輪作油葵對馬鈴薯生長發育及抗性生理指標的影響①
2017-04-24 09:36:43徐雪風李朝周張俊蓮
土壤 2017年1期
徐雪風,李朝周,張俊蓮
(1 甘肅省作物遺傳改良和種質創新重點實驗室,蘭州 730070;2 甘肅農業大學生命科學技術學院,蘭州 730070;3 甘肅農業大學園藝學院,蘭州 730070)
輪作油葵對馬鈴薯生長發育及抗性生理指標的影響①
徐雪風1,2,李朝周1,2,張俊蓮1,3*
(1 甘肅省作物遺傳改良和種質創新重點實驗室,蘭州 730070;2 甘肅農業大學生命科學技術學院,蘭州 730070;3 甘肅農業大學園藝學院,蘭州 730070)
為了探究輪作油葵對連作馬鈴薯生長發育及抗性生理的影響,選取馬鈴薯連作4年、連作4年后輪作油葵1年、連作6年、連作6年后輪作油葵1年的同一塊試驗田,以該試驗田前兩年分別種植藜麥、玉米的地塊為對照,測定土壤理化性質、土壤酶活性及土壤微生物數量變化,測評土壤環境。再在該試驗田種植馬鈴薯,對其幼苗光合作用、抗氧化酶活性以及馬鈴薯生長發育指標進行測定。結果表明,隨連作年限增加,馬鈴薯根際土壤pH總體升高,偏堿性,有機質和有效磷含量逐漸減少,堿解氮含量上升;土壤酶活性與連作年限呈負相關;隨連作年限增加,土壤中細菌數量、放線菌數量和細菌與真菌比(B/F)呈下降趨勢,真菌數量呈增加趨勢;土壤環境的惡化導致馬鈴薯植株生長量減少,葉綠素相對含量降低,葉片光合速率下降,超氧化物酶(SOD)活性下降,O2–.產生速率加快,丙二醛(MDA)含量上升。輪作油葵明顯降低了土壤 pH,提高了土壤有機質、有效磷和堿解氮含量,增加了土壤酶活性、細菌數量和B/F值,降低了真菌數量,改善了根際土壤微環境,對植株生長發育起到促進作用;增加了馬鈴薯葉片相對葉綠素含量、光合速率,SOD活性增強,而O2–.水平和MDA含量下降,可見輪作油葵減輕了馬鈴薯葉片膜脂過氧化作用和自由基傷害,促進了馬鈴薯生長發育,且整體效果以連作4年后輪作油葵較好。
馬鈴薯;連作;輪作;土壤環境
馬鈴薯 (Solanum tuberosum L.)是世界范圍內除水稻、小麥、玉米以外的第四大糧食作物,廣為種植[1–2]。農耕面積的有限和馬鈴薯集約化經營,使得馬鈴薯連作現象日益嚴重。長期連作導致土壤養分比例失調,土壤生物學環境惡化,根系分泌物的自毒作用加強,從而導致馬鈴薯產量及品質的下降[3–4]。谷巖等[5]對大豆連作研究表明,連作導致大豆土壤酶活性降低,根系分泌物增加,從而導致連作障礙。孟品品等[6]研究指出,連作馬鈴薯根際土壤中土傳病害病原菌數量增加,真菌種群成為優勢種群,導致根際生物學環境惡化,作用到根系,根系活力下降,塊莖產量及品質營養下降。呂毅等[7]研究發現,輪作能夠改善土壤環境,提高土壤微生物多樣性,減輕病害的發生和自毒作用。曹莉等[8]研究指出輪作豆科牧草可以改善連作馬鈴薯土壤環境,降低了土壤電導率,增加土壤脲酶、堿性磷酸酶及過氧化氫酶活性,對后茬馬鈴薯產量有明顯的影響。Mohr等[9]認為,在馬鈴薯生產體系中,輪作影響著植物和土壤的健康和生產力,與連作相比,輪作降低了土傳病害的發生率,增加了微生物種群及多樣性,從而減少畸形塊莖的比例,對馬鈴薯塊莖品質及產量產生影響。
目前關于輪作對連作馬鈴薯生長發育及抗性生理的影響及機制研究鮮有報道,本試驗針對甘肅省景泰縣馬鈴薯大田生產現狀,將油葵作為輪作作物應用到馬鈴薯種植中,通過測定馬鈴薯根際土壤理化指標及生物指標、馬鈴薯生長發育指標、光合作用及抗氧化酶活性指標,探究輪作油葵對連作馬鈴薯生長發育及抗性生理的影響及機理,以期為預防和減輕馬鈴薯連作障礙提供理論依據,并進一步優化馬鈴薯田間管理技術。
1 材料與方法
1.1 試驗材料
馬鈴薯(Solanum tuberosum L.)品種“大西洋”。
1.2 試驗設計
試驗于甘肅省白銀市景泰縣條山集團馬鈴薯種植基地進行,一年僅種植一茬作物。選取已有馬鈴薯連作0年(之前兩年分別種植藜麥、玉米)、連作4年、連作4年后輪作油葵1年,連作6年及連作6年后輪作油葵1年的同一塊試驗田,種植馬鈴薯,分別記作“對照”、“連作4年”、“連作4年+油葵”、“連作6年”和“連作6年+油葵”。種植前對上述5種連作田的土壤理化性質和土壤酶活性進行測定,采用5點法取樣,采集0 ~ 30 cm根區耕層土,混勻后用于測定土壤理化性質、土壤酶活性和土壤微生物數量。參照胡慧蓉和田昆[10]的方法,土壤有機質采用 K2Cr2O7容量法測定;土壤堿解氮采用堿解擴散法測定;土壤速效磷采用NaHCO3法測定;土壤pH采用酸度計法測定。測定結果見表1。

