
柔嫩艾美耳球蟲江蘇株SAG10基因的克隆及分析
2017-05-02 11:24:17仇保豐景瑾董蓉蓮宋鴻雁劉春朱順
江蘇農業科學 2017年3期
關鍵詞:分析
仇保豐 景瑾 董蓉蓮 宋鴻雁 劉春 朱順星 邵義祥 劉文斌 李建


摘要:根據GenBank中已發表的柔嫩艾美耳球蟲(Eimeria tenella) SAG10基因序列,利用計算機軟件設計1對引物,從E. tenella江蘇株的第2代裂殖子中成功克隆出SAG10基因。將該基因與GenBank中國內外不同E. tenella分離株的SAG10基因進行比對,發現E. tenella江蘇株與北京株的SAG10基因同源性最高,高達99%,其后依次為楊凌株、豪頓株。4種不同分離株SAG10基因的ORF之間有11個堿基不同,其中3個為無義突變,8個為有義突變。分析4種分離株SAG10基因推導蛋白質的親水性、抗原指數、表面可及性發現,國內外不同E. tenella分離株SAG10蛋白的抗原性總體比較接近且很強,但國內不同分離株之間的抗原性更加接近,并優于國外的豪頓株,這可能是由于氨基酸突變引起蛋白質的空間構象和親水性等發生變化而引起。將上述SAG10基因序列與復頂門原蟲的其他相關SAGs基因序列進行遺傳進化分析,發現Eimeria屬球蟲SAGs基因之間相對比較保守,且與犬新孢子蟲(Neospora caninum)、N. hughesi、剛地弓形蟲(Toxoplasma gondii)的SAG基因之間同源性較低,與已有觀點一致;但神經肉孢子蟲(Sarcocystis neurona)的2條SAG1基因序列卻與E. tenella的SAG1基因序列表現出更高的同源性,這與其他研究結論不符。
關鍵詞:柔嫩艾美耳球蟲;SAG10基因;克隆;分析
中圖分類號: Q785文獻標志碼: A
文章編號:1002-1302(2017)03-0024-04
雞球蟲基因克隆始于1987年,迄今已有240多個基因被成功克隆,雞球蟲的重組疫苗也得到了廣泛研究[1]。有分析認為,參與艾美耳球蟲宿主細胞入侵過程的相關分子是最具應用潛力的保護性抗原,如子孢子和裂殖子表面抗原(surface antigens,簡稱SAGs)以及微線、棒狀體、折光體等分泌型亞細胞器的分泌蛋白[1-2]。柔嫩艾美耳球蟲(Eimeria tenella)表面抗原SAG10(surface antigen 10)是一種C端具有疏水性的糖基化磷脂酰肌醇(glycosylphosphatidylinositol,簡稱GPI)錨定結構域的表面蛋白[3]。由于SAG10基因在E. tenella子孢子和第2代裂殖子階段均有表達,且SAGs被認為在啟動頂復門原蟲識別、黏附和侵襲宿主細胞、免疫調節、免疫逃避、宿主特異性等方面均發揮著重要作用,因此國外很多學者正加大對SAG10等表面抗原的研究力度[3-6]。韓德強等[7]、麥博等[2]分別對E. tenella北京株、楊凌株的SAG10基因進行了研究,但目前尚無對E. tenella江蘇株SAG10基因進行研究的報道。麥博等研究發現,SAG10在不同E. tenella株間存在抗原性差異,這與認為E. tenella不存在明顯株間抗原性差異的傳統觀點相對立[2]。本研究通過對E. tenella江蘇(Jiangsu)株的SAG10基因進行克隆,并將其與國內外學者在GenBank中公布的E. tenella豪頓(Houghton)株、北京(Beijing)株、楊凌(Yangling)株的SAG10基因進行比對和分析,為研究 E. tenella SAG10的生物學功能、篩選E. tenella保護性抗原等工作提供依據。
1材料與方法
1.1試驗材料
1.1.1球蟲卵囊
