增加煙草一級和二級側(cè)根是抵御干旱的生理機制
2017-04-14 05:28:44趙陽王樹聲張亞麗薛韌王程棟
植物營養(yǎng)與肥料學(xué)報 2017年2期
趙陽,王樹聲,張亞麗,薛韌,王程棟*
(1 中國農(nóng)業(yè)科學(xué)院煙草研究所,山東青島 266100;2 南京農(nóng)業(yè)大學(xué)資源與環(huán)境科學(xué)學(xué)院,南京 210095)
增加煙草一級和二級側(cè)根是抵御干旱的生理機制
趙陽1,王樹聲1,張亞麗2,薛韌2,王程棟1*
(1 中國農(nóng)業(yè)科學(xué)院煙草研究所,山東青島 266100;2 南京農(nóng)業(yè)大學(xué)資源與環(huán)境科學(xué)學(xué)院,南京 210095)
【目的】挖掘植物自身的抗旱能力是提高農(nóng)業(yè)水分利用效率的有效措施。本研究通過比較水分脅迫處理下 5 個烤煙品種苗期根系發(fā)生和伸長以及葉片光合特性的變化,探討烤煙抗旱能力差異的生理機制,為煙草生產(chǎn)的可持續(xù)發(fā)展提供理論依據(jù)。【方法】采用水培試驗,設(shè)置對照和水分脅迫處理 (添加 2.5% PEG 處理來模擬中度干旱),研究 5 個烤煙品種苗期的地上部和根系的生物量、一級和二級側(cè)根的發(fā)生和伸長,利用 Li-Cor 6400光合測定系統(tǒng)測定葉片的光合速率和氣孔導(dǎo)度等參數(shù),利用 FLIR 手持式紅外熱成像儀測定新展開葉的葉片溫度,分析煙草抗旱能力差異明顯的生理機制。【結(jié)果】水分脅迫條件下 5 個烤煙品種的生長差異明顯。與對照處理相比,水分脅迫對大平板和農(nóng)大 202 的生長影響不明顯,但柳葉尖、金星 6007 和中煙 100 的地上部和根系生物量顯著被抑制,降幅范圍在 66%~83%,達到差異極顯著水平。水分脅迫抑制 5 個烤煙品種一級和二級側(cè)根的伸長,但大平板和農(nóng)大 202 的一級側(cè)根數(shù)目和二級側(cè)根密度顯著增加,最終導(dǎo)致根系生物量沒有下降;另外 3 個煙草品種柳葉尖、金星 6007 和中煙 100 的側(cè)根發(fā)生增加趨勢不明顯。水分脅迫處理下抗旱能力強的大平板和農(nóng)大 202 地上部生物量和葉片溫度變化不明顯,而葉片的凈光合速率顯著下降,但其降幅顯著低于 3 個抗旱能力弱的烤煙品種。與對照處理相比,抗旱能力弱的柳葉尖、金星 6007 和中煙 100 葉片的氣孔導(dǎo)度和蒸騰速率顯著下降,葉片溫度上升 5~7℃。進一步的研究表明,PEG處理下耐旱能力強烤煙品種側(cè)根數(shù)目的顯著增加是地上部維持較強光合能力的主要原因。【結(jié)論】抗旱能力強的煙草品種苗期受到水分脅迫時其一級和二級側(cè)根顯著增加,進而能保持較高的光合特性。
煙草;干旱脅迫;側(cè)根發(fā)生;光合特性
我國水資源不足是影響農(nóng)業(yè)可持續(xù)發(fā)展的重要因素。煙草作為我國重要的經(jīng)濟作物,起源于降雨充沛的熱帶,整個生育期對水分的需求都很高。在煙草生長季節(jié)中,倘若田間持水量未能保持在 50%以上,就會嚴重影響煙葉產(chǎn)量和品質(zhì)[1]。在西南丘陵山區(qū)種植煙草往往能夠生產(chǎn)出高質(zhì)量的煙葉[1],但該地區(qū)煙田土壤極易出現(xiàn)水分虧缺[2],因此干旱脅迫成為煙草生長的重要限制因子。
在干旱脅迫條件下,作物感受水分脅迫并通過信號傳導(dǎo),進而啟動生理生化反應(yīng)來響應(yīng)和適應(yīng)水分脅迫[3-5]。根系是植物吸收水分的主要器官,最先感受到土壤水分的虧缺[6],因此根系對于水分虧缺的響應(yīng)對植物的耐旱能力具有重要作用。目前干旱對根系形態(tài)影響的研究主要集中在豌豆[7]、小麥[8]和水稻[9]等作物上,對不同煙草品種抗旱能力的研究主要集中在葉片的光合及生理生化反應(yīng)等指標[10-13],較少涉及煙草根系的發(fā)生和伸長與抗旱能力的關(guān)系。