,IL,USA,2003)和Origin 8.0(Origin Lab Corp,Northampton,MA,USA,2013);所有數據均以“平均數±標準差”表示。配對樣本t-test法進行顯著性差異分析,P≤0.05認為差異性顯著。
2 結果與分析
2.1 DmNav22、TipE質粒的酶切線性化
對所提質粒 DmNav22、TipE用NotⅠ進行酶切線性化。酶切2 h,分別取2 μL酶切液于10 g/L的瓊脂糖凝膠電泳。如圖2所示,酶切后DNA為1條帶,說明酶切完全。
2.2 DmNav22、TipE cRNA質量濃度測定
以純化的線性化 DmNav22、TipE質粒為模板,進行體外轉錄合成 DmNav22、TipE的cRNA。取1 μL純化后的cRNA用紫外分光光度計測定其質量濃度及A260/A280, DmNav22、TipEcRNA的質量濃度分別為645 ng/μL和1 712 ng/μL,本試驗中A260/A280測值均在2.0附近,表明cRNA無蛋白質污染(表1)。符合后續卵母細胞注射對cRNA的要求。

M為DNA marker,1和2分別代表TipE和 DmNav22的線性化質粒 M is DNA marker,1 and 2 respectively reprent linearized plasmids ofTipEand DmNav22
圖2 DmNav22和TipE的線性化質粒
Fig.2 Linearized plasmids of DmNav22 andTipE
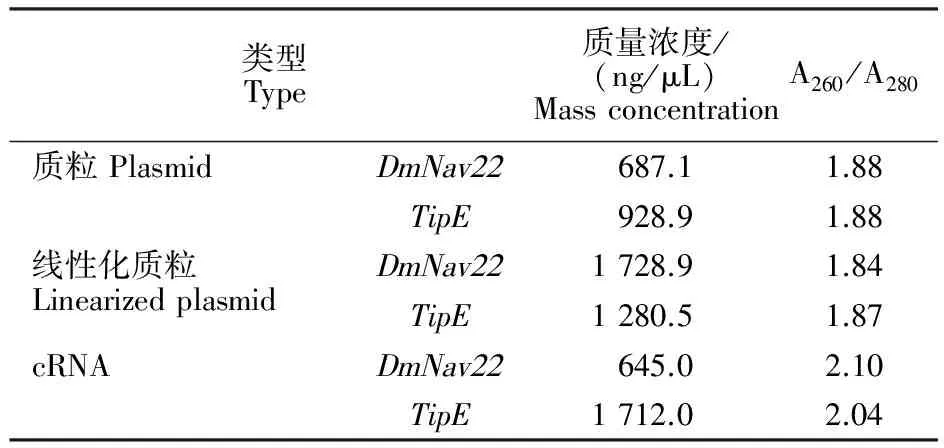
表1 DmNav22和TipE的質粒、線性化質粒及cRNA的質量濃度和A260/A280Table 1 Mass concentration and A260/A280 of DmNav22 and TipE plasmids,linearized DNAs and cRNAs
2.3 果蠅鈉通道在非洲爪蟾卵母細胞中的功能性表達