E. tenella分離自江蘇省某雞場,由南通市出入境檢驗檢疫局有害生物檢疫實驗室鑒定并保存。
1.1.2試驗雞
1日齡AA肉雞,購自江蘇省南通市某商品雞孵化場,飼養于嚴格消毒且無球蟲的籠舍中,飼料中不含抗球蟲藥。
1.1.3試劑
焦碳酸二乙酯(DEPC),購自Amresco公司;莫洛尼氏鼠白血病病毒(M-MLV)、Oligo(dT)15 Primer,購自Promega公司;pMD18-T克隆載體、Taq DNA聚合酶、dNTP、DL 2 000 Marker、限制性內切酶BamHⅠ、EcoRⅠ、DNA膠回收試劑盒,均購自寶生物工程(大連)有限公司。
1.2試驗方法
1.2.1引物設計與合成
根據Tabares等在GenBank中發表的E. tenella SAG10基因序列[3](登錄號:AJ586552),利用Primer Premier 5.0軟件設計1對特異性擴增引物,上游引物為5′-CA[ZZ(Z]GGATCC[ZZ)]ATGCTACAGCGGAAGCTAC-3′,下游引物為5′-TG[ZZ(Z]GAATTC[ZZ)]TAAAGTCATAATGCCGAAC-3′,擴增的目的片段長度為799 bp。為便于后續操作,分別在上、下游引物中設計了酶切位點BamHⅠ、EcoRⅠ(如下劃線部分所示)。引物由寶生物工程(大連)有限公司合成。
1.2.2cDNA模板的制備
將30羽1日齡AA肉雞飼養至14日齡,經口接種新鮮孢子化卵囊1×105個/羽,接種后 120 h 殺雞取盲腸。參考文獻[8]的方法分離和純化 E. tenella 第2代裂殖子,再采用一步法[9]提取總RNA并進行反轉錄,以反轉錄cDNA作為PCR模板。
1.2.3SAG10基因的克隆及鑒定
采用特異性引物進行SAG10基因的PCR擴增。PCR反應體系:10×Taq buffer 2.5 μL、MgCl2(25 mmol/L)1.0 μL、dNTP Mix(10 mmol/L each)0.5 μL、上游及下游引物(50 μmol/L)各0.5 μL、Taq DNA聚合酶(5 U/μL)0.5 μL、滅菌雙蒸水14.5 μL、cDNA模板5 μL。PCR反應條件:94 ℃ 5 min;94 ℃ 55 s,58 ℃ 55 s,72 ℃ 55 s,35個循環;72 ℃延伸15 min。循環完畢后,向擴增產物中加入5 μL上樣緩沖液,在1%瓊脂糖凝膠上電泳,并在紫外燈下觀察結果。回收SAG10基因目的條帶,連接pMD18-T載體并轉化DH5α,用BamHⅠ+EcoRⅠ雙酶切及PCR鑒定出陽性克隆,挑取3個克隆送至寶生物工程(大連)有限公司測序,將測序結果輸入“http://www.ncbi.nlm.nih.gov/BLAST/”進行網上比對。
1.2.4SAG10蛋白的特性分析
采用DNAStar 4.0軟件分別編輯GenBank中報道的E. tenella豪頓株、北京株、楊凌株、本研究克隆的江蘇(Jiangsu)株的SAG10基因序列,分析開放閱讀框(ORF)的堿基序列,并推導出編碼蛋白的氨基酸序列。采用MegAlign Clustal V軟件對SAG10基因ORF的堿基序列和推導氨基酸序列進行比對,分析E. tenella不同分離株之間SAG10基因堿基和推導氨基酸的突變情況,進一步采用DNAStar Protean軟件預測這些突變引起SAG10蛋白親水性結構(Hydrophilicity Plot)、抗原性指數(Antigenicindex)、表面可及性(Surface Probability Plot)的變化情況。
1.2.5SAG10基因遺傳進化分析
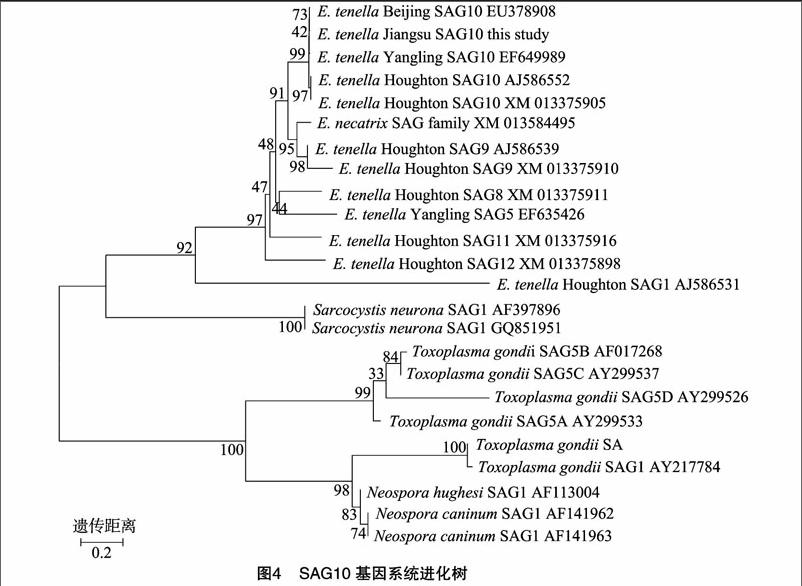