在水稻上的研究表明[14-15],在育秧階段采取不同的水分管理方式對秧苗素質(zhì)影響顯著,與濕潤育秧相比旱育秧的秧苗根系具有很強的生理優(yōu)勢,主要表現(xiàn)在根數(shù)增多和根系活力增強。近年來煙草漂浮育苗已成為主要育苗方式,煙草漂浮育苗后期的“斷水”煉苗,就是讓煙苗處于適當缺水的條件,經(jīng)過抗旱鍛煉的煙苗更能適應(yīng)以后的干旱環(huán)境。移栽前的“斷水”煉苗能顯著增加煙苗素質(zhì),提高移栽成活率。我們在生產(chǎn)實踐中發(fā)現(xiàn)“斷水”煉苗增加煙草秧苗素質(zhì)的效應(yīng)因煙草品種而異,但其機制并不清楚。本研究采用 2.5% 聚乙二醇 (PEG) 溶液模擬中度干旱[16]條件研究了 5 個烤煙品種一級和二級側(cè)根的發(fā)生和伸長,發(fā)現(xiàn)在水分脅迫條件下抗旱能力強的烤煙品種的側(cè)根發(fā)生顯著增加;在此基礎(chǔ)上還研究了水分脅迫條件對烤煙的光合特性及葉片溫度的影響,進一步闡明抗旱能力強煙草的生理特征,為明晰煙草抗旱的生理機制并為其科學(xué)的育秧方式提供堅實的理論基礎(chǔ)。
1 材料與方法
1.1 試驗材料
根據(jù) 文 獻[13,17-18]報道,農(nóng)大 202 為抗旱烤煙 品 種 ,金星 6007 和中煙 100 為抗旱能力弱的烤煙品種;為了讓研究更具普遍意義,利用更多遺傳背景的烤煙品種來闡明煙草的抗旱機制,除了上述的三個烤煙品種外,前期試驗又增加了 10 個烤煙品種來研究它們的抗旱能力,最終選出 5 個抗旱能力差異大的烤煙品種作為深入研究的供試材料。在本實驗研究中,2 個抗旱能力強的烤煙品種 (農(nóng)大 202 和大平板)和 3 個抗旱能力弱的烤煙品種 (柳葉尖、金星 6007和中煙 100) 為供試材料,試驗在南京農(nóng)業(yè)大學(xué)日光溫室進行。所用材料由國家農(nóng)作物種質(zhì)資源平臺煙草種質(zhì)資源子平臺 (中國農(nóng)業(yè)科學(xué)院煙草研究所) 提供。
1.2 試驗方法
煙草種子消毒后在無菌水中黑暗催芽,露白后播種到基質(zhì) (泥炭和蛭石比例為 1∶1) 中培養(yǎng)約 25天。選取長勢均勻一致的幼苗移栽到 7 L 周轉(zhuǎn)箱,營養(yǎng)液為 1/4 濃度的霍格蘭營養(yǎng)液。每個周轉(zhuǎn)箱里面培養(yǎng) 10 株煙苗,緩苗 3 天后開始設(shè)置處理:對照處理和 2.5% PEG 處理 (滲透勢約為-0.08 Mpa,相當于中度干旱)。每個處理重復(fù) 8 次,并進行 3 次獨立的生物學(xué)重復(fù)。為了進一步明確側(cè)根數(shù)目的增加是抗旱能力強的煙草的重要生理表現(xiàn),以農(nóng)大 202 為試材,設(shè)置三個處理 (對照、PEG 和 PEG + 剪根處理),PEG + 剪根處理是在實施試驗的過程中通過剪根使 PEG 處理的煙苗一級側(cè)根的數(shù)目和對照處理保持一致。營養(yǎng)液 pH 值每天調(diào)至 5.5 左右,每兩天更換一次營養(yǎng)液。處理 7 天后,采收試驗材料并測定各項指標。
1.3 測定項目
1.3.1生物量 采樣分地上部和根系,于 105℃ 烘箱中烘 3 天至恒重后稱其重量。
1.3.2總根長及根系總體積 根系形態(tài)參數(shù)采用 LA1600+根系掃描儀成像。成像前,于成像掃描儀配套的凹槽中選擇大小接近根系的加約 1/2 水,將根系浸沒其中,用鑷子將根系分開并盡量減少對根系的傷害。成像后,選擇目標根系區(qū)域,通過 WinRHizo2003b分析圖像參數(shù),測定總根長和根系體積。
1.3.3一級側(cè)根和二級側(cè)根 于成像掃描儀配套的凹槽中選擇大小接近根系的加約 1/2 水,將根系浸沒其中,用鑷子將根系分開,測量一級側(cè)根長度,計算一級側(cè)根數(shù)量。用已知的總根長減去一級側(cè)根長度,即得二級側(cè)根長度。選取最長的三條一級側(cè)根,計算每條一級側(cè)根上的二級側(cè)根數(shù)。用一級側(cè)根長度除以二級側(cè)根數(shù),即得二級側(cè)根密度。