表1 輪作油葵對連作馬鈴薯土壤理化性質的影響Table 1 Effects of oil-sunflower rotation on soil physical and chemical properties under continuous cropping potato
1.3 測定指標與方法
1.3.1 土壤酶活性分析 脲酶活性采用苯酚鈉次氯酸鈉比色法測定,結果以 24 h后每克土壤(干重)中NH4+-N的毫克數表示[11];過氧化氫酶活性用高錳酸鉀滴定法測定,其活性以每克土所消耗的0.1 mol/L KMnO4溶液的毫升數表示[11];蔗糖酶的活性用3, 5-二硝基水楊酸比色法測定,以24 h后每克土葡萄糖毫克數表示[11];堿性磷酸酶活性采用磷酸苯二鈉法測定,結果以24 h后每克土壤中釋放出酚的毫克數表示[11]。
1.3.2 土壤微生物測定 采用牛肉膏蛋白胨培養基培養細菌,改良高氏1號培養基分離放線菌(每300 ml培養基加3% 的重鉻酸鉀1 ml,以抑制細菌和霉菌的生長),真菌采用改良馬丁氏培養基(每1 000 ml培養基中加1% 孟加拉紅水溶液3.3 ml、1% 鏈霉素3 ml)。每個連作年限土壤樣品稱取10 g,加入裝有90 ml無菌水的三角瓶中,振蕩30 min,即為10–1濃度的土壤樣液,靜置1 min,再依次稀釋(細菌稀釋至10–5,放線菌稀釋至 10–4,真菌稀釋至 10–3)。每個處理重復3次,平板涂布法接種,細菌放置于28℃ 恒溫箱中培養2 d,放線菌置于30℃ 恒溫箱中7 d,真菌置于25℃ 恒溫箱中5 d,最后計算每克干土中的微生物數量:
1 g干土中的菌數[12]=(每個培養基的菌落平均數′稀釋倍數)/干土重
1.3.3 生長發育指標測定 收獲前一周進行生長發育指標的測定,每個指標重復3次。株高:以植株主莖最高部位距土面的高度為準,用卷尺測量。莖粗:以植株與土面交界處地上莖直徑代表,用游標卡尺測量;匍匐莖數:收獲后目測;根長:取5條最長根,取平均值。
1.3.4 抗性生理指標測定 取馬鈴薯幼苗側莖部位第二、三葉進行測定,每個指標重復3次。相對葉綠素含量:SPAD-502葉綠素儀測定。凈光合速率、胞間 CO2濃度、氣孔導度:采用美國便攜式LI-6400光合儀,在上午10:00—11:00測定;丙二醛(Malondialdehyde,MDA)含量測定參照鄒琦[13];超氧化物歧化酶(Superoxide dismutase,SOD)活性測定參照 Spychalla 和 Desborough[14],以抑制氮藍四唑光化還原一半的酶量為一個酶活力單位;超氧陰離子(Superocide anion rate,O2–.) 產生速率測定采用羥胺氧化法[15]。
1.3.5 數據分析 應用SPSS17.0軟件對數據進行統計分析,并用LSD法進行顯著性檢驗。采用Excel 2003對數據進行處理、繪圖。
2 結果與分析
2.1 輪作油葵對連作馬鈴薯土壤酶活性的影響
圖1表明,隨著連作年限的增加,馬鈴薯土壤中堿性磷酸酶、脲酶、蔗糖酶及過氧化氫酶活性顯著減少,輪作油葵均提高了這4種土壤酶活性,但上升幅度有差別。連作4年+油葵處理對堿性磷酸酶和蔗糖酶活性的促進作用最大,分別比連作 4年提高了46.41% 和31.11%(圖1A,C),連作6年+油葵處理對脲酶和過氧化物酶活性的促進作用最大,分別比連作6年提高了42.93% 和40.72%(圖1B,C)。

圖1 輪作油葵對連作馬鈴薯土壤酶活性的影響Fig. 1 Effects of oil-sunflower rotation on activities of soil enzymes under continuous cropping potato
2.2 輪作油葵對連作馬鈴薯根際土壤微生物數量的影響
