低溫脅迫對黃姑魚(Nibea albiflora)抗氧化酶、Na+/-K+-ATP酶及Hsp70蛋白含量的影響
2017-05-13 03:02:59羅勝玉徐冬冬樓寶陳睿毅詹煒毛國民
海洋通報 2017年2期
羅勝玉,徐冬冬,樓寶,陳睿毅,詹煒,毛國民
(1.浙江海洋大學,浙江舟山316004;2.浙江省海洋水產研究所,浙江省海水增養殖重點實驗室,浙江舟山316100)
低溫脅迫對黃姑魚(Nibea albiflora)抗氧化酶、Na+/-K+-ATP酶及Hsp70蛋白含量的影響
羅勝玉1,2,徐冬冬2,樓寶2,陳睿毅2,詹煒2,毛國民2
(1.浙江海洋大學,浙江舟山316004;2.浙江省海洋水產研究所,浙江省海水增養殖重點實驗室,浙江舟山316100)
為探討低溫處理對黃姑魚幼魚生理機能的影響,以18℃為對照組,設置8℃、10℃和14℃3個低溫脅迫組,測定不同溫度急性脅迫對黃姑魚幼魚(Nibea albiflora)超氧化物歧化酶(SOD)、過氧化氫酶(CAT)、Na+/-K+-ATP酶活力以及熱休克蛋白70(Hsp70)蛋白含量的影響。研究結果表明,低溫脅迫組(8℃、10℃和14℃)的SOD和CAT酶活力隨著脅迫溫度的降低及其脅迫時間的延長呈先升高后降低的趨勢,在72 h時恢復到對照組水平。肌肉中的Na+/-K+-ATP酶活力則隨處理時間的延長呈現先降低后升高的趨勢,10℃和14℃組能夠在72 h時恢復到對照組水平,而8℃組則在72 h仍顯著低于對照組。低溫脅迫下黃姑魚肌肉的Hsp70含量隨著處理時間的延長呈先升高后降低趨勢,10℃和14℃組能夠在72 h時恢復到對照組水平,而8℃組則在72 h仍顯著高于對照組。由此說明,SOD、CAT、Na+/-K+-ATP以及Hsp70蛋白參與了黃姑魚低溫脅迫應答過程,可以作為其低溫脅迫應答的標志物。
微低溫脅迫;黃姑魚;SOD酶;CAT酶;Na+/-K+-ATP酶;Hsp70
黃姑魚(Nibea albiflora)是我國重要的海水經濟魚類,具有生長快、抗逆性強、繁殖力強等特點,是開展近海網箱和池塘養殖的適宜品種(雷霽霖等,1992)。隨著黃姑魚繁育技術日趨成熟,其養殖規模在浙江、福建沿海迅速擴大,成為我國東南沿海重要的養殖魚種。但是,近年來由于全球環境破壞導致氣候異常變化,我國沿海地區冬季常遭寒潮侵襲,致使養殖的黃姑魚大量凍死,尤以浙江沿海地區為甚,導致巨大的經濟損失,嚴重制約了養殖產業的發展。因此,提高養殖黃姑魚的耐低溫能力,培育耐寒品種具有重要意義。