分別收集若干條GenBank中公布的E. tenella和毒害艾美耳球蟲、新孢子蟲、肉孢子蟲、弓形蟲等其他頂復門原蟲SAGs基因家族(SAGs gene family)序列,結合GenBank中已公布及本研究克隆的E. tenella SAG10基因序列,共同采用MEGA 6.06軟件構建遺傳進化發生樹,分析本研究克隆SAG10基因和其他SAG基因之間的遺傳進化關系。
2結果與分析
2.1SAG10基因的克隆
從E. tenella第2代裂殖子中提取總RNA并進行逆轉錄PCR(RT-PCR)擴增,產物經1%瓊脂糖凝膠電泳檢測,發現2個泳道中均有1條約799 bp的條帶(圖1),與預期大小相吻合。切膠并回收目的條帶后,連接pMD18-T載體,轉化大腸桿菌DH5α。
2.2SAG10基因的鑒定
提取pMD18-T-SAG10基因重組質粒,經雙酶切鑒定,可獲得與預期大小一致的目的條帶,進一步采用PCR方法進行鑒定,也能得到約799 bp的條帶(圖2),與理論值相吻合。從雙酶切和PCR鑒定的陽性克隆中挑取3個菌落進行DNA測序,然后將測序結果輸入GenBank進行Blast比對, 發現本研究克隆的E. tenella江蘇株SAG10基因與GenBank中公布的E. tenella北京株SAG10基因同源性最高,高達99%,其后依次為楊凌株、豪頓株,表明SAG10基因克隆成功。
2.3SAG10的蛋白特性分析
目前,GenBank中公布的E. tenella SAG10基因序列共有4條,其中同源性為100%的2條豪頓株SAG10基因序列由國外不同研究小組發表,北京株、楊凌株SAG10基因序列均由國內學者發表(表1)。這4條SAG10基因均只有完整的ORF部分,無兩端序列,全長786 bp,編碼261個氨基酸,蛋白分子量約為27.9 ku。本研究克隆的E. tenella江蘇(Jiangsu)株SAG10基因全長799 bp,其中不僅包含E. tenella SAG10基因完整ORF,同時還包含引物設計過程中人為添加的保護堿基和酶切位點。采用DNAStar 4.0軟件分析本研究克隆的SAG10基因的ORF,并推導出氨基酸序列,與GenBank中發表的4條序列共同采用MegAlign Clustal V軟件比對ORF內堿基序列和推導氨基酸序列。結果發現,4種不同E. tenella分離株SAG10基因之間共有11個位點存在差異(本研究中引物設計時人為引入的突變未計算在內,下同),除去第426、435、552位3個位點的堿基突變為無義突變外,其余8個堿基突變均為有義突變,引起了8個氨基酸突變(表1)。進一步采用DNAStar Protean軟件中的Kety-Doolittle、Jameson-Wolf、Emini算法,依次分析SAG10蛋白的親水性結構、抗原指數、表面可及性的變化情況。結果顯示,不同E. tenella分離株SAG10蛋白的N端和C端疏水性均較強,而中間段則有很多親水性區域。國內外不同E. tenella分離株之間,SAG10蛋白抗原性總體比較接近,且抗原性很強,但國內不同分離株之間的抗原性更加接近,它們與國外豪頓株的氨基酸序列在140~160位差異均較大,且國內分離株的抗原性優于國外豪頓株(圖3)。這可能是由于E. tenella國內分離株和國外豪頓株的SAG10氨基酸序列在140~160位變化頻率較高(表1),引起蛋白質的空間構象和親水性發生一定變化而導致。
160位氨基酸序列位于SAG10蛋白表面的可能性較大,一定程度上可解釋為何該區域氨基酸突變頻率較高會對國內外分離株之間抗原性差異產生影響。
2.4SAG10基因遺傳進化分析