由表2可知,馬鈴薯土壤根際細菌數量和放線菌數量隨連作年限的延長呈下降趨勢。連作6年時分別比對照下降了 73.89%、42.78%,差異達到極顯著。輪作油葵后,均顯著增加(表 2)。細菌和放線菌數量在連作 4年+油葵處理中分別比連作 4年提高了72.04%、58.50%,連作6年+油葵處理的細菌數量和放線菌數量分別比連作6年提高了64.15%、19.42%。真菌數量則表現為隨連作年限增加呈上升趨勢。輪作油葵降低了土壤根際真菌數量,對于連作4年馬鈴薯的土壤真菌數量下降幅度(62.26%)顯著大于連作6年馬鈴薯的土壤真菌數量降幅(15.66%)。細菌與真菌的比值可以表征土壤肥力大小,隨著連作年限的增加,B/F值呈下降趨勢,連作4年B/F值是對照的10.66%,連作6年B/F值是對照的3.72%,由此看來連作馬鈴薯根系分泌物抑制細菌繁殖,改變了微生物群落結構,削減土壤肥力。輪作油葵相對提高了B/F值,尤其在連作4年后輪作油葵B/F值是連作4年的5.38倍(P<0.05)。

表2 輪作油葵對連作馬鈴薯土壤微生物數量的影響Table 2 Effects of oil-sunflower rotation on the quantities of soil microorganism under continuous cropping potato
2.3 輪作油葵對連作馬鈴薯幼苗葉片光合生理指標的影響
由表3可知,隨連作年限增加,馬鈴薯幼苗葉片葉綠素相對含量、凈光合速率、胞間 CO2濃度、氣孔導度逐漸減小,而輪作油葵后則顯著提高。連作4年+油葵處理的馬鈴薯葉片葉綠素相對含量、凈光合速率、胞間CO2濃度分別比連作4年增加了11.73%、9.20%、8.00%,且均高于對照,氣孔導度增加了7.67%;連作 6年+油葵處理則分別提高了 4.71%、8.12%、5.05%、11.53%,增加幅度較連作4年+油葵處理小(氣孔導度除外)。由此可以得出,連作4年后輪作油葵對于增強連作馬鈴薯幼苗光合作用的效果顯著。

表3 輪作油葵對連作馬鈴薯幼苗葉片光合生理指標影響Table 3 Effects of oil-sunflower rotation on photosynthesis indexes of potato seedling leaves under continuous cropping potato
2.4 輪作油葵對連作馬鈴薯幼苗葉片丙二醛含量、O2–.產生速率、SOD活性的影響
一般情況下,當植物遭受環境脅迫時,細胞內迅速積累活性氧,引起膜脂過氧化產物丙二醛(MDA)含量上升。由圖2可知,馬鈴薯葉片MDA含量的變化隨連作年限增加而積累,植物受到過氧化傷害;輪作油葵降低了MDA含量,減輕了連作引起的馬鈴薯傷害。對于4年和6年連作,輪作油葵比不輪作處理分別降低了34.49% 和22.77%,下降幅度連作4年后輪作油葵明顯要高,馬鈴薯受傷害較輕(圖 2A)。O2–.隨連作年限的延長逐漸升高,輪作油葵后其產生速率明顯下降,連作4年和連作6年輪作油葵分別比不輪作處理下降了18.68%、11.35%(圖2B),說明輪作油葵可以降低連作馬鈴薯O2–.產生速率。超氧化物酶(SOD)活性隨連作年限延長呈先增后減趨勢,連作6年時的SOD活性比對照下降了27.38%。輪作油葵提高了SOD活性,且連作4年+油葵處理的SOD活性顯著高于連作6年+油葵處理(P<0.05)(圖2C)。具體效果視連作年限而異。
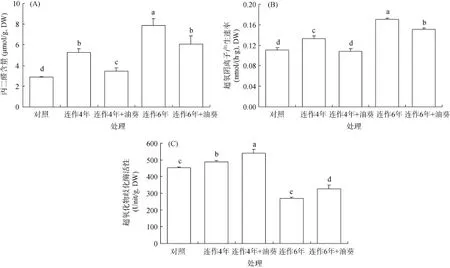
圖2 輪作油葵對連作馬鈴薯幼苗葉片丙二醛含量、超氧陰離子產生速率和、超氧化物酶活性的影響Fig. 2 Effects of oil-sunflower rotation on MDA content, SOD activity and production rate of O2–.of potato seedling leaves under continuous cropping
2.5 輪作油葵對連作馬鈴薯生長發育的影響
由表4可知,連作馬鈴薯株高、莖粗、根長、地上部干重、單株薯重均比對照低,且與連作年限呈負相關。連作年限對馬鈴薯株高、莖粗和根長的抑制效果明顯,且連作6年處理分別比對照下降了41.62%、43.18%、19.33%。輪作油葵相對提高了馬鈴薯的株高,使得莖粗增加,促進根的伸長生長,連作4年+油葵處理下分別較連作 4年處理上升了 8.97%、20.43%、37.14%,連作6年+油葵處理下分別較連作6年處理升高了3.16%、13.33%、36.10%,且根長在輪作后要比對照值高。連作處理下的地上部干重和單株產量顯著低于對照,連作 4年處理比對照下降了55.79%、41.75%,差異顯著;連作 6年處理下進一步降低,但與連作4年差異不顯著,分別比對照降低了 61.30%、51.14%。輪作油葵提高了地上部干重和單株產量,連作4年+油葵處理下分別較連作4年處理上升了37.52%、33.99%,連作6年+油葵處理下分別較連作6年處理升高了35.75%、10.06%。以上數據表明,連作4年輪作油葵對于提高連作馬鈴薯植株株高、莖粗、根長、地上部干重和單株產量較連作6年+油葵處理相對效果好,促進了連作馬鈴薯的生長發育。