魚類作為變溫動物,其生理狀況與溫度變化密切相關,對低溫的適應過程也是一系列細胞應答、逐級調控全身系統參與抵御的復雜生理過程(Donaldson et al,2008)。低溫脅迫是研究魚類耐寒機理的重要手段,從生理、生化等角度尋找與響應低溫適應的標志物是研究魚類低溫響應過程并解析其低溫適應機制的可行途徑。一般而言,與免疫、代謝等相關的酶在魚類低溫脅迫應答中起到十分重要的作用,受到研究者的關注,其中,抗氧化酶、熱休克蛋白、三磷酸腺苷酶等常常作為檢測魚類低溫脅迫應答的標志物。Heise等(2006)研究了北海綿鳚(Zoarces viviparous)低溫脅迫后抗氧化酶活力變化;Mladineo等(2009)研究了金槍魚在低溫和高溫脅迫下的Hsp70和Na+/K+-ATPase在高溫和低溫適應的表達模式;劉波等(2011)研究了低溫應激對肝臟Hsp70基因表達及吉富羅非魚血清生化指標的影響。
目前,關于黃姑魚的研究主要集中在苗種繁育、養殖技術、生態毒理、病害和雌核發育等方面(雷霽霖等,1992;徐冬冬等,2013;楊育凱等,2013),而關于黃姑魚低溫適應方面的研究較少涉及。本文通過研究低溫脅迫下黃姑魚幼魚超氧化物歧化酶(SOD)、過氧化氫酶(CAT)等抗氧化酶以及Hsp70和Na+/K+-ATPase的變化,探討低溫處理對黃姑魚幼魚生理機能的影響,以期為研究黃姑魚對環境溫度的適應機制和健康養殖提供必要的理論依據和數據支持,并為黃姑魚耐寒品種的選育奠定基礎。
1 材料與方法
1.1實驗材料
實驗用黃姑魚由浙江省海洋水產研究所西軒漁業科技島人工培育,體長為(18.25±2.10)cm,體重為(120.15±2.36)g。實驗開始前于(18.0± 1.0)℃海水暫養2周。所用海水經過沙濾,鹽度28,pH 8.2~8.6,溶氧≥6 mg/L,連續充氣,每日換水1次,換水量50%。
1.2實驗方法
預實驗發現黃姑魚由18℃移至6℃后短時間內出現死亡,而移至8℃后未發現死亡℃,因此設置18℃、14℃、10℃、8℃4個溫度組,每組設置2個平行。每個平行組即1個桶(500 L)放置30尾魚,當水溫降至設定溫度后迅速將魚放入實驗桶內,水溫由海水制冷機循環系統調控,溫度誤差控制±0.2℃。
在0 h、2 h、6 h、12 h、24 h、48 h、72 h分別取樣,每個處理組隨機取3尾,解剖取其鰓絲、肌肉和肝臟,用生理鹽水洗凈,保存于RNA保護液中備用。蛋白含量以牛血清蛋白為標準,采用考馬斯亮蘭法測定。所有酶活力及Hsp70含量均采用南京建成生物公司的試劑盒測定,操作步驟參照說明書。SOD酶活力單位定義為每毫克組織蛋白在1 mL反應液中SOD抑制率達50%時所對應的SOD量為一個SOD活力單位(U)。CAT活力單位定義:每毫克組織蛋白每秒鐘分解1 μmol的H2O2的量為一個活力單位(U)。Na+/-K+-ATP酶活力的單位定義:規定每小時每毫克組織蛋白的組織中ATP酶分解ATP產生1 μmol無機磷的量為一個ATP酶活力單位(U),即微摩爾磷/毫克蛋白/小時(μmolPi/mgprot/h)。Hsp70含量單位為ng/mgprot。