1.3.4葉片光合特性 釆用 Li-Cor 6400 光合測定系統(tǒng)測定新完全展開葉的凈光合速率 (Pn),測定時間為9:30~11:00,CO2濃度為 (385 ± 5) μmol/mol,測定光強設(shè)定為 1500 μmol/(m2·s)、流量設(shè)定為 500 mL/s,葉片溫度為 (25.5 ± 2) ℃,相對濕度維持在 45% 左右。1.3.5葉片溫度 采用 FLIR 手持式紅外熱成像儀對新完全展開葉進行拍照,再通過 FLIR Tools 軟件進行圖片處理從而提取數(shù)據(jù)。
1.4 數(shù)據(jù)處理
數(shù)據(jù)采用 SPSS22 軟件進行 ANVOA 方差分析和多重比較,文中的數(shù)據(jù)表示為平均值 ± SD。
2 結(jié)果與分析
2.1 生物量
從圖 1 可看出,水分脅迫條件下 2 個耐旱能力強與 3 個耐旱能力弱的煙草品種的生長差異明顯。與對照處理相比,大平板和農(nóng)大 202 地上部的生物量分別下降 18% 和 17%,根系生物量分別下降 14%和 6%,但差異未達顯著水平;柳葉尖、金星 6007和中煙 100 生長受到顯著抑制,地上部和根系的降幅在 66%~83%,差異達到極顯著水平。
2.2 根系形態(tài)
2.2.1總根長和根系體積 水分脅迫條件下,2 個耐旱能力強與 3 個耐旱能力弱的煙草品種的總根長和根系體積差異顯著 (圖 2)。與對照處理相比,大平板和農(nóng)大 202 的總根長和根系體積有下降的趨勢,但差異未達顯著水平;3 個抗旱能力弱的烤煙品種的總根長和根系體積顯著下降,總根長降幅在 47%~69%,根系體積降幅在 25%~43%,差異均達到顯著或極顯著水平。
2.2.2一級側(cè)根的發(fā)生和伸長 水分脅迫條件下,5個煙草品種的一級側(cè)根平均長度均顯著下降,但 3個抗旱能力弱的煙草品種的降幅遠高于 2 個抗旱能力強的煙草品種 (圖 3)。抗旱能力差異大的煙草的一級側(cè)根數(shù)目顯著增加,大平板和農(nóng)大 202 的增幅分別為 135% 和 109%,增幅達極顯著水平;3 個抗旱能力弱的烤煙品種的增幅僅在 12%~28%,增幅未達顯著水平。
2.2.3二級側(cè)根的發(fā)生和伸長 5 個烤煙品種二級側(cè)根的發(fā)生和伸長對水分脅迫的響應(yīng)和一級側(cè)根的變化類似 (圖 4)。水分脅迫條件下,5 個烤煙品種的二級側(cè)根平均長度均顯著降低。抗旱能力強的大平板和農(nóng)大 202 二級側(cè)根平均長度的降幅分別為 31% 和39%,3 個抗旱能力弱的烤煙品種的降幅顯著高于抗旱能力強的品種。5 個煙草品種的二級側(cè)根密度增加幅度不同,抗旱能力強的大平板和農(nóng)大 202 二級側(cè)根密度的增幅分別為 49% 和 41%,增幅達顯著水平,3 個抗旱能力弱的烤煙品種的二級側(cè)根密度增幅范圍在 8%~25%,未達顯著水平。
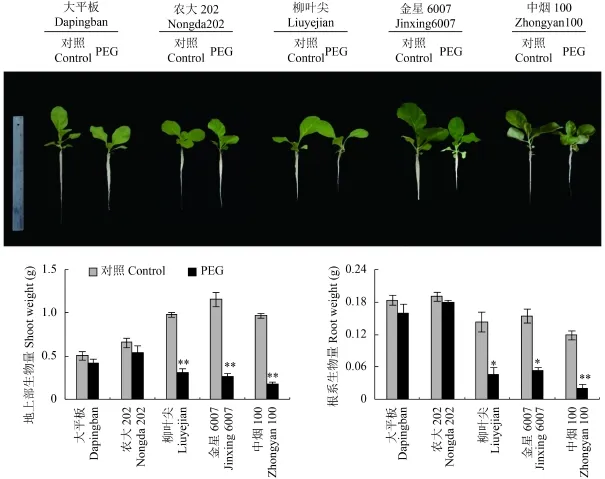
圖1 水分脅迫對煙草生長的影響Fig. 1 Tobacco growth in response to water stress[注(Note):* 和 ** 分別表示處理間在 0.05 和 0.01 水平差異差異顯著Indicate the significant differences between different treatments at the 0.05 and 0.01 levels,respectively.]
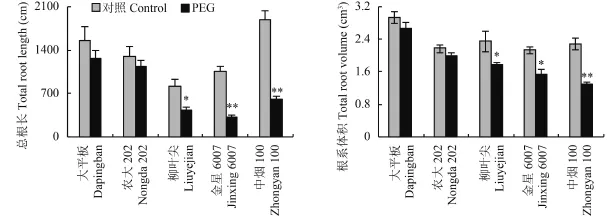
圖2 水分脅迫對煙草總根長和根體積的影響Fig. 2 Tobacco total root elongation and root volume in response to water stress[注(Note):* 和 ** 分別表示處理間差異在 0.05 和 0.01 水平差異顯著Indicate the significant differences between different treatments at the 0.05 and 0.01 levels, respectively.]