表4 輪作油葵對連作馬鈴薯生長發育的影響Table 4 Effects of oil-sunflower rotation on growth and development of potato under continuous cropping potato
3 討論
3.1 輪作油葵對連作馬鈴薯根際土壤環境的影響
作物生長的首要條件是擁有良好的土壤環境,土壤酶在土壤物質循環和能量流動過程中至關重要,土壤酶活性的強弱,直接反映了土壤肥力和生產力大小[16–17]。本研究發現,與對照相比,隨著連作年限的增加,馬鈴薯土壤中的堿性磷酸酶、脲酶、蔗糖酶及過氧化氫酶活性降低(圖1),磷酸酶活性的下降抑制了土壤中有機磷化物的分解,使得土壤中有效利用的磷素含量下降,有效磷含量也隨之下降(表1),土壤肥力下降,這與康亞龍等[18]的研究結果一致。通過輪作油葵,土壤酶活性均有所提高,在連作 4年馬鈴薯田輪作油葵對土壤堿性磷酸酶活性和蔗糖酶活性的促進作用顯著,土壤中可利用的磷素和營養物質增多,土壤狀況好轉,這與Marchinkeviciene等[19]研究結果相似,說明輪作加快了土壤物質代謝,使得土壤中的有機物質循環加快,根系分泌物的自毒作用下降,土壤酶活性增加。
土壤微生物是土壤生態系統的重要組成成分之一,連作導致土壤微生物數量變化,有益微生物減少,病原菌微生物增加[20]。本試驗結果表明,長期連作使得馬鈴薯根際土壤細菌、放線菌數量及 B/F值下降,真菌數量增加,向真菌型土壤過渡,這與顧美英等[21]研究結果相似,不利于馬鈴薯生長。輪作改變了土壤微生物環境,促進根系吸收能力,從而加快微生物生長和繁殖[22]。本試驗中輪作油葵使得有益微生物細菌數量、放線菌數量增加,真菌數量相對降低,土壤轉向細菌型(表2),土壤肥力增加,且連作4年輪作油葵對土壤微生物環境的改善效果較好。
有機質是土壤的重要組成成分,可以反映土壤養分狀況[23]。本試驗結果表明,隨著連作年限(0,4,6年)的增加,馬鈴薯土壤堿解氮含量上升,pH介于7.5 ~ 8.5,有機質和有效磷含量減少,土壤養分比例失調,土壤質量下降。輪作油葵在一定程度上增加了土壤有機質、有效磷和堿解氮含量,這與王濤等[24]對黃瓜連作的研究一致。相反,土壤pH相對降低,其中輪作油葵對連作 4年的馬鈴薯土壤理化性質的改良效果更有效(表1)。徐成龍[25]研究也認為,輪作苜蓿能提高土壤有機質、速效氮、有效磷含量,從而提高常規農田土壤質量和作物產量。結合馬鈴薯生長發育指標(表 4),輪作油葵明顯提高了馬鈴薯株高、莖粗、根長、地上部干重和單株產量,說明輪作油葵對土壤環境的改變為馬鈴薯生長發育提供有利的生長條件。整體上馬鈴薯在連作4年后輪作油葵處理的生長明顯較連作6年后輪作油葵處理要好。
3.2 輪作油葵對連作馬鈴薯幼苗光合生理指標的影響
光合作用是植物進行物質循環和能量流動的基礎,葉綠素則是植物光合作用過程中的重要色素,與光合效率呈正相關,可以作為反映植物逆境生理指標之一[26–27]。本研究表明,連作使得馬鈴薯幼苗葉片葉綠素相對含量下降,凈光合速率、胞間 CO2濃度以及氣孔導度下降,植株光合作用減弱(表 3),影響馬鈴薯生長,與許楠等[28]的研究結果相一致。輪作油葵能顯著提高連作馬鈴薯葉片葉綠素相對含量、凈光合速率,增強了光合速率,進一步促進馬鈴薯植株正常生長。
3.3 輪作油葵對連作馬鈴薯幼苗抗氧化酶活性的影響
植物生命過程中都會有自由基的產生,逆境條件下體內的自由基積累,產生自由基鏈式反應,引起膜脂過氧化,對細胞膜產生損傷,而細胞膜脂過氧化作用的強弱可以用丙二醛(MDA)含量的高低來反映,超氧化物酶(SOD)是細胞內清除超氧自由基的重要酶[29]。研究結果顯示,隨著連作年限的增加,馬鈴薯幼苗葉片MDA含量增加,SOD活性下降,使得 O2–.積累,膜脂過氧化作用加快,連作 6年時表現最為明顯,馬鈴薯遭受脅迫明顯。輪作油葵明顯抑制了SOD活性的下降,使得O2–.水平下降,降低了MDA含量,從而減輕膜脂過氧化作用對細胞膜的損傷,提高植株的抗氧化能力(圖 2),促進馬鈴薯正常生長,這與呂毅等[7]的研究結果相似。
