吉林蛟河針闊混交林樹木生長的空間關聯格局
2017-05-15 12:36:06郝珉輝張忠輝趙珊珊張春雨趙秀海
生態學報 2017年6期
郝珉輝, 張忠輝, 趙珊珊, 張春雨,*, 趙秀海
1 北京林業大學森林資源與生態系統過程北京市重點實驗室, 北京 100083 2 吉林省林業科學研究院, 長春 130033
吉林蛟河針闊混交林樹木生長的空間關聯格局
郝珉輝1, 張忠輝2, 趙珊珊2, 張春雨1,*, 趙秀海1
1 北京林業大學森林資源與生態系統過程北京市重點實驗室, 北京 100083 2 吉林省林業科學研究院, 長春 130033
以吉林蛟河21.12hm2(660m×320m)針闊混交林樣地為對象,利用2009年和2014年森林生長觀測數據,研究樹木生長的空間自相關格局及其生境影響機制。在樣地生境型劃分結果的基礎上,采用Ripley′sL(r)函數分析不同生境型中樹木種群空間分布特征;利用標記相關函數分析不同生境型中樹木生長特征的空間關聯格局。研究結果表明:(1)紅松(生境型3:1—5m)、蒙古櫟(生境型3:1—3m)、胡桃楸(生境型2:1—2m;生境型3:1—7m)、黃檗(生境型2:1—3m;生境型4:1—5m)、水曲柳(生境型3:1—2m;生境型4:1—2m)、瘤枝衛矛(生境型2:1—15m)在特定生境和空間尺度上呈隨機分布,但空間格局仍以聚集性分布為主;其余10個物種則在全部0—30m尺度上呈聚集分布。(2)標記相關函數分析顯示春榆、毛榛、色木槭、瘤枝衛矛和千金榆的徑向生長至少在一個生境中表現出正相關格局;暴馬丁香、胡桃楸、裂葉榆、瘤枝衛矛、水曲柳、紫椴、糠椴、毛榛、色木槭和白牛槭的徑向生長至少在一個生境中表現出負相關格局;紅松、黃檗、蒙古櫟和簇毛槭的徑向生長在全部尺度上均未檢測到顯著的空間關聯格局。因此,不同樹種徑向生長的空間自相關特征不同,樹種生長特征的空間關聯格局具有明顯的生境依賴性。
徑向生長; 生境差異; 競爭作用; 生境依賴性; Ripley′sL(r)函數
森林中普遍存在著種內和種間相互作用,林木個體之間的相互作用可大體分為正向作用和負向作用[1- 2]。由于土壤水分、礦質元素和光照等環境資源的有限性,生長在同一棲息地上的樹木往往存在競爭作用,表現為生長上的負相關;為了阻止其它物種進入某個棲息地,該棲息地上的樹木則會彼此之間產生促進作用,即生長上的正相關[1]。相鄰樹木之間的相互作用對森林群落的空間格局存在著深遠影響。