經雙電極電壓鉗系統測定果蠅鈉通道電壓門控電流發現,未注射空白對照組的卵母細胞,進行脈沖激活時,沒有記錄到鈉電流,而注射cRNA的卵母細胞進行脈沖激活時,產生明顯的內向電流(圖3-A)。大約在-50 mV時檢測到果蠅鈉通道電流,-15 mV左右達到最大值(圖3-B)。穩態激活電壓依賴性測定,卵母細胞鉗置在-120 mV,從-80 mV去極化至65 mV,階躍電壓是5 mV。穩態激活電壓依賴性數據擬合曲線如圖3-C。半激活電壓(U1/2)是(-31.6±2.0) mV,斜率(K)是(5.0±0.6) mV(表2)。穩態失活電壓依賴性測定,卵母細胞先施予一組從-120 mV至40 mV的預前刺激,時間200 ms,階躍電壓是5 mV,接著再給予一組到-10 mV的測試刺激,時間20 ms。穩態失活電壓依賴性數據擬合曲線如圖3-D。半失活電壓(U1/2)是(-47.5±1.3) mV,斜率(K)是(5.2±0.3) mV(表2)。
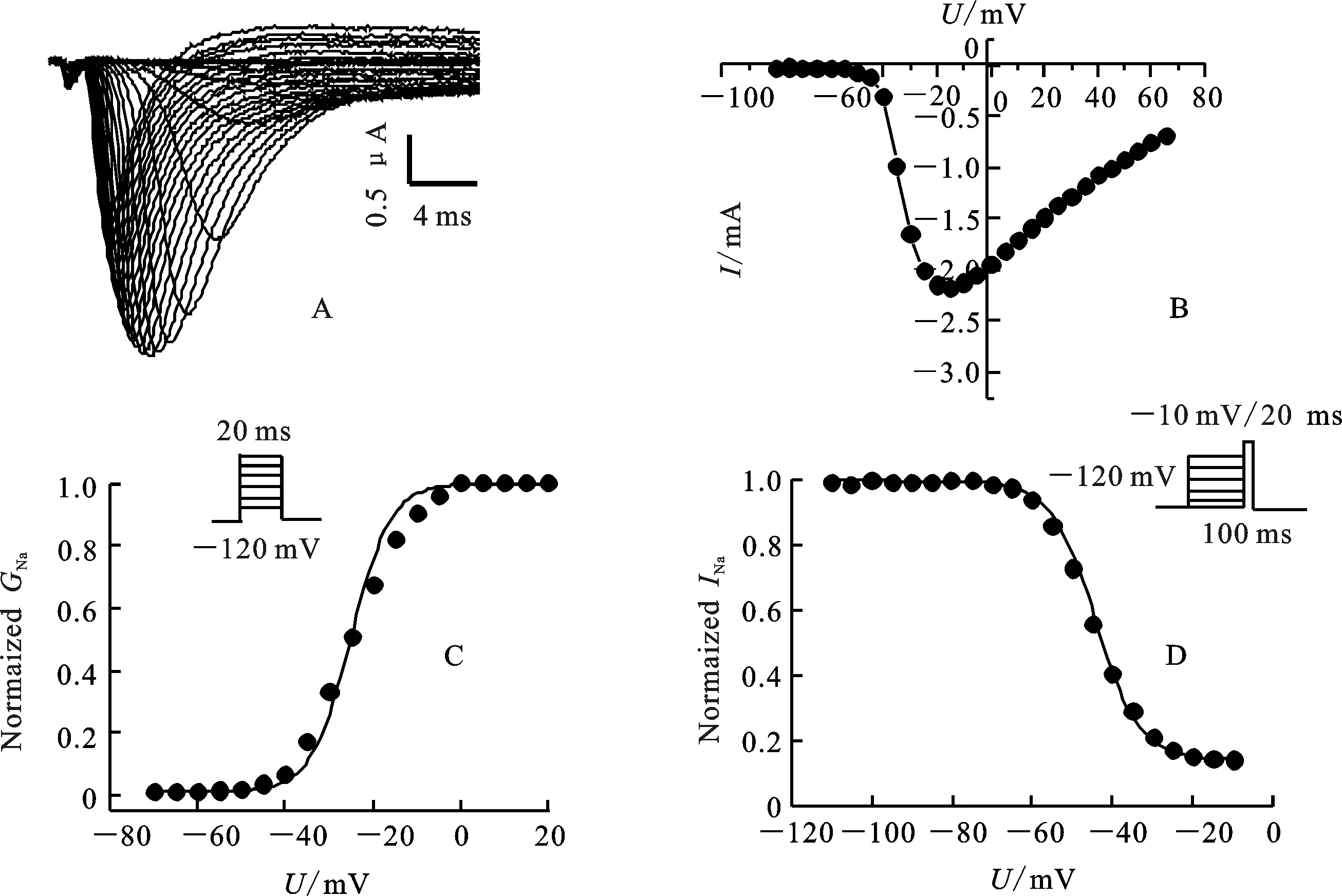
A:鈉電流曲線。鈉電流測定,卵母細胞鉗置在-120 mV,從-80 mV去極化至65 mV,階躍電壓5 mV;B:電流電壓曲線(I-U),電流由圖3-A記錄,峰值電流對應去極化電壓作圖,細胞數n=5;C:果蠅鈉通道活化電壓依賴性,細胞數n=3;D:果蠅鈉通道失活電壓依賴性,細胞數n=3。
A:Sodium current traces. Sodium currents were recorded from a holding potential of -120 mV during depolarizations from -80 to 65 mV in 5 mV increments.B:Sodium current-voltage relation (I-Ucurve). Current were recorded as described for Fig.3-A,and the peak current amplitude is plotted versus the depolarization potential. The number of oocytes was 5.The voltage-dependence of activation C and inactivation D ofDrosophilasodium channels. The number of oocytes was 3.