4 結論
馬鈴薯連作(4,6年)后有機質含量下降,氮肥、磷肥比例失調,土壤理化性質發生改變,土壤酶活性的降低以及土壤微生物組成變化,從根源上抑制了馬鈴薯生長,使得葉片光合作用減弱,丙二醛積累,超氧陰離子產生速率增加,導致膜脂過氧化加重。輪作油葵在一定程度上改善了土壤環境,促進了馬鈴薯生長發育,增強了連作馬鈴薯的光合作用和抗氧化能力,減輕脂質過氧化作用。連作4年后輪作油葵對于改善土壤環境、增強葉片光合作用和抗氧化能力、降低丙二醛含量較連作6年后輪作油葵效果更明顯,更有利于減輕馬鈴薯連作障礙。可見,馬鈴薯連作障礙不僅表現在馬鈴薯生長發育的下降,也表現在土壤生物和非生物環境因子的惡化;輪作油葵則從整體上逆轉了連作導致的植株和土壤兩方面的不利變化,而且這種作用以連作4年時輪作油葵效果最為顯著。
[1] 李瑞琴, 劉星, 邱慧珍, 等. 連作馬鈴薯根際干腐病優勢病原菌熒光定量 PCR快速檢測及在根際的動態變化[J]. 草業學報, 2013, 22(6): 239–248
[2] 回振龍, 李朝周, 史文煊, 等. 黃腐酸改善連作馬鈴薯生長發育及抗性生理的研究[J]. 草業學報, 2013, 22(4): 130–136
[3] 郭紅偉, 郭世榮, 劉來, 等. 辣椒連作對土壤理化性狀、植株生理抗性及離子吸收的影響[J]. 土壤, 2012, 44(6): 1041–1047
[4] 張仕祥, 過偉民, 李輝信, 等. 煙草連作障礙研究進展[J].土壤, 2015, 47(5): 823–829
[5] 谷巖, 邱強, 王振民, 等. 連作大豆根際微生物群落結構及其土壤酶活性[J]. 中國農業科學, 2012, 45(19): 3955–3964
[6] 孟品品, 劉星, 邱慧珍, 等. 連作馬鈴薯根基土壤真菌種群結構及其生物效應[J]. 應用生態學報, 2012, 23(11): 3079–3086
[7] 呂毅, 宋富海, 李園園, 等. 輪作不同作物對蘋果園連作土壤環境及平邑甜茶幼苗生理指標的影響[J]. 中國農業科學, 2014, 47(14): 2830–2839
[8] 曹莉, 秦舒浩, 張俊蓮, 等. 輪作豆科牧草對連作馬鈴薯田土壤微生物群落及酶活性的影響[J]. 草業學報, 2013, 22(3): 139–145
[9] Mohr R M, Derksen D A, Irvine R B, et al. Effects of rotation on crop yield and quality in an irrigated potato system[J]. Am. J. Pot Res., 2011, 88: 346–359
[10] 胡慧蓉, 田昆. 土壤學實驗指導教程[M]. 北京: 中國林業出版社, 2012
[11] 吳金水, 林啟美, 黃巧云, 等. 土壤微生物量測定方法及其應用[M]. 北京: 氣象出版社, 2006
[12] 李坤. 葡萄連作障礙機理及調控途徑的研究[D]. 沈陽:沈陽農業大學, 2010
[13] 鄒琦. 植物生理生化實驗指導[M]. 北京: 中國農業出版社, 2003
[14] Spychalla J P, Desborough S L. Superoxide dismutase, catalase and alpha tocopherol content of stored potato tubers[J]. Plant Physiology, 1990, 94: 1214–1218
[15] 王愛國, 羅廣華. 植物的超氧物自由基與羥胺反應的定量關系[J]. 植物生理學通訊, 1990(6): 55–57
[16] 沈芳芳, 袁穎紅, 樊后保, 等. 氮沉降對杉木人工林土壤有機碳礦化和土壤酶活性的影響[J]. 生態學報, 2012, 32(2): 517–527
[17] 徐雪風, 回振龍, 李自龍, 等. 馬鈴薯連作障礙與土壤環境因子變化相關研究[J]. 干旱地區農業研究, 2015, 33(4): 16–23
[18] 康亞龍, 景峰, 孫文慶, 等. 加工番茄連作對土壤理化性質及微生物數量的影響[J]. 土壤學報, 2016, 53(2): 533–542