1.3實驗數據處
所得數據均以平均值±標準差的形式表示,各組數據經統計軟件SPSS 22.1中的方差分析(ANOVA)和Duncan氏檢驗法進行顯著性分析,顯著水平設置為P=0.05。采用EXCEL 2007軟件作圖。
2 結果
2.1不同溫度處理的黃姑魚組織的SOD酶活力比較
脅迫溫度和脅迫持續時間顯著影響肝臟、鰓和肌肉組織中的SOD酶活力(Two-way ANOVA,P<0.01)(如圖1)。8℃處理組中肝臟組織的SOD酶活力在2 h時急劇升高,之后隨脅迫時間延長逐漸下降,在72 h時與對照組無顯著性差異(P> 0.05);鰓和肌肉中的SOD酶活力則在6 h時達到最高,隨后逐漸降低,在72 h恢復到對照組水平。10℃處理組中,肝臟和肌肉SOD酶活力在6 h最高,鰓中SOD酶活力則在12 h最高,隨后逐漸降低至對照組水平。14℃處理組中,肝臟SOD酶活力在6 h最高,而鰓和肌肉SOD酶活力則在12 h最高。3種組織中SOD酶活力均呈先升高后下降的趨勢,在72 h與正常對照組無顯著性差異(ANOVA,P>0.05)。
2.2不同溫度處理的黃姑魚組織中CAT酶活力比較
不同低溫脅迫處理組黃姑魚幼魚組織中CAT酶活力的比較(如圖2)。脅迫溫度和脅迫持續時間顯著影響3種組織中的CAT酶活力(Two-way ANOVA,P<0.05)。3種組織的CAT酶活力隨脅迫時間延長呈先升高后下降的趨勢,在72 h恢復至正常對照組水平。8℃、10℃和14℃處理組的肝臟和肌肉組織的CAT活力在脅迫6 h后達到最高,其后逐漸下降,在72 h時與正常對照組無顯著性差異。在鰓組織中,8℃和10℃處理組的CAT酶活力在脅迫2 h后達最高,其后下降至對照組水平。
2.3不同溫度處理黃姑魚的肌肉中Na+/-K+-ATP酶活力比較
不同低溫處理組的黃姑魚幼魚肌肉中Na+/-K+-ATP酶活力比較(如圖3)。8℃組的肌肉組織中Na+/-K+-ATP酶活力隨時間呈先下降后上升的趨勢,Na+/-K+-ATP酶活力在脅迫12 h達到最低,其后酶活力有所升高,但在72 h仍顯著低于對照組(P<0.05)。10℃和14℃組的Na+/-K+-ATP酶活力隨脅迫時間延長呈現先降低后上升的趨勢,10℃和14℃分別在脅迫后12 h和6 h最低,在72 h時與對照組的Na+/-K+-ATP酶活力無顯著性差異(P> 0.05)。