2.3 光合特性
與對照處理相比,水分脅迫條件下 5 個烤煙品種的凈光合速率、氣孔導(dǎo)度和蒸騰速率均呈下降趨勢,但抗旱能力弱的烤煙品種下降幅度要高于抗旱能力強的品種 (圖 5)。水分脅迫條件下大平板和農(nóng)大202 的凈光合速率的降幅分別為 27% 和 24%,3 個抗旱能力弱的烤煙品種的凈光合速率的下降幅度高達 52%~64%,其中柳葉尖和中煙 100 的降幅與對照處理相比差異達極顯著水平。氣孔導(dǎo)度、蒸騰速率和凈光合速率的變化基本一致,而兩個處理下 5個煙草品種的胞間二氧化碳濃度均沒有顯著差異。
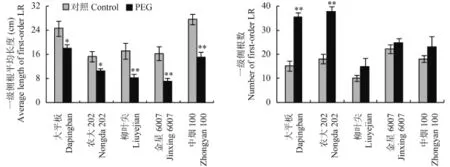
圖3 水分脅迫對煙草一級側(cè)根發(fā)生和伸長的影響Fig. 3 First-order lateral root elongation and formation in response to water stress[注(Note):LR—Lateral root. * 和 ** 分別表示處理間差異在 0.05 和 0.01 水平差異顯著Indicate the significant differences between different treatments at the 0.05 and 0.01 levels, respectively.]
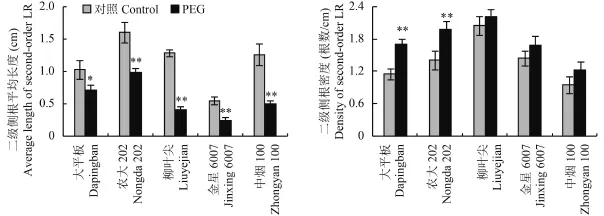
圖4 水分脅迫對煙草二級側(cè)根發(fā)生和伸長的影響Fig. 4 Second-order lateral root elongation and formation in response to water stress[注(Note):LR—Lateral root. * 和 ** 分別表示處理間在 0.05 和 0.01 水平差異顯著Indicate the significant differences between different treatments at the 0.05 and 0.01 levels, respectively.]
2.4 葉片溫度
水分脅迫對 5 個烤煙品種的葉片溫度影響不同(圖 6)。與對照處理相比,水分脅迫條件下大平板和農(nóng)大 202 的葉片溫度上升約 1℃,與對照相比差異不顯著;柳葉尖、金星 6007 和中煙 100 的葉片溫度上升 5℃~7℃,與對照相比差異達到極顯著水平。
2.5 水分脅迫條件下剪根處理對農(nóng)大 202 生長的影響
從圖 7 可看出,在水分脅迫條件下剪根處理對農(nóng)大 202 的地上部生長和光合特性影響顯著。與對照相比,剪根處理下農(nóng)大 202 地上部生物量的降幅高達 67%,降幅和水分脅迫處理下 3 個抗旱能力弱的地上部生物量的降幅相當。類似的現(xiàn)象也發(fā)生在凈光合速率、氣孔導(dǎo)度、蒸騰速率和葉片溫度上。這說明抗旱能力強的煙草品種的一級側(cè)根數(shù)目的增加是其抗旱能力強的重要生理原因。
3 討論
根系是植物吸水的主要器官,最先感知干旱脅迫并通過根系形態(tài)的適應(yīng)變化來減少水分散失和提高水分利用率[19-20]。水分虧缺的程度、作物品種及實驗條件不同,根系對水分虧缺響應(yīng)的程度不一樣[21-24]。在本試驗水分脅迫條件下,5 個烤煙品種的一級和二級側(cè)根伸長均受抑制,抗旱能力強的兩個品種側(cè)根長度的降幅顯著小于 3 個抗旱能力弱的烤煙品種;而且 2 個抗旱能力強的煙草品種的一級側(cè)根數(shù)目和二級側(cè)根密度顯著增加。根據(jù) 5 個煙草品種在干旱脅迫條件下地上部和根系生物量的變化,我們推測其根系對干旱的反應(yīng)對于煙草的抗旱性具有至關(guān)重要的作用,即抗旱能力強的大平板和農(nóng)大 202 由于一級和二級側(cè)根雖然長度有所下降,但側(cè)根數(shù)量顯著增加,最終反饋到地上部生長,因此在干旱脅迫條件下大平板和農(nóng)大 202 能維持和對照處理相當?shù)牡厣喜可锪浚欢购的芰θ醯娜齻€品種柳葉尖、金星 6007 和中煙 100,由于一級和二級側(cè)根長度的下降和總體數(shù)量的顯著降低,最終表現(xiàn)為生物量的顯著降低。
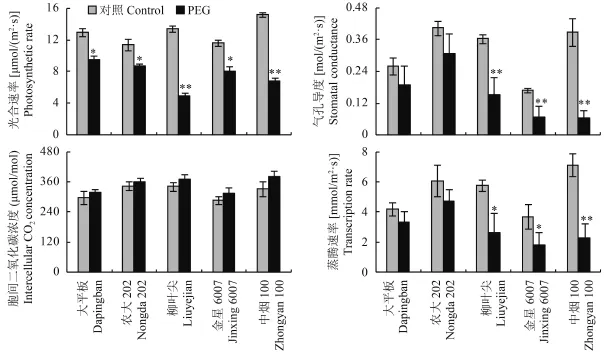
圖5 水分脅迫對煙草葉片光合特性的影響Fig. 5 Photosynthetic characteristics of tobacco leaves in response to water stress[注(Note):* 和 ** 分別表示處理間在 0.05 和 0.01 水平差異顯著Indicate the significant differences between different treatments at the 0.05 and 0.01 levels, respectively.]