大量研究顯示森林中不同生長階段的樹木分布格局也不盡相同,小徑級樹木多呈聚集分布,隨著徑級的增加聚集強度逐漸減弱,最終甚至呈現均勻或隨機分布[3- 5]。這種空間格局變化很大程度上是由樹木之間的相互作用導致的:在個體水平上,樹木的生長、擴散和死亡等過程都會受到相鄰樹木的影響[5];在群體水平上,相鄰樹木之間的相互作用影響著森林群落動態和林木空間分布格局[3- 5]。相鄰樹木間的相互作用受林木的種類、特征和相對位置影響[1, 6],同時也受外界環境條件影響[7]。空間自相關性是指地理空間中某種空間單元之間的關系,以空間上的相鄰性為基本特征。彼此靠近的空間位置上空間單元的屬性值越相近,空間相關性就越強[8]。空間自相關性是自然界中存在秩序、格局和多樣性的一個根本原因[9]。
森林群落組成結構越復雜,揭示樹木生長的空間格局及其影響機制也就越困難。通過樹木特征值的空間關聯性可以有效地推測相應的生態學過程[10- 12],這種研究思路已經成功應用于熱帶雨林[10- 11]和溫帶森林[12]之中。以往研究主要關注于胸徑、樹高和冠幅等樹木大小特征的空間自相關性;相比之下,樹木生長特征對相鄰個體之間的相互作用和生境變化更加敏感。因此,在林分水平上比較環境變異對樹木生長空間自相關格局的影響,可以更加直觀地揭示群落生長格局的構建機制。本文以位于吉林省蛟河林業試驗區管理局的21.12hm2(660m×320m)森林監測樣地為對象,在2009年和2014年森林生長觀測數據的基礎上,研究樹木生長的空間關聯格局及其對生境變異的響應,旨在揭示生境變異對樹種生長格局構建過程的影響機制。
1 研究方法
1.1 研究區概況
研究區位于吉林省蛟河林業試驗區管理局(43°57.897′—53°58.263′N, 127°42.789′—127°43.310′E)。屬于受季風影響的溫帶大陸性氣候,年均溫為3.8℃,其中7月份為最熱月,平均溫度為21.7℃;1月份為最冷月,平均溫度為-18.6℃;年平均降水量為695.9mm。林分類型為典型的近成熟次生針闊混交林;土壤為棕色森林土,土層深度范圍為20—100cm。
2009年建立面積為21.12hm2(660m×320m)的研究樣地;樣地最低海拔為425.3m,最高海拔為525.8m,最大高差為100.5m。記錄樣地中所有胸徑(DBH)≥1cm的木本植物的物種、胸徑、樹高、冠幅和相對位置坐標,并掛牌標記進行長期監測。2014年復測時樣地內觀測到DBH≥1cm的木本植物共計38599株,隸屬于18科30屬46種。