本研究收集了GenBank中已發表的E. tenella 4條SAG10基因和1~2條不等的SAG1、SAG5、SAG8、SAG9、SAG11、SAG12基因序列。同時收集了毒害艾美耳球蟲(Eimeria necatrix)SAG基因家族序列和犬新孢子蟲(Neospora caninum)、洪氏新孢子蟲(Neospora hughesi)、神經肉孢子蟲(Sarcocystis neurona)的SAG1基因序列,以及剛地弓形蟲(Toxoplasma gondii)SAG1(別稱P30)和SAG5(可分為A、B、C、D共4種多肽)的基因序列,結合本研究克隆的E. tenella SAG10基因序列,共同采用MEGA 6.06軟件構建遺傳進化發生樹(圖4)。結果表明,E. tenella和E. necatrix的SAGs基因家族序列均分布在進化樹的同一較大分支上,表明艾美耳屬(Eimeria)球蟲的SAGs基因之間可能相對比較保守。N. caninum、N. hughesi、T. gondii的SAG基因則分布在進化樹的另一較大分支上,其中同為新孢子蟲屬(Neospora)的N. caninum和N. hughesi SAG1基因序列也分布在同一親緣關系較近的分支上,這與Eimeria屬球蟲SAGs基因的特性相似。另外,GenBank中已發表的4條E. tenella SAG10基因與本研究克隆SAG10基因的親緣關系較近,分布在同一較小分支上,且E. tenella江蘇株和北京株的SAG10基因分布最近,與“2.2”節中江蘇株SAG10基因的BLAST結果一致。
已有研究認為,頂復門原蟲的SAGs編碼基因在組織寄生型原蟲(包囊形成型原蟲,如T. gondii、S. neurona、N. caninum)間是高度同源的,但Eimeria與隱孢子蟲、瘧原蟲及這三者與弓形蟲間無任何同源性[10-11]。本研究發現,T. gondii、N. caninum、N. hughesi、Eimeria球蟲SAGs基因之間確實表現出了上述特點,但例外的是,S. neurona的2條SAG1基因卻與T. gondii、N. caninum、N. hughesi的SAGs基因序列均表現出了較低的同源性,反而與Eimeria球蟲SAGs基因分布在進化樹的同一較大分支上,且與E. tenella的SAG1基因間表現出較高的同源性,這與上述已有觀點明顯不符。
3結論與討論
GPI錨定蛋白是一類通過其羧基末端的糖基化磷脂酰肌醇結構錨定于真核細胞膜表面的蛋白,與傳統的跨膜型表面蛋白不同,它們不跨越細胞膜脂質雙層,只通過其羧基末端的GPI錨定于細胞上,同時這種GPI錨在磷脂酶C的處理下又可從細胞膜上脫落[12]。已發現的GPI錨定蛋白種類很多,包括水解酶、細胞黏附分子、表面抗原、受體、朊病毒等[13]。GPI錨定蛋白功能廣泛,涉及細胞識別、信號轉導、生長發育、分化[CM(25]、程序性死亡等重要生命過程,與血栓形成、白血病等很多[CM)][FL)]
[FK(W22][TPCBF4.tif][FK)]
疾病有著一定聯系[14]。SAGs是一類GPI錨定蛋白,大量分布于弓形蟲、瘧原蟲、隱孢子蟲、新孢子蟲等侵襲性階段蟲體細胞表面,是蟲體與宿主細胞黏附的主要分子[10]。目前,弓形蟲的SAGs已被深入研究,但針對E. tenella SAGs的研究仍有待進一步加強[3,10]。