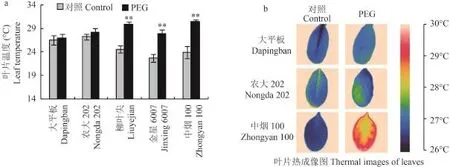
圖6 水分脅迫對煙草葉片溫度的影響Fig. 6 Leaf temperature in five tobacco plants in response to water stress[注(Note):** 表示處理間在 0.01 水平差異顯著Indicate the significant differences between different treatments at the 0.01 level, respectively.]
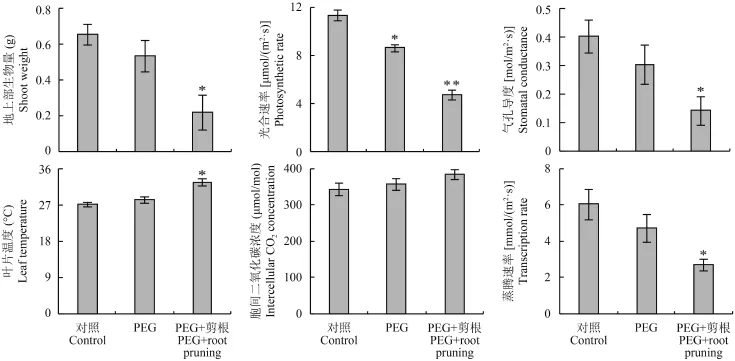
圖7 剪根對煙草品種農(nóng)大 202 生長的影響Fig. 7 Plant growth of Nongda 202 in response to root pruning[注(Note):* 和 ** 分別表示與對照處理相比在 0.05 和 0.01 水平差異顯著 Indicate the significant differences between control and PEG treatments, or control and PEG + root pruning treatments at the 0.05 and 0.01 levels, respectively.]
水分可以通過調(diào)控氣孔的開閉進而影響葉片的光合作用和蒸騰作用。干旱脅迫會導(dǎo)致葉片氣孔關(guān)閉,嚴重時會損傷葉肉細胞并降低 Rubisco 酶活性,導(dǎo)致植物的光合速率降低[25-26]。在本試驗條件下,雖然水分脅迫處理下 2 個抗旱煙草品種地上部生物量沒有下降,但葉片的凈光合速率卻顯著下降、氣孔導(dǎo)度和蒸騰速率也有降低的趨勢,但降幅顯著低于 3個不抗旱煙草品種,表明抗旱能力強的煙草品種光合作用的下降幅度要小于抗旱能力弱的煙草品種[27]。植物葉片受到太陽輻射后通過蒸騰作用散失水分來調(diào)節(jié)自身溫度[28-29]。干旱脅迫下由于氣孔關(guān)閉可導(dǎo)致葉片溫度升高[30-32]。本試驗條件下,由于水分脅迫條件下兩個抗旱能力強的煙草品種的氣孔導(dǎo)度變化不明顯,導(dǎo)致葉片溫度也沒有變化,而不抗旱品種氣孔導(dǎo)度的降幅超過 50%,最終導(dǎo)致葉片溫度上升 5℃~7℃,進而影響植物的生長。剪根實驗的結(jié)果表明,抗旱能力強的品種通過根系數(shù)量的增加,保證了地上部水分的供應(yīng),緩解了水分脅迫帶來的葉片溫度升高和光合能力下降等不利影響,而較強的光合作用效率反饋促進了根系側(cè)根數(shù)目的增加,進而減少了水分脅迫的傷害。抗旱能力差異顯著的不同煙草品種根系是如何感知并迅速產(chǎn)生信號物質(zhì)向上傳遞來影響光合作用的機制需要進一步研究。
4 結(jié)論
抗旱能力強的烤煙品種在水分脅迫條件下,可以增加一級和二級側(cè)根的數(shù)量,增加對水分的吸收,緩解葉片溫度升高和光合速率下降的趨勢。反過來,地上部高的光合能力能保證根系生長所需的碳水化合物進而促進根系的生長,最終表現(xiàn)為對干旱有較強的抵御能力。
[1]周翼衡, 朱小平, 王彥亭, 等. 煙草 生理與生物化學(xué)[M]. 合肥 : 中國科學(xué)技術(shù)大學(xué)出版社, 1996. Zhou Y H, Zhu X P, Wang Y T, et al. Tobacco physiology and biochemistry[M]. Hefei: Press of University of Science and Technology of China, 1996.
[2]姚玉璧, 王勁松, 尚軍林, 等. 基于相對濕潤度指數(shù)的 西南春季干旱10 年際演變特征[J]. 生態(tài)環(huán)境學(xué)報, 2014, 23(4): 547-554. Yao Y B, Wang J S, Shang J L, et al. Decadal variations of spring drought based on relative moisture index in southwest of China[J]. Ecology and Environmental Sciences, 2014, 23(4): 547-554.