物種重要值(IV)計算公式采用:IV(%)=(相對多度+相對頻度+相對優勢度)/3[13]。樣地內重要值位于前五的闊葉樹種依次為胡桃楸(Juglansmandshurica)、色木槭(Acermono)、千金榆(Carpinuscordata)、暴馬丁香(Syringareticulatavar.amurensis)和紫椴(Tiliaamurensis);主要針葉樹種為紅松(Pinuskoraiensis)。為了滿足樣本量需求,本文以所有生境型中株數均大于30的物種為研究對象,共計16個物種如表1所示。
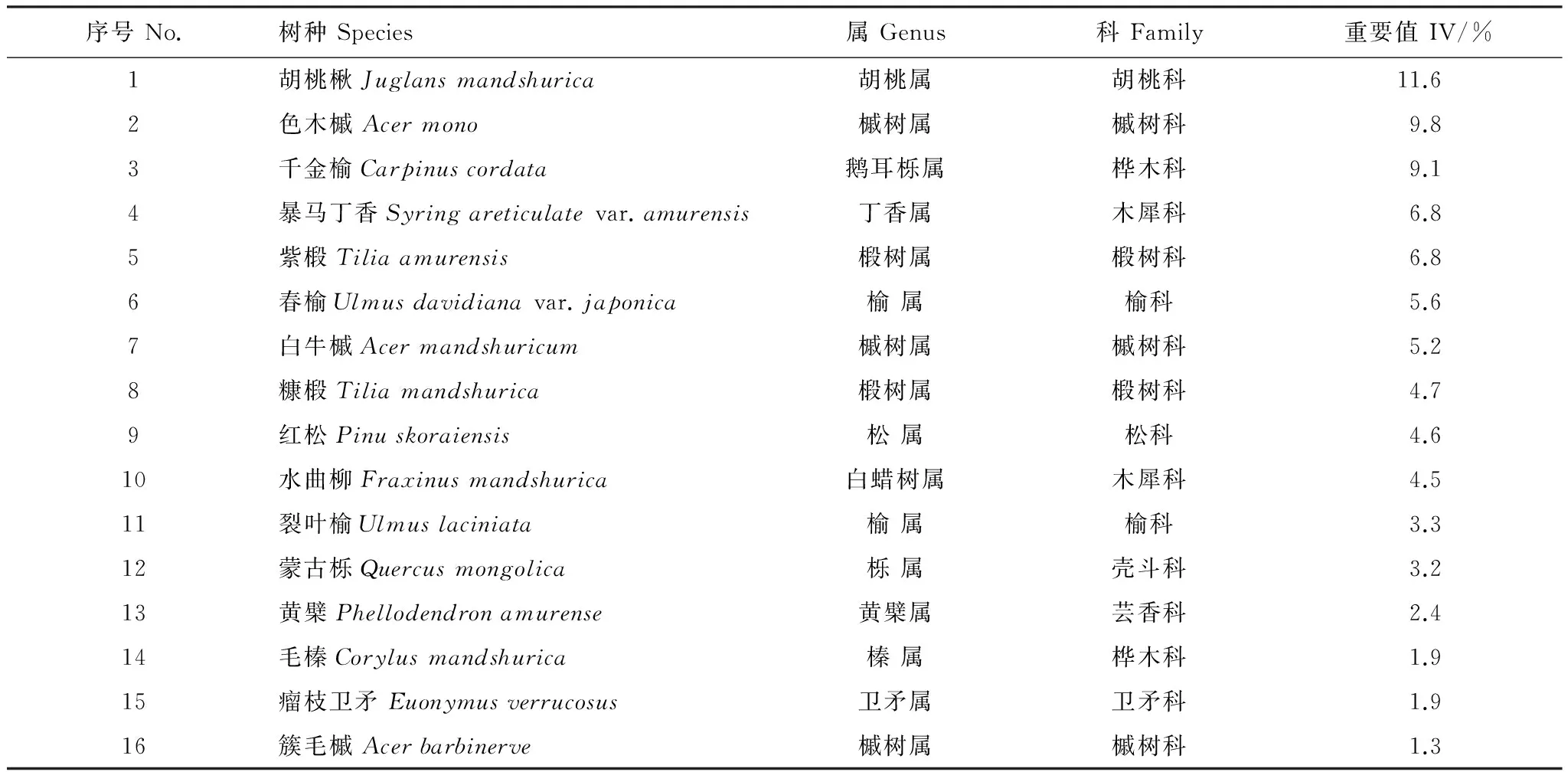
表1 16種木本植物及其重要值
1.2 生境類型劃分
Zhang 等[14]利用多元回歸樹(Multivariate Regression Tree)方法,根據地形變量將研究樣地劃分成4種不同的生境型(圖1)。其中,生境型1包含248個20m×20m樣方,生境指示種有春榆(Ulmusdavidianavar.japonica)、擰筋槭(A.triflorum)、暴馬丁香、金銀忍冬(Loniceramaackii)等;生境型1位于樣地中下方,屬于地勢平緩的低海拔生境。生境型2包含85個20m×20m樣方,生境指示種有千金榆、紅松、青楷槭(A.tegmentosum)等;生境型2位于樣地左上方和右下角,屬于海拔大于453.6m的東坡(坡向≥186度)生境。生境型3和生境型4主要位于樣地右上方和左上角,屬于海拔大于453.6m的西坡(坡向<186度)生境。生境型3包含52個20m×20m樣方,海拔低于465.72m,在樣地中面積最小,指示種有白牛槭(A.mandshuricum)、東北山梅花(Philadelphusschrenkii)、簇毛槭(A.barbinerve)和沙松(Abiesholophylla)等。生境4包含143個樣方,海拔大于465.72m,指示種有色木槭、水榆花楸(Sorbusalnifolia)、裂葉榆(U.laciniata)、糠椴(T.mandshurica)等。
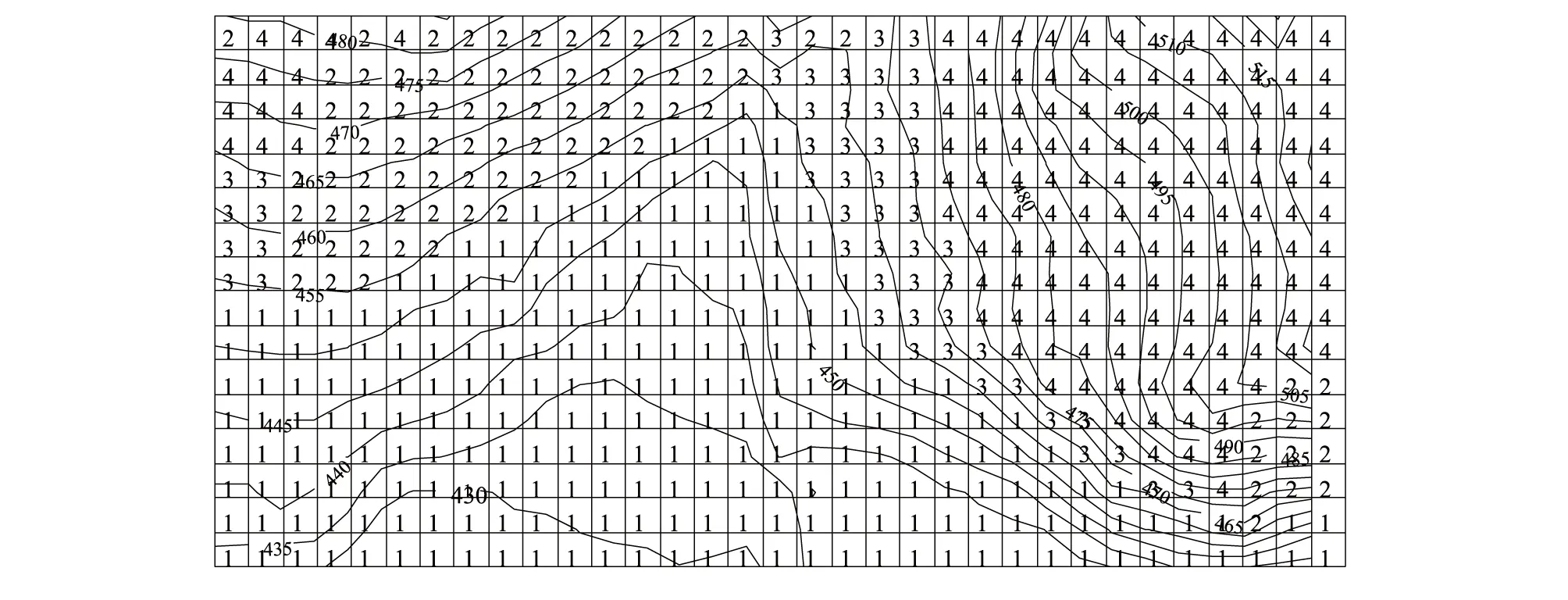
圖1 吉林蛟河21.12hm2研究樣地的生境類型分布Fig.1 The distribution of the habitat types in a 21.12hm2 research plot in Jiaohe, Jinlin Province圖中每個樣方大小為20m×20m, 曲線代表等高距5m的等高線, 數字代表不同生境類型