圖3 果蠅鈉通道在爪蟾卵母細胞中的功能性表達
Fig.3 Functional expression ofDrosophilasodium channels in Xenopus oocytes
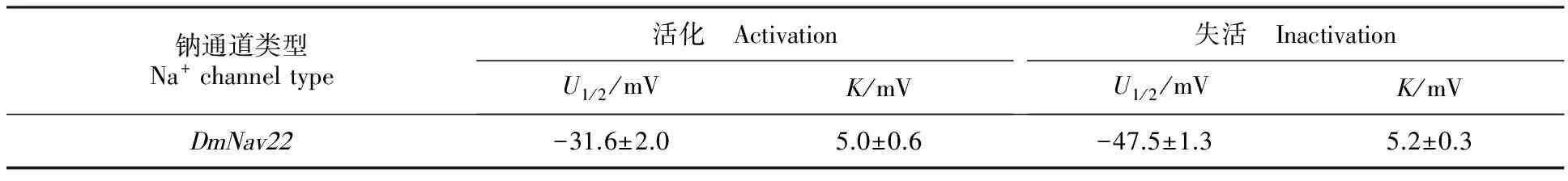
表2 果蠅鈉通道 DmNav22活化、失活電壓依賴性參數Table 2 Parameters of voltage dependence of activation and inactivation of Drosophila DmNav22 sodium channels
注:U1/2為活化或失活半最大電壓,K為活化或失活斜率,細胞數n=5。
Note:U1/2is the half-maximal voltage for activation or inactivation andKis the slope factor for activation or inactivation.Five oocytes.
2.4 HA對鈉通道失活電壓依賴性的影響
穩態失活電壓依賴性測定,卵母細胞先施予一組從-120 mV至40 mV的預前刺激,時間200 ms,階躍電壓是5 mV,接著再給予一組到-10 mV的測試刺激,時間20 ms。結果表明,0.3 μmol/L和3 μmol/L HA均使鈉通道失活電壓向超極化方向移動(圖4)。0.3 μmol/L和3 μmol/L 藥劑分別使鈉通道失活半最大電壓向超極化方向移動1.1 mV(0.3 μmol/L)和2.9 mV(3 μmol/L)(表3)。
3 討 論
本試驗成功建立以非洲爪蟾卵母細胞為載體的果蠅鈉通道體外表達系統,并利用此系統測定HA對鈉通道失活門控特性的影響。未注射空白對照組的卵母細胞,進行脈沖激活時,沒有記錄到鈉電流,注射cRNA的卵母細胞進行脈沖激活時,產生明顯的內向電流。試驗測得的通道活化半最大電壓(U1/2)、斜率(K)及通道失活半最大電壓(U1/2)、斜率(K)與文獻報到的基本一致[17],證明以非洲爪蟾卵母細胞為載體的果蠅鈉通道體外表達系統成功建立。
HA對鈉通道失活門控測定結果表明,0.3 μmol/L和3 μmol/L HA均可以使鈉通道失活電壓向超極化方向移動,其趨勢與文獻報道[17]一致。失活電壓移動幅度隨藥劑濃度的增加而增大,表明HA對鈉通道失活門控的影響具有濃度依賴性。