[19] Marcinkeviciene A, Boguzas V, Balnyte S, et al. Influence of crop rotation, intermediate crops, and organic fertilizers on the soil enzymatic activity and humus content in organic farming systems[J]. Eurasian Soil Science, 2013, 46(2): 198–203
[20] 張云偉, 徐智, 湯利, 等. 不同有機肥對烤煙根際土壤微生物的影響[J]. 應用生態學報, 2013, 24(9): 2551–2556
[21] 顧美英, 徐萬里, 茆軍, 等. 新疆綠洲農田不同連作年限棉花根際土壤微生物群落多樣性[J]. 生態學報, 2012, 32(10): 3031–3040
[22] 虎德鈺, 毛桂蓮, 許興. 不同草田輪作方式對土壤微生物和土壤酶活性的影響[J]. 西北農業學報, 2014, 23(9): 106–113
[23] 高菊生, 曹衛東, 李冬初, 等. 長期雙季稻綠肥輪作對水稻產量及稻田土壤有機質的影響[J]. 生態學報, 2011, 31(16): 4542–4548
[24] 王濤, 喬衛花, 李玉奇, 等. 輪作和微生物菌肥對黃瓜連作土壤理化性狀及生物活性的影響[J]. 土壤通報, 2011, 42(3): 578–583
[25] 徐成龍. 苜蓿輪作農田和常規農田土壤質量與作物產量的比較研究[D]. 蘭州: 蘭州大學, 2014
[26] 李夏, 妙佳源, 高小麗, 等. 連作條件下谷子葉片衰老與活性氧代謝研究[J]. 中國農業大學學報, 2016, 21(4): 1–9
[27] 曾令杰, 林茂茲, 李振方, 等. 連作對太子參光合作用及藥用品質的影響[J]. 作物學報, 2012, 38(8): 1522–1528
[28] 許楠, 張會慧, 李鑫, 等. 連作對烤煙生長特性和光合能力的影響[J]. 草業學報, 2012, 29(9): 1435–1440
[29] 李朝周, 左麗萍, 李毅, 等. 兩個海拔分布下紅砂葉片對滲透脅迫的生理響應[J]. 草業學報, 2013, 22(1): 176–182
Effects of Oil-sunflower Rotation on Growth and Resistance Physiology Indexes of Potato
XU Xuefeng1,2, LI Chaozhou1,2, ZHANG Junlian1,3*
(1 Gansu Key Laboratory of Crop Genetics & Germplasm Enhancement, Lanzhou 730070, China; 2 College of Life Sciences and Technology, Gansu Agricultural University, Lanzhou 730070, China; 3 College of Hoticulture, Gansu Agricultural University, Lanzhou 730070, China)
Different farmlands including only cropping quinoa(CK), 4 years continuous cropping potato, 1 year oilsunflower rotation after 4 years continuous cropping potato, 6 years continuous cropping potato, 1 year oil-sunflower rotation after 6 years continuous cropping potato were selected, and which soil properties, soil enzyme activities, soil microorganism quantities and the growth and development indicators of potatoes were measured and compared to