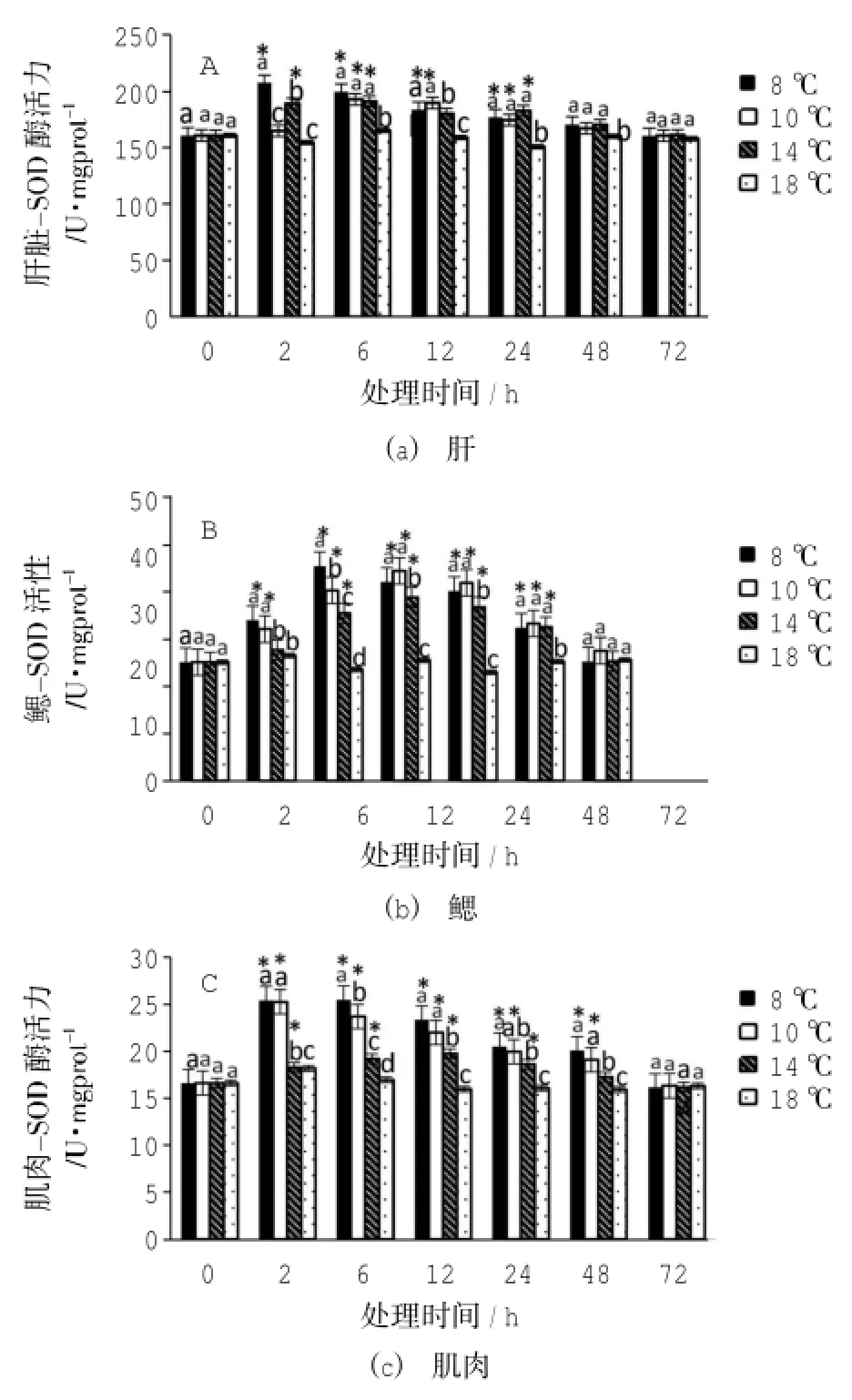
圖1 低溫脅迫下黃姑魚肝臟、鰓和肌肉等組織SOD酶活力
2.4不同溫度處理的黃姑魚肌肉中Hsp70含量比較
脅迫溫度和脅迫時間顯著影響黃姑魚肌肉中的Hsp70含量(Two-way ANOVA,P<0.05)。8℃組肌肉中Hsp70含量在6 h內急劇升高,其后隨脅迫時間延長而降低,在72 h時仍顯著高于對照組(P>0.05)。10℃組和14℃組隨著脅迫時間的延長呈現先升高后降低的趨勢,10℃組Hsp70含量在6 h時最高,14℃組在Hsp70含量在12 h時最高,在72 h時,兩組的Hsp70含量與對照組無顯著性差異(P>0.05)。

圖2 低溫脅迫下黃姑魚肝臟、鰓和肌肉等組織CAT酶活力
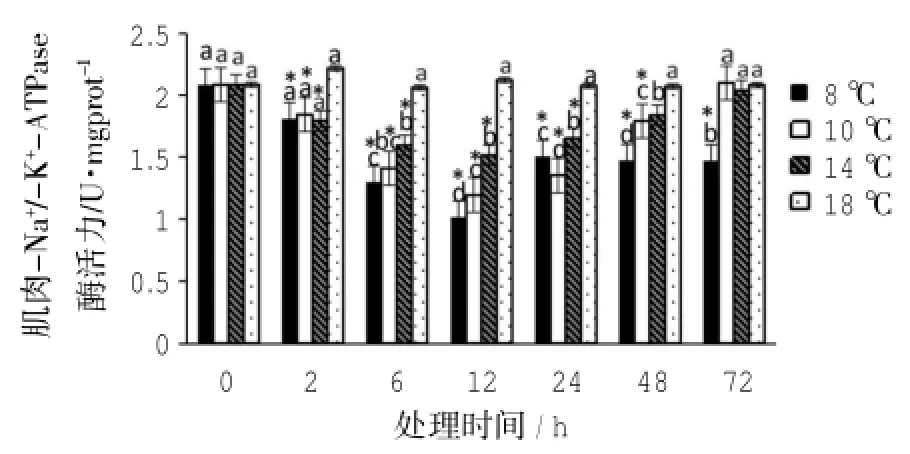
圖3 低溫脅迫下黃姑魚肌肉Na+/-K+-ATP酶活力
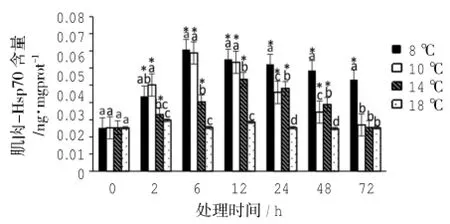
圖4 低溫脅迫下黃姑魚肌肉Hsp70含量
3 討論
3.1低溫脅迫對黃姑魚抗氧化酶活性的影響
低溫脅迫可造成自由基在體內快速大量積累,導致正常細胞和組織的損壞,引發膜脂質過氧化反應,從而引起疾病(Adams et al,1985;Pascual et al,2003;Morales et al,2004;Martínez-álvarez et al,2005;謝妙,2012)。超氧化物歧化化酶(SOD)和過氧化氫酶(CAT)是自由基清除體系中的兩種關鍵酶類,其動態變化可以準確地反映機體內自由基的代謝及組織的氧化損傷情況,對判斷機體的健康狀況及免疫防御能力具有重要的參考價值(嚴建偉,1998;洪美玲等,2007;謝妙,2012)。本研究中,低溫脅迫對黃姑魚肝臟、鰓和肌肉組織的SOD和CAT氧化酶活力具有顯著影響,在脅迫初期兩種氧化酶活力顯著升高,表明黃姑魚在受到溫度脅迫后,其體內的抗氧化還原酶活力會產生相應的響應變化,其他研究中也表明了這點。如卵形鯧鲹(Trachinotusovatus)受到低溫脅迫后,其體內的SOD和CAT等抗氧化還原酶活性在24 h內顯著升高(劉汝建等,2013);對北海綿鳚(Zoarcesviviparous)進行急性冷刺激也發現其肝臟SOD活性顯著增加(Heise et al,2006)。SOD、CAT等酶活力的升高的原因可能是低溫引起體內活性氧自由基(ROS)急劇升高有關,由此誘導機體內抗氧化酶活力逐漸升高以清除體內多余ROS。