國內外新農藥研究與開發的實踐證明,從植物中提取、分離殺蟲活性成分,并以這些活性成分為探針發現新靶標是新農藥創制的重要途徑[20]。HA是從透骨草中分離鑒定出的一種具有二氧雙環辛烷新骨架的木脂素類化合物,殺蟲活性高,試蟲攝毒后的主要中毒癥狀為拒食,肌肉松弛,最終死亡[13]。鑒于其較高的殺蟲活性、新穎的結構、獨特的作用癥狀,對HA殺蟲作用機理的探討有助于推動中國新型生物合理殺蟲劑的創制。
以非洲爪蟾卵母細胞為載體的果蠅鈉通道體外表達系統的建立,為從分子水平探討HA在鈉通道上的受體位點奠定基礎,不僅有利于進一步獲得鈉通道的一些結構特征,而且有利于闡明HA可能的分子作用機理,從而可在對HA進行三維定量構效關系研究的基礎上,設計出新的高效殺蟲劑。
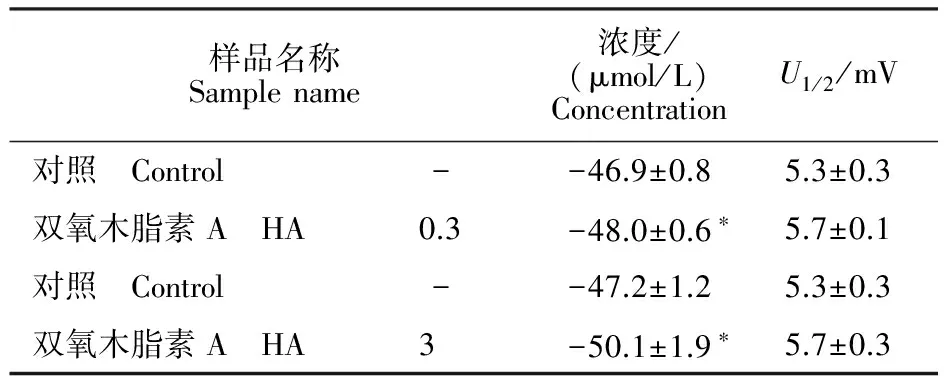
表3 HA對果蠅鈉通道穩態失活參數的影響Table 3 Effects of HA on steady-state inactivation parameters for Drosophila sodium channels
注:U1/2為半最大失活電壓,K為失活斜率,細胞數n=3。*表示對照組與處理組差異顯著(P≤0.05)。
Note:U1/2is the half-maximal voltage for inactivation andKis the slope factor for inactivation.Each value represents the mean ± SEM for three oocytes. Asterisks indicate significant differences between control and treatment(P≤0.05).
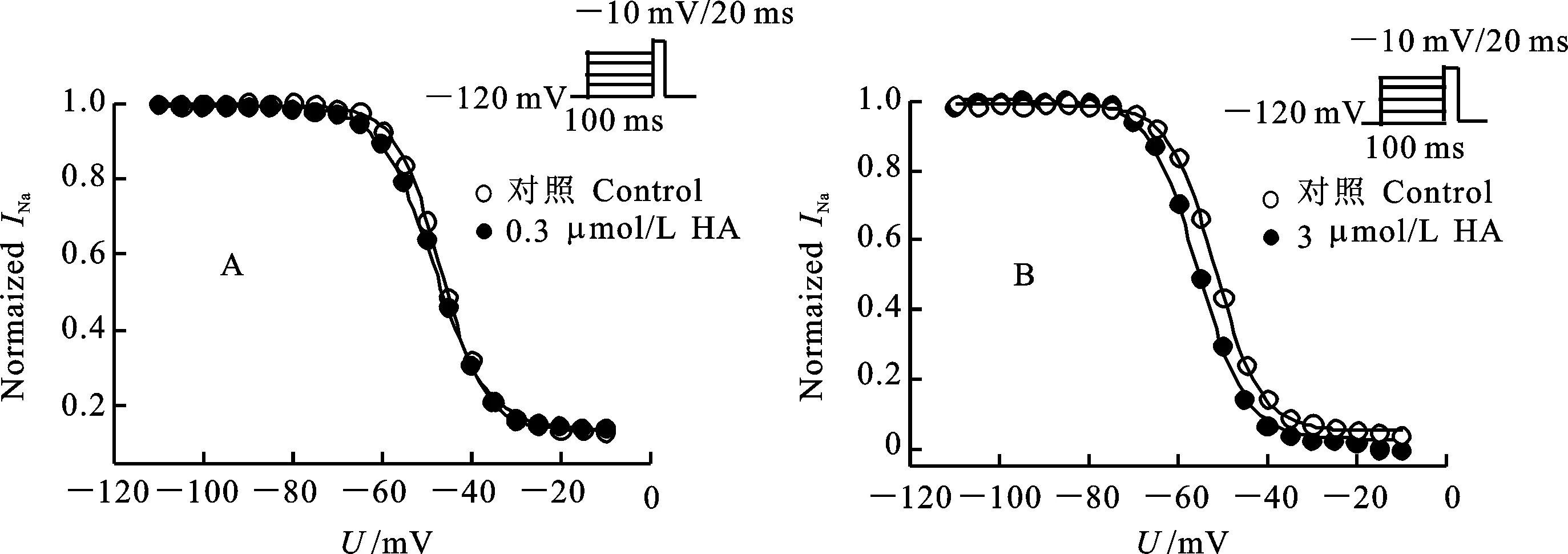
A:0.3 μmol/L HA加藥前后果蠅鈉通道失活電壓依賴性;B:3 μmol/L HA加藥前后果蠅鈉通道失活電壓依賴性,失活曲線擬合兩元Boltzmann方程,細胞數n=3。
A and B:the voltage dependence of inactivation ofDrosophilasodium channels before and after the application of 0.3 μmol/L (A) and 3 μmol/L (B) of HA. The inactivation curves were fitted with two-state Boltzmann equation. The number of oocytes was 3.