[3]關(guān)軍鋒, 劉海龍, 李 廣敏. 干旱脅 迫 下小麥幼苗根 、葉多胺含量和多胺氧化酶活性的變化[J]. 植物生態(tài)學(xué)報, 2003, 27(5): 655-660. Guan J F, Liu H L, Li G M. Changes of polyamines content and polyamine oxidase activity of roots and leaves during drought stress in wheat seedlings[J]. Acta Phytoecologica Sinica, 2003, 27(5):655-660.
[4]榮智媛, 張曉海, 楊雙龍, 等. 抗氧化系統(tǒng)參與循環(huán)干 旱鍛煉提高煙草植株抗旱性的形成[J]. 植物生理學(xué)報, 2012, 48(7): 705-713. Rong Z Y, Zhang X H, Yang S L, et al. Involvement of antioxidant defense system in enhancement of drought resistance in tobacco (Nicotiana tabacum L.) plants through circular drought-hardening[J]. Plant Physiology Journal, 2012, 48(7): 705-713
[5]時向東, 方圓, 焦峰, 劉國順. 干旱脅迫 下烤煙葉片細胞壁 expansin積累動態(tài)研究[J]. 中國煙草學(xué)報, 2010, 16(1): 41-44. Shi X D, Fang Y, Jiao F, Liu G S. Research on the dynamics of leaf cytoderm expansin accumulation under drought stress in flue-cured tobacco[J]. Acta Tabacaria Sinica, 2010, 16(1): 41-44.
[6]Jia W S, Zhang J H. Stomatal movements and long-distance signaling in plants[J]. Plant Signaling and Behavior, 2008, 3: 772-777.
[7]Manavalan L P, Guttikonda S K, Tran L S P, Nguyen H T. Physiological and molecular approaches to improve drought resistance in soybean[J]. Plant Cell Physiology, 2009, 50: 1260-1276.
[8]Bengough A G, McKenzie B M, Hallett P D, Valentine T A. Root elongation, water stress, and mechanical impedance: a review oflimiting stresses and beneficial root tip traits[J]. Journal of Experimental Botany, 2011, 62: 59-68.
[9]Henry A, Cal A J, Batoto T C, et al. Root attributes affecting water uptake of rice (Oryza sativa) under drought[J]. Journal of Experimental Botany, 2012, 63: 4751-4763.
[10]余濼, 高明, 王子芳, 等. 土壤水分對烤煙生長、物質(zhì)分配和養(yǎng)分吸收的影響[J]. 植物營養(yǎng)與肥料學(xué)報, 2011, 17(4): 989-995. Yu S, Gao M, Wang Z F, et al. Effects of soil moisture content on growth, biomass partition and nutrient uptake of tobacco[J]. Plant Nutrition and Fertilizer Science, 2011, 17(4): 989-995.
[11]袁有波, 李繼新, 丁福章, 蘇賢坤. 不同干旱脅迫對煙草葉片保護酶活性的影響[J]. 中國煙草科學(xué), 2009, 30(5): 10-13. Yuan Y B, Li J X, Ding F Z, Su X K. Effects drought stress n activity of cell defense enzymes in flue-cured tobacco leaves[J]. Chinese Journal of Tobacco Science, 2009, 30(5): 10-13.
[12]馬文廣, 崔華威, 李永平, 等. 20 個煙草品種干旱脅迫下發(fā)芽和苗期生理特性及耐旱性評價[J]. 種子, 2012, 31(2): 25-30. Ma W G, Cui H W, Li Y P, et al. Physiological characteristics of 20 tobacco cultivars under drought stress and assessment of their drought tolerance at germination and seedling stages[J]. Seed, 2012, 31(2): 25-30.
[13]馬新蕾, 房燕, 王玉軍, 等. 十個烤煙品種的抗旱性鑒定[J]. 中國煙草學(xué)報, 2005, 11(5): 26-30. Ma X L, Fang Y, Wang Y J, et al. The evaluation of drought resistance of ten tobacco cultivars[J]. Acta Tabacaria Sinica, 2005, 11(5): 26-30.
[14]盧向陽, 彭麗莎, 唐湘如, 等. 早稻旱育秧形態(tài)、組織結(jié)構(gòu)和生理特性[J]. 作物學(xué)報, 1997, 23(3): 360-369. Lu X Y, Peng L S, Tang X R, et al. Study on the morphology, tissue structure and physiological characteristics of early rice (Oryza sativa L.) seedlings raised in dry nursey[J]. Acta Agronomica Sinica, 1997, 23(3): 360-369.
[15]丁艷鋒, 王強盛, 王紹華, 黃丕生. 水稻旱育秧苗與濕潤秧苗根系生理活性的比較研究[J]. 南京農(nóng)業(yè)大學(xué)學(xué)報, 2001, 24(3): 1-5. Ding Y F, Wang Q S, Wang S H, Huang P S. Comparison studies of roots physiology activity between rice dry seedbed seedlings and wet seedbed seedlings[J]. Journal of Nanjing Agricultural University, 2001, 24(3): 1-5.