explore the effects of oil-sunflower rotation on the growth and resistance physiology of potato under different continuous cropping potato. The results showed that with the increase of continuous cropping year, soil pH of potato rhizosphere increased and tended to alkalinity in general, the contents of organic matter and available phosphorus decreased, alkali-hydrolyzable nitrogen content increased. Soil enzyme activities were negatively correlated with continuous cropping year and were lowest in the 6 years continuous cropping. With the increase of continuous cropping year, the quantities of soil bacteria and actinomycetes and the ratio of bacteria and fungi (B/F) decreased, but fungi quantity increased. The deterioration of potato rhizosphere soil environment resulted in the reduction in potato growth, chlorophyll content, leaf photosynthesis, SOD activity and the increase in production rate of O2–.and MDA content. In general, compared with non-rotation, oil-sunflower rotation significantly reduced soil pH, increased contents of organic matter, available phosphorus and alkali-hydrolyzable, improved soil enzyme activities, the quantity of bacteria and B/F, reduced the quantity of fungi, improved soil microenvironment, promoted potato growth, increased relative chlorophyll content, photosynthesis and SOD activity, decreased production rate of O2–.and MDA content, reduced membrane lipid peroxidation and free radical damage, enhanced antioxidant capacity. Generally, oil-sunflower rotation in 4 years continuous cropping was better, which well alleviated potato continuous cropping obstacles.
Potato; Continuous cropping; Rotation; Soil environment
S532
A
10.13758/j.cnki.tr.2017.01.013
國家科技支撐計劃項目(2012BAD06B03)和國家馬鈴薯產業技術體系項目(CARS-10-P18)資助。
* 通訊作者(zhangjunlian@163.com)
徐雪風(1991—),女,陜西富平人,碩士研究生,主要從事植物生態生理方面的研究。E-mail: 1075167312@qq.com