圖4 HA引起果蠅鈉通道失活電壓依賴性向超極化方向移動
Fig.4 HA induced a hyperpolarization direction shift of the voltage dependence of inactivation ofDrosophilasodium channels
Reference:
[1] 王洪躍,魏樹源.昆蟲鈉通道的研究及其應用[J].中國醫學裝備,2009,6(3):13-16.
WANG H Y,WEI SH Y.Researches of sodium channels in insects[J].ChinaMedicalEquipment,2009,6(3):13-16 (in Chinese with English abstract).
[2] DONG K,DU Y Z,RINKEVINCH F,etal.Molecular biology of insect sodium channels and pyrethroid resistance[J].InsectBiochemistry&MolecularBiology,2014,50:1-17.
[3] RINKEVINCH F D,DU Y Z,DONG K.Diversity and convergence of sodium channel mutations involved in resistance to pyrethroids[J].PesticideBiochemistry&Physiology,2013,106(3):93-100.
[4] LOUGHNEY K,KREBER R,GANETZKY B.Molecular analysis of the para locus,a sodium channel gene inDrosophila[J].Cell,1989,58(6):1143-1154.
[5] WARMKE J W,REENAN R A,WANG P,etal.Functional expression ofDrosophilapara sodium channels.Modulation by the membrane protein TipE and toxin pharmacology[J].JournalofGeneralPhysiology,1997,110(2):119-133.
[6] FENG G,DEAK P,CHOPRA M,etal.Cloning and functional analysis ofTipE,a novel membrane protein that enhancesDrosophilapara sodium channel function[J].Cell,1995,82(6):1001-1011.
[7] 劉曉燕,鄭建全,艾 平,等.爪蟾卵母細胞雙電極電壓鉗記錄技術及其內源性外向電流的研究[J].軍事醫學科學院院刊,2005,29(5):460-463.
LIU X Y,ZHENG J Q,AI P,etal.Two-electrode voltage clamp recording technique forXenopuslaevisoocytes and its endogenous outward currents[J].BulletinoftheAcademyofMilitaryMedicalSciences,2005,29(5):460-463 (in Chinese with English abstract).
[8] SMITH T J,SODERLUND D M.Action of the pyrethroid insecticide cypermethrin on rat brain IIa sodium channels expressed inXenopusoocytes[J].Neurotoxicology,1998,19(6):823-832.
[9] LIU Z,VALLES S M,DONG K.Novel point mutations in theGermancockroachpara sodium channel gene are associated with knockdown resistance (kdr) to pyrethroid insecticides[J].InsectBiochemistryandMolecularBiology,2000,30(10):991-997.
[10] TAN J,LIU Z Q,TSAI T D,etal.Novel sodium channel gene mutations inBlattellagermanicareduce the sensitivity of expressed channels to deltamethrin.[J].InsectBiochemistry&MolecularBiology,2002,32(4):445-454.
[11] DU Y Z,NOMURA Y,LIU Z Q,etal.Functional expression of an arachnid sodium channel reveals residues responsible for tetrodotoxin resistance in invertebrate sodium channels[J].JournalofBiologicalChemistry,2009,284(49):33869-33875.
[12] DU Y Z,NOMURA Y,STAR G,etal.Molecular evidence for dual pyrethroid-receptor sites on a mosquito sodium channel[J].ProceedingsoftheNationalAcademyofSciences,2013,110(29):11785-11790.
[13] TANIGUCHI E,IMAMURA K,ISHIBASHI F,etal.Structure of the novel insecticidal sesquilignan,haedoxan A[J].Agricultural&BiologicalChemistry,1989,53(3):631-643.