隨著低溫脅迫時間的延長,各溫度組的SOD和CAT酶活力恢復到對照組水平,表明隨著脅迫時間的延長,黃姑魚幼魚體內的抗氧化酶能夠消除體內自由基的累積,建立新的適應機制,從而SOD和CAT酶活力恢復到到正常水平(Martínez-álvarez et al,2005)。就肝臟、鰓和腎臟三種組織而言,SOD和CAT酶活力受低溫影響的程度在不同組織存在差異,但低溫脅迫后SOD和CAT酶活力均呈現出先升高后降低的趨勢。
3.2低溫脅迫對黃姑魚肌肉中Na+/-K+-ATPase活力的影響
Na+/-K+-ATPase屬于膜束縛酶,低溫能夠引起細胞膜結構的變化從而引起膜的半透性和膜束縛酶的活性等一系列生理過程的改變(徐德立,2002)。Na+/-K+-ATPase酶(又稱Na-K泵)是細胞膜固有的一種蛋白脂質復合體,其主要生理功能是維持生物體正常的生理代謝和能量代謝。許多研究表明,低溫能夠改變Na+/-K+-ATPase酶活力(Haque et al,2011;Galarza-Mu?oz et al,2011;Kang et al,2015),因此,Na+/-K+-ATPase酶常作為評價環境壓力的指標。本研究中,低溫脅迫組的Na+/-K+-ATPase酶活力顯著低于對照組,表明低溫能夠顯著影響黃姑魚Na+/-K+-ATPase酶活力,這與其他水產動物如克氏原螯蝦、吉富羅非魚等的研究結果一致(郭春雨等,2007;王海貞等,2012)。在低溫脅迫的12h內,黃姑魚肌肉的Na+/-K+-ATPase酶活力顯著降低,此后隨脅迫時間的延長各低溫脅迫組的酶活力逐漸升高,10℃和14℃組在72 h達到對照組水平。黃姑魚體內Na+/-K+-ATPase的酶活力變化與抗氧化酶活力變化呈現不同趨勢。Na+/-K+-ATPase酶活力變化一方面可能是由于在低溫脅迫的最初階段,黃姑魚受到冷刺激后生物膜流動性降低,引起膜束縛酶活性降低,從而導致Na+/-K+-ATPase酶活力降低,另一方面可能是由于低溫刺激本身也可使ATPase構象發生改變,進而使其活力下降;其后,隨著脅迫時間延長,魚體逐漸適應周圍環境,調節體內的適應機制達到新的平衡,故Na+/-K+-ATPase酶活性升高(Kang et al,2015)。然而,8℃組Na+/-K+-ATPase酶活力在急劇升高后有所降低,但在脅迫過程中顯著高于18℃組,該組黃姑魚表現出游動減少、行動緩慢,推測可能是過低的溫度破壞了細胞膜流動性,無法在魚體內建立新的平衡,導致Na+/-K+-ATPase酶活力降低。
3.3低溫脅迫對黃姑魚Hsp70的影響