麥博等對E. tenella楊凌株SAG10基因及其氨基酸序列進行了研究,發現SAG10在不同E. tenella株間存在抗原性差異[2]。簡永利等對E. tenella楊凌株SAG2基因進行研究后也發現了類似現象[10]。但這些發現與認為E. tenella不存在明顯株間抗原性差異的傳統觀點相對立。韓德強等對E. tenella北京株SAG10基因進行了研究,未報道是否發現這一現象[7]。為澄清這一問題,同時為研究E. tenella SAG10的生物學功能以及為篩選理想的E. tenella保護性抗原等提供信息,本研究對E. tenella江蘇株的SAG10基因進行了克隆,并將它與國內外在GenBank中發表的E. tenella豪頓株、北京株、楊凌株的SAG10基因進行了系統比對和分析。結果表明,E. tenella江蘇株SAG10基因與GenBank中收錄的E. tenella北京株SAG10基因同源性最高,高達99%,其后依次為楊凌株、豪頓株SAG10基因。對4種不同E. tenella分離株SAG10基因和推導氨基酸序列進行研究發現,SAG10基因之間共有11個位點存在差異,其中8個堿基突變為有義突變,引起了8個氨基酸突變。分析SAG10蛋白的親水性、抗原性、表面可能性等特性發現,國內外不同E. tenella分離株之間的SAG10蛋白抗原性總體比較接近,且抗原性很強,但國內不同分離株之間的抗原性更加接近,它們均與國外豪頓株的氨基酸序列在140~160位差異較大,且國內分離株的抗原性優于國外豪頓株。這可能是由于E. tenella國內分離株和國外豪頓株的SAG10氨基酸序列在140~160位變化頻率較高,引起蛋白的空間構象、親水性等發生變化而導致。
目前對于E. tenella SAGs的研究深度遠遠落后于弓形蟲等其他復頂門原蟲SAGs,因此在開展E. tenella SAGs的蛋白結構、生物學功能等研究過程中,常須要從弓形蟲等其他復頂門原蟲SAGs的研究中獲得啟發,并進行對比和推測。簡永利等研究發現,E. tenella楊凌株與豪頓株的SAG2基因序列高度同源,但與T.gondii、Neospora caninum、Sarcosystis neurona SAG2無任何同源性[10]。更有觀點認為,頂復門原蟲的SAGs編碼基因在組織寄生型原蟲(包囊形成型原蟲,如T. gondii、S. neurona、N. caninum)間是高度同源的,但Eimeria與隱孢子蟲、瘧原蟲及這三者與弓形蟲間無任何同源性[10-11]。為證實這一觀點,本研究收集了GenBank中已發表的E. tenella 4條SAG10基因和1~2條不等的SAG1、SAG5、SAG8、SAG9、SAG11、SAG12的基因序列,同時收集了E. necatrix、N. caninum、N. hughesi、S. neurona、T. gondii的相關SAG基因序列,結合本研究克隆的E. tenella SAG10基因序列,共同采用MEGA 6.06軟件構建遺傳進化發生樹。結果發現,E. tenella和E. necatrix的SAGs基因家族的序列均分布在進化樹的同一較大分支上,表明同為Eimeria屬的球蟲SAGs之間可能相對比較保守。N. caninum、N. hughesi、T. gondii的SAG基因則分布在進化樹的另一較大分支上,其中,同為Neospora屬的N. caninum和N. hughesi SAG1基因序列也分布在同一親緣關系較近的分支上,這與Eimeria屬球蟲SAGs基因的特性相似,也與上述其他學者的觀點一致。但例外的是,S. neurona的2條SAG1基因卻均與T. gondii、N. caninum、N. hughesi的SAG1基因序列表現出了較低的同源性,反而與Eimeria球蟲SAGs基因分布在進化樹的同一較大分支上,且與E. tenella的SAG1基因間表現出較高的同源性,這與已有研究結果[10-11]明顯不符。導致這一異常現象的原因有待進一步研究,同時須分析比對更多的序列。
參考文獻:
[1]曹利利,姚新華,侯洪烈,等. 柔嫩艾美耳球蟲長春株SAG2基因的克隆與序列分析[J]. 動物醫學進展,2013,34(12):115-120.