[14] XIAO X M,HU ZH N,SHI B J,etal.Larvicidal activity of lignans fromPhrymaleptostachyaL.againstCulexpipienspallens[J].ParasitologyResearch,2012,110(3):1079-1084.
[15] XIAO X M,JI Z Q,ZHANG J W,etal.A new lignan fromPhrymaleptostachya[J].ChemistryofNaturalCompounds,2013,49(1):21-23.
[16] 肖新敏,胡兆農,姬志勤,等.透骨草殺蟲活性成分分離鑒定及其生物活性研究[J].農藥學學報,2012,14(5):583-586.
XIAO X M,HU ZH N,JI ZH Q,etal.Isolation,structure identification and bioactivity of active ingredients fromPhrymaleptostachya[J].ChineseJournalofPesticideScience,2012,14(5):583-586 (in Chinese with English abstract).
[17] HU ZH N,DU Y Z,XIAO X M,etal.Insight into the mode of action of Haedoxan A fromPhrymaleptostachya[J].Toxins,2016,8(2):53.
[18] DU Y Z,NOMURA Y,LUO N,etal.Molecular determinants on the insect sodium channel for the specific action of type Ⅱ pyrethroid insecticides[J].Toxicology&AppliedPharmacology,2009,234(2):266-272.
[19] TAN J G,LIU Z Q,WANG R W,etal.Identification of amino acid residues in the insect sodium channel critical for pyrethroid binding[J].MolecularPharmacology,2005,67(2):513-522.
[20] 肖新敏.透骨草殺蟲活性成分及其作用機理研究[D].陜西楊凌:西北農林科技大學,2014.
XIAO X M.Studies on insecticidal activity components and reaction mechanisms ofPhrymaleptostachyaL.[D].Yangling Shaanxi:Northwest A&F University,2014 (in Chinese with English abstract).
(責任編輯:郭柏壽 Responsible editor:GUO Baishou)
Application ofDrosophilaSodium Channel DmNav22 in Investigating the Action Mode of Haedoxan A
YAN Xiufang,QIE Xingtao,WU Wenjun and HU Zhaonong
(1.Institute of Pesticide Science,College of Plant Protection,Northwest A&F University,Yangling Shaanxi 712100,China; 2.Shaanxi Province Key Laboratory of Botanical Pesticide Research and Development,Northwest A&F University,Yangling Shaanxi 712100,China)
To investigate the molecular mechanism of haedoxan A (HA) against insect pests,gene expression system ofDrosophilasodium channel inXenopuslaevisoocytes was established. cRNAs ofDrosophilasodium channel subunits DmNav22 andTipEwere obtained by transcription in vitro,which were mixed with equal amount of substance and then microinjected in the freshX.laevisoocytes. The oocytes were incubated in ND96 buffer at 18 ℃ for 12-18 hours after microinjection and the sodium channel current was detected by two-electrode voltage clamp (TEVC). The results showed that the voltage-gated inward current was recorded in the oocytes injected cRNA,while no current was recorded in the blank oocytes. HA shifted the voltage dependence ofDrosophilasodium channels inactivation to the hyperpolarizing direction. These results demonstrated that theDrosophilasodium channel was expressed inX.laevisoocytes successfully and can be used to further study the insecticidal mechanism of HA.
Drosophilasodium channel; Two-electrode voltage clamp;Xenopuslaevisoocytes; Haedoxan A (HA)
YAN Xiufang,female,master student. Research area:pesticide toxicology. E-mail:xiufangyan@163.com
HU Zhaonong,male,Ph.D,professor,doctoral supervisor. Research area:pesticide toxicology. E-mail:huzhaonong@nwsuaf.edu.cn
日期:2017-03-30
2016-03-16
2016-05-08
國家自然科學基金(31672055);陜西省農業科技創新與攻關(2015NY033)。
閆秀芳,女,碩士研究生,從事農藥毒理學研究。E-mail:xiufangyan@163.com
胡兆農,男,博士,教授,博士生導師,主要從事農藥毒理學研究。E-mail:huzhaonong@nwsuaf.edu.cn
S481
A
1004-1389(2017)04-0635-06
網絡出版地址:http://kns.cnki.net/kcms/detail/61.1220.S.20170330.1509.040.html
Received 2016-03-16 Returned 2016-05-08
Foundation item The National Natural Science Foundation of China (No.31672055); Agricultural Science and Technology Key Project of Shaanxi Province(No.2015NY033).