熱休克蛋白Hsp70在機體內作為一種分子伴侶而對機體起到保護作用,可調節細胞的生理功能,提高細胞對應激反應的耐受性(謝妙,2012),因此,Hsp70常作為機體受到應激刺激時反映其細胞功能狀態和抗應激能力的指標之一。Hsp70根據其表達情況可以分為組成型和誘導型,前者存在于非應激狀態的正常細胞中,在正常情況下即表達,應激后不變或稍有增加;后者在正常細胞中,表達水平較低甚至無法檢測到,而在應激后則明顯升高(王美垚,2009)。本研究測定了黃姑魚細胞內總的Hsp70蛋白表達量,Hsp70蛋白含量隨脅迫溫度和脅迫時間表現顯著差異,表明低溫誘導了黃姑魚Hsp70蛋白的表達。在實驗中,黃姑魚受到低溫脅迫后肌肉中Hsp70含量在6 h內迅速升高,其他魚類如金槍魚、羅非魚、大西洋鮭(Salmo salar)等在受到冷刺激后其體內Hsp70蛋白表達量有所升高(Takle et al,2005;Mladineo et al,2009;王美垚,2009),這可能與魚體受到刺激后產生誘導Hsp70蛋白合成以提高機體耐受性有關,是魚體對外界環境進行自主適應的生理表現。隨著脅迫時間的延長,14℃組和10℃組能夠降至18℃組水平,表明在魚體內受到低溫刺激后建立新的平衡;但是8℃組卻沒能恢復到18℃組水平,這可能是由于黃姑魚無法建立適應低溫環境的生理平衡,無法應對過冷的刺激。熱休克蛋白合成的調節包括轉錄水平和翻譯水平的調節。本研究僅從蛋白水平研究Hsp70低溫應答的表達模式,對其低溫脅迫后的基因表達模式及其調節機制還需要進一步的研究。
綜上,低溫脅迫下黃姑魚體內SOD、CAT、Na+/-K+-ATPase的酶活力以及Hsp70含量呈現規律性的變化趨勢,表明上述幾種酶參與了黃姑魚低溫脅迫應答過程,可以作為檢測其應答低溫脅迫的標志物。隨著低溫脅迫時間的延長,黃姑魚體內能夠建立適應低溫的生理平衡,其體內的抗氧化酶以及Na+/-K+-ATPase的酶活力和Hsp70含量能夠恢復;但在極端溫度下如8℃,由于低溫造成的損傷嚴重,Na+/-K+-ATPase的酶活力和Hsp70含量無法恢復導致其正常的生理活動無法維持。
致謝:浙江海洋學院孫連蓮、平洪領以及浙江省海洋水產研究所試驗場徐麒祥、馬濤等多人參與實驗與取樣工作,在此一并致謝。
Adams S M,Mclean R B,1985.Estimation of largemouth bass, Micropterus salmoides Lacépède,growth using the liver somatic index and physiological variables.Journal of Fish Biology,26(2): 111-126.