[16]Kaufmann M R, Eckard A N. Evaluation of water stress control with polyethylene glycols by analysis of guttation[J]. Plant Physiology, 1971, 47: 453-456.
[17]尚曉潁, 劉化冰, 張小全, 等.干旱脅迫對不同烤煙品種根系生長和生理特性的影響[J]. 西北植物學(xué)報, 2010, 30(2): 357-361. Shang X Y, Liu H B, Zhang X Q, et al. Growth and physiological characteristics of roots in different flue-cured tobacco varieties under drought stress[J]. Acta Botanica Boreali-Occidentalia Sinica, 2010, 30(2): 357-361.
[18]任慶成, 陳秀華, 張生杰, 等.不同烤煙品種抗旱生理特征比較研究[J]. 西北植物學(xué)報, 2009, 29(10): 2019-2025. Ren Q C, Chen X H, Zhang S J, et al. Comparison of drought resistance characteristics of different flue-cured tobacco varieties[J]. Acta Botanica Boreali-Occidentalia Sinica, 2009, 29(10): 2019-2025.
[19]Sharp R E, Poroyko V, Hejlek L G, et al. Root growth maintenance during water deficits: physiology to functional genomics[J]. Journal of Experimental Botany, 2004, 55: 2343-2351.
[20]Moreno-Risueno M A, Van Norman J M, Moreno A, et al. Oscillating gene expression determines competence for periodic arabidopsis root branching[J]. Science, 2010, 329: 1306-1311.
[21]胡夢蕓, 張正斌, 徐萍, 等. 虧缺灌溉下小麥水分利用效率與光合產(chǎn)物積累運轉(zhuǎn)的相關(guān)研究[J]. 作物學(xué)報, 2007, 33(10): 1711-1719. Hu M Y, Zhang Z B, Xu P, et al. Relationship of water use efficiency with photoassimilate accumulation and transport in wheat under deficit irrigation[J]. Acta Agronomica Sinica, 2007, 33(10):1711-1719.
[22]馬富舉, 李丹丹, 蔡劍, 等. 干旱脅迫對小麥幼苗根系生長和葉片光合作用的影響[J]. 應(yīng)用生態(tài)學(xué)報, 2012, 23(3): 724-730. Ma F J, Li D D, Cai J, et al. Responses of wheat seedlings root growth and leaf photosynthesis to drought stress[J]. Chinese Journal of Applied Ecology, 2012, 23(3): 724-730.
[23]馬旭鳳, 于濤, 汪李宏, 等. 苗期水分虧缺對玉米根系發(fā)育及解剖結(jié)構(gòu)的影響[J]. 應(yīng)用生態(tài)學(xué)報, 2010, 21(7): 1731-1736. Ma X F, Yu T, Wang L H, et al. Effects of water deficit at seedling stage on maize root development and anatomical structure[J]. Chinese Journal of Applied Ecology, 2010, 21(7): 1731-1736.
[24]慕自新, 張歲岐, 郝文芳, 等. 玉米根系形態(tài)性狀和空間分布對水分利用效率的調(diào)控[J]. 生態(tài)學(xué)報, 2005, 25(11): 2895-2900. Mu Z X, Zhang S Q, Hao W F, et al. The effect of root morphological traits and spatial distribution on WUE in maize[J]. Acta Ecologica Sinica, 2005, 25(11): 2895-2900
[25]Bota J, Medrano H, Flexas J. Is photosynthesis limited by decreased Rubisco activity and RuBP content under progressive water stress?[J]. New Phytologist, 2004, 162: 671-681.
[26]Parry M A J, Andralojc P J, Khan S, et al. Rubisco activity: effects of drought stress[J]. Annals of Botany, 2002, 89: 833-839.
[27]胡瑋, 康俊, 劉陽, 等. 干旱脅迫對不同煙草品種光合生理特性的影響[J]. 中國煙草科學(xué), 2013, 34(2): 69-73. Hu W, Kang J, Liu Y, et al. The effect of drought stress on photosynthetic physiological characteristics of the different tobacco varieties[J]. Chinese Journal of Tobacco Science, 2013, 34(2): 69-73.
[28]Garrity D P, O’Toole J C. Selection for reproductive stage drought avoidance in rice, using infrared thermometry[J]. Agronomy Journal, 1995, 87(4): 773-779.
[29]Shahenshah, Isoda A. Effects of water stress on leaf temperature and chlorophyll fluorescence parameters in cotton and peanut[J]. Plant Production Science, 2010, 13(3): 269-278.
[30]呂樹作, 謝惠民, 張潔, 等. 不同冬小麥品種氣冠溫差與抗旱節(jié)水性的關(guān)系研究[J]. 麥類作物學(xué)報, 2007, 27(3): 533-538. Lü S Z, Xie H M, Zhang J, et al. Study on the relation of CTD and drought resistance water-saving in different winter wheats[J]. Journal of Triticeae Crops, 2007, 27(3): 533-538.