[2]麥博,覃宗華,于三科,等. 柔嫩艾美球蟲楊凌株表面抗原SAG10基因的克隆與原核表達[J]. 中國獸醫科學,2007,37(9):751-755.
[3]Tabarés E,Ferguson D,Clark J,et al. Eimeria tenella sporozoites and merozoites differentially express glycosylphosphatidylinositol-anchored variant surface proteins[J]. Molecular&Biochemical Parasitology,2004,135(1):123-132.
[4]Ngo H M,Hoppe H,Joiner K. Differential sorting and post-secretory targeting of proteins in parasitic invasion[J]. Trends in Cell Biology,2000,10(2):67-72.
[5]Frasch A C. Functional diversity in the trans-sialidase and mucin families in Trypanosoma cruzi[J]. Parasitology Today,2000,16(7):282-286.
[6]Lekutis C,Ferguson D,Grigg M,et al. Surface antigens of Toxoplasma gondii:variations on a theme[J]. International Journal for Parasitology,2001,31(12):1285-1292.
[7]韓德強,丁宏標,喬宇,等. 雞柔嫩艾美耳球蟲表面抗原基因sag10在畢赤酵母中的表達及鑒定[J]. 寄生蟲與醫學昆蟲學報,2006,13(4):199-203.[HJ1.5mm]
[8]蔣建林,蔣金書. 柔嫩艾美耳球蟲各階段蟲體純化方法的改進[J]. 中國農業大學學報,1996,1(5):99-102.
[9]薩姆布魯克 J,拉塞爾 D W. 分子克隆實驗指南[M]. 3版.北京:科學出版社,2002:1256-1259.
[10]簡永利,蔡建平,于三科,等. 柔嫩艾美耳球蟲表面抗原SAG2基因的克隆與表達[J]. 畜牧與獸醫,2006,38(6):10-13.
[11]Jung C,Lee C Y,Grigg M E. The SRS superfamily of Toxoplasma surface proteins[J]. International Journal for Parasitology,2004,34(3):285-296.
[12]熊茂林,宋暢,羅榮城,等. 人糖基磷脂酰肌醇(GPI)-B7-1真核表達載體的構建及表達[J]. 中國病理生理雜志,2005,21(1):154-158.
[13]蘇建亞,沈晉良. 棉鈴蟲幼蟲中腸非糖基磷酯酰肌醇錨著氨肽酶N基因的克隆和測序[J]. 昆蟲學報,2005,48(3):444-449.
[14]張云華,黃瑾. GPI錨定蛋白的合成及功能[J]. 農墾醫學,2012,34(1):59-62.
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
財經界(學術版)(2015年20期)2015-12-23 09:20:13
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