Donaldson M R,Cooke S J,Patterson D A,et al,2008.Cold shock and fish.Journal of Fish Biology,73(7):1491-1530.
Galarza-Mu?oz G,Soto-Morales S I,Holmgren M,et al,2011. Physiological adaptation of an Antarctic Na+/K+-ATPase to the cold.The Journal of experimental biology,214(13):2164-2174.
Haque M S,Roy S K,Shahjahan M,2011.Arsenic impairs the effect of low temperature on the regulation of Na+-K+-ATPase activity in skeletal muscle of fish(Channa punctata).Turkish Journal of Fisheries and Aquatic Sciences,11(3):339-344.
Heise K,Puntarulo S,Nikinmaa M,et al,2006.Oxidative stress and HIF-1 DNA binding during stressful cold exposure and recovery in the North Sea eelpout(Zoarces viviparus).Comparative Biochemistry& Physiology Part A:Molecular&Integrative Physiology,143(4): 494-503.
Kang C K,Chen Y C,Chang C H,et al,2015.Seawater-acclimation abates cold effects on Na+K+-ATPase activity in gills of the juvenile milkfish,Chanos chanos.Aquaculture,446:67-73.
Martínez-álvarez R M,Morales A E,Sanz A,2005.Antioxidant Defenses in Fish:Biotic and Abiotic Factors.Reviews in Fish Biology& Fisheries,15(1-2):75-88.
Mladineo I,Block B A,2009.Expression of Hsp70,Na+/K+ATP-ase, HIF-1α,IL-1β and TNF-α in captive Pacific bluefintuna (Thunnus orientalis)after chronic warm and cold exposure.Journal of Experimental Marine Biology&Ecology,374(1):51-57.
Morales A E,Pérez-Jiménez A,Hidalgo M C,et al,2004.Oxidative stress and antioxidant defenses af8er prolonged starvation in Dentex dentex liver.ComparativeBiochemistry&PhysiologyToxicology& Pharmacology,139(1-3):153-161.
Pascual P,Pedrajas J R,Toribio F,et al,2003.Effect of food deprivation on oxidative stress biomarkers in fish(Sparus aurata).Chemicobiological interactions,145(2):191-199.
Takle H,Baeverfjord G,Lunde M,et al,2005.The effect of heat and cold exposure on Hsp70 expression and development of deformities during embryogenesis of Atlantic salmon(Salmo salar).Aquaculture, 249(1):515-524.
郭春雨,管越強,劉波蘭,2007.溫度、pH和鹽度對克氏原螯蝦鰓Na+-K+-ATPase活性的影響.動物學雜志,42(6):96-102.
洪美玲,陳立僑,顧順樟,等,2007.不同溫度脅迫方式對中華絨螯蟹免疫化學指標的影響.應用與環境生物學報,13(6):818-822.
雷霽霖,陳超,徐延康,等,1992.黃姑魚工廠化育苗技術研究.海洋科學,11(6):5-10.
劉波,王美垚,謝駿,等,2011.低溫應激對吉富羅非魚血清生化指標及肝臟Hsp70基因表達的影響.生態學報,31(17): 4866-4873.
劉汝建,區又君,李加兒,等,2013.鹽度、溫度對卵形鯧鲹選育群體肝抗氧化酶活力的影響.動物學雜志,48(3):428-436.
王海貞,王輝,強俊,等,2012.溫度和鹽度對吉富品系尼羅羅非魚幼魚鰓Na+-K+-ATPase活力的聯合效應.生態學報,32(3):898-906.
王美垚,2009.急性低溫脅迫及恢復對吉富羅非魚血清生化、免疫以及應激蛋白Hsp70基因表達的影響.南京農業大學.