[31]黃春燕, 王登偉, 肖莉娟, 王雅芳. 不同水分條件下棉花光譜數(shù)據(jù)對冠層葉片溫度的響應(yīng)特征[J]. 棉花學(xué)報, 2014, 26(3): 244-251. Huang C Y, Wang D W, Xiao L J, Wang Y F. The responsive characteristics between cotton canopy leaves temperature from infrared thermography and hyperspectral data under different water conditions[J]. Cotton Science, 2014, 26(3): 244-251.
[32]Greer D H, Weedon M M. Modelling photosynthetic responses to temperature of grapevine (Vitis vinifera cv. Semillon) leaves on vines grown in a hot climate[J]. Plant Cell and Environment, 2012, 35:1050-1064.
Role of increasing first- and second-order lateral roots of tobacco for drought tolerance
ZHAO Yang1, WANG Shu-sheng1, ZHANG Ya-li2, XUE Ren2, WANG Cheng-dong1*
( 1 Institute of Tobacco Research of Chinese Academy of Agricultural Sciences,Qingdao, Shandong 266100, China; 2 College of Resources and Environmental Sciences,Nanjing Agricultural University, Nanjing 210095, China )
【Objectives】Drought stress is one of key environmental factors of limiting the growth and productivity of plants. However, which physiological processes to make plant more tolerant to drought stress remains unclear. The purpose of this study was to investigate the special physiological responses of droughttolerant tobacco (Nicotiana tabacum L.) adapted to drought stress.【Methods】A hydroponic culture experiment was conducted to analyze effect of drought stress (mimicking moderate drought-stress condition by application of 2.5% PEG) on five tobacco cultivars with contrasting responses to drought stress in this study. We investigated plant growth, including dry weight of shoot and root, formation and elongation of lateral roots in five tobacco cultivars. Meanwhile, we analyzed net photosynthetic rate and gas exchange parameters through Li-Cor 6400 photosynthetic analysis system and leaf temperature through an infrared camera (SC 620, FLIR Systems, Inc., USA) with a spectral sensitivity ranging from 7.5 mm to 13 mm and a spatial resolution of 0.65 mrad.【Results】Significant differences were observed in plant growth among five tobacco cultivars under drought stresscompared with the control treatment. Compared with the control treatment, two tobacco cultivars (Dapingban and Nongda 202) kept the similar shoot and root dry weight as those under drought stress. However, three other tobacco cultivars showed the drought-inhibited shoot and root growth with the decrement ranging from 66% to 83%. Accordingly, the two tobacco cultivars were considered as drought-tolerant tobacco cultivars and the three others as drought-susceptible cultivars. Further result showed that although drought stress decreased the elongation of first- and second-order lateral roots in five tobacco cultivars, formation of first- and second-order lateral roots was increased by drought stress only in two drought-tolerant tobacco cultivars. Interestingly, although significant decrement of shoot biomass induced by drought stress wasn’t observed in two droughttolerant tobacco cultivars, the net photosynthetic rate was decreased by drought stress relative to control treatment. The decrement of net photosynthetic rates of two drought-tolerant tobacco cultivars was less than three drought-susceptible tobacco cultivars. Furthermore, compared with the control treatment, the stomatal conductance and transpiration rate were decreased only in three drought-susceptible cultivars. After number of first-order lateral roots under the PEG treatment was trimmed to the similar level as that under the control treatment during the experimental stage, surprisingly, the shoot dry weight, net photosynthetic rate, stomatal conductance and transpiration rate of nongda202 were decreased to comparable extents as comparison with those in three drought-susceptible tobacco cultivars under the PEG treatment. This suggested that increment of lateral root formation was the main reason for two drought-tolerant tobacco cultivars to keep plant growth under drought stress relative to the control treatment.【Conclusions】The results indicated that compared with the control treatment, the formation of first- and second -order lateral roots was induced by moderate drought stress in drought-tolerant tobacco cultivars, and it resulted in higher photosynthetic characteristics in droughttolerant than in drought-susceptible tobacco cultivars under drought stress.
tobacco; drought stress; formation of lateral root; photosynthetic characteristics
2016-03-24 接受日期:2016-09-18
中國農(nóng)業(yè)科學(xué)院基本科研業(yè)務(wù)費預(yù)算增量項目資助。
趙陽(1994—),女,山東曹縣人,碩士研究生,主要從事作物栽培學(xué)與耕作學(xué)研究。E-mail:zhaoyang940201@163.com
* 通信作者 E-mail:wangchengdong198@163.com
猜你喜歡
奧秘(創(chuàng)新大賽)(2023年3期)2023-05-06 01:48:20
發(fā)明與創(chuàng)新(2022年30期)2022-10-03 08:40:56
動漫星空(興趣百科)(2020年12期)2020-12-12 05:31:40
人大建設(shè)(2018年6期)2018-08-16 07:23:10
文理導(dǎo)航·科普童話(2017年5期)2018-02-10 19:42:14
無人機(2017年10期)2017-07-06 03:04:36
浙江中西醫(yī)結(jié)合雜志(2017年2期)2017-01-12 18:23:59
當代化工研究(2016年9期)2016-03-20 16:22:08
小星星·閱讀100分(低年級)(2015年10期)2015-10-22 08:30:04
聲屏世界(2014年6期)2014-02-28 15:18:09