謝妙,2012.低溫脅迫對斜帶石斑魚生理、生化、脂肪酸的影響.廣東海洋大學.
徐德立,2002.低溫脅迫對草魚ZC-7901細胞系和淡水白鯧CBS細胞系某些細胞功能影響的研究.浙江大學.
徐冬冬,樓寶,薛寶貴,等,2013.黃姑魚(Nibea albiflora)雌核發育的人工誘導及鑒定.海洋與湖沼,44(2):310-317.
嚴建偉,阮積惠,1998.生物體系中自由基檢測方法評述.浙江大學學報:理學版,25(3):75-80.
楊育凱,簡林江,王志勇,等,2013.黃姑魚正常二倍體和雌核發育體胚胎發育及早期生長的比較研究.上海海洋大學學報,22(5):690-697.
(本文編輯:袁澤軼)
Effects of low temperature stress on activities of antioxidant enzymes,Na+/-K+-ATP enzyme and Hsp70 content of Nibea albiflora
LUO Sheng-yu1,XU Dong-dong2,LOU Bao2,CHEN Rui-yi2,ZHAN Wei2,MAO Guo-ming2
(1.ZhejiangOcean University,Zhoushan 316004,China;2.Zhejiang Province Key Lab of MaricultureandEnhancement, Marine Fishery Institute of Zhejiang Province,Zhoushan 316021,China)
In order to investigate the effects of low temperature treatment on the physiological function of juvenile Nibea albiflora,we investigated the effect of cold stress on activities of antioxidant enzymes,Na+/-K+-ATP enzyme and Hsp70 content in juvenile Nibeaalbiflora.Four treatments were set up,and 18℃was as the control and 8℃,10℃and 14℃groups were as experimental groups.We measured the activities of antioxidant enzymes,Na+K+-ATP enzymes and Hsp70 content of N.albiflora at different temperatures.The results showed that the temperature significantly influenced the SOD and CAT enzyme activities,and Na+/-K+-ATP enzyme activities and Hsp70 content of N.albiflora.The results showed that antioxidant enzyme activities increased in the beginning and then decreased under the low temperature stress,and at 72 h post-stress, antioxidant enzyme activities in low temperature treatments were at the same level as the control(P>0.05).Na+/-K+-ATP enzyme activities in the muscles of low temperature treatments showed the trend of decreasing firstly and then rising during the process of cold temperature stress.The Na+/-K+-ATP enzyme activities of 10℃and 14℃treatments could recover and have no significant differences at 72 h post-stress compared with the 18℃treatment,but the activities of Na+/-K+-ATP enzyme in 8℃treatment remained lower at 72 h post-stress.The Hsp70 contents in different treatments were also impacted by the low temperature,and showed the trend of increasing firstly and then decreasing during the process of cold temperaturestress.Compared with the control,the Hsp70 contents of 10℃and 14℃treatments recovered and showed insignificant differences at 72 h post-stress.However,the Hsp70 contents remained higher than that of control at 72 h post-stress. Therefore,SOD,CAT,Na+/-K+-ATPase and Hsp70 were involved in the process when N.albiflora was responding to low temperature stress,and they could be the markers corresponding to cold stress in N.albiflora.
low temperature stress;Nibea albiflora;superoxide dismutase enzyme;catalase enzyme;Na+/-K+-ATP enzyme;heat shock protein 70
P735
A
1001-6932(2017)02-0189-06
10.11840/j.issn.1001-6392.2017.02.010
2015-12-31;
2016-04-06
國家自然科學基金(41476127);浙江省科技計劃項目(2012C12907-8;2015F10001)。
羅勝玉(1990-),男,碩士研究生,主要從事水產動物遺傳育種研究。電子郵箱:548559704@qq.com。
徐冬冬,教授級高工。電子郵箱:xudong0580@163.com;樓寶,研究員。電子郵箱:loubao6577@163.com。