1.3 空間分析
采用Ripley′sL(r)函數分析樣地內樹種的空間分布特征[15- 16],采用Monte Carlo方法檢驗觀測點的L(r)值偏離隨機分布的顯著性程度[16]。本文通過完全空間隨機模擬技術得到顯著性為95%的置信區間,如果L(r)值落在置信區間內格局為隨機分布,大于置信區間上限為聚集分布,小于置信區間下限為均勻分布;研究尺度r的取值范圍為0—30m。
采用標記相關函數(mark correlation function,MCF)分析樣地中樹木種群的種內空間關聯性[17- 20]。標記相關函數通常用來研究在不同空間尺度上樹木特征值的空間自相關性[5, 12, 18],它不僅考慮研究對象的空間位置,還考慮了研究對象的樹高、胸徑和生長量等數量化特征(即標記值),因此,標記相關函數是一種將樹木的位置坐標和特征參數相結合的函數。本文中特征參數值為樹木5a的胸高斷面積生長量(2009年和2014年胸高斷面積的差值)。標記相關函數同樣采用Monte Carlo方法檢驗觀測點的k[mm](r)值偏離獨立分布的顯著性程度。如果k[mm](r)值落在置信區間內,則表示樹木的生長是相互獨立的、中性的;如果k[mm](r)值落在置信區間上限之上,則表示樹木的生長是相互促進的,具有正向相關性;如果k[mm](r)值落在置信區間下限之下,則表示樹木的生長是相互抑制的,具有負向相關性,即個體間存在競爭關系。
2 結果
2.1 空間位置點格局分析
Ripley′sL(r)函數以物種分布的空間點圖為基礎,量化空間點之間的位置關聯性。本文分析了16個物種在不同生境型中的空間分布格局(表2),紅松、蒙古櫟(Quercusmongolica)、胡桃楸、黃檗(Phellodendronamurense)、水曲柳(Fraxinusmandshurica)和瘤枝衛矛(Euonymusverrucosus)主要呈聚集性分布,但在特定生境中呈現小尺度隨機分布,空間格局具有明顯的生境依賴性。以上6個物種呈現隨機分布的特定生境及其空間尺度分別是:紅松(生境型3:1—5m)、蒙古櫟(生境型3:1—3m)、胡桃楸(生境型2:1—2m;生境型3:1—7m)、黃檗(生境型2:1—3m;生境型4:1—5m)、水曲柳(生境型3:1—2m;生境型4:1—2m,12—16m)、瘤枝衛矛(生境型2:1—15m)。其余10個物種則在全部研究尺度上呈聚集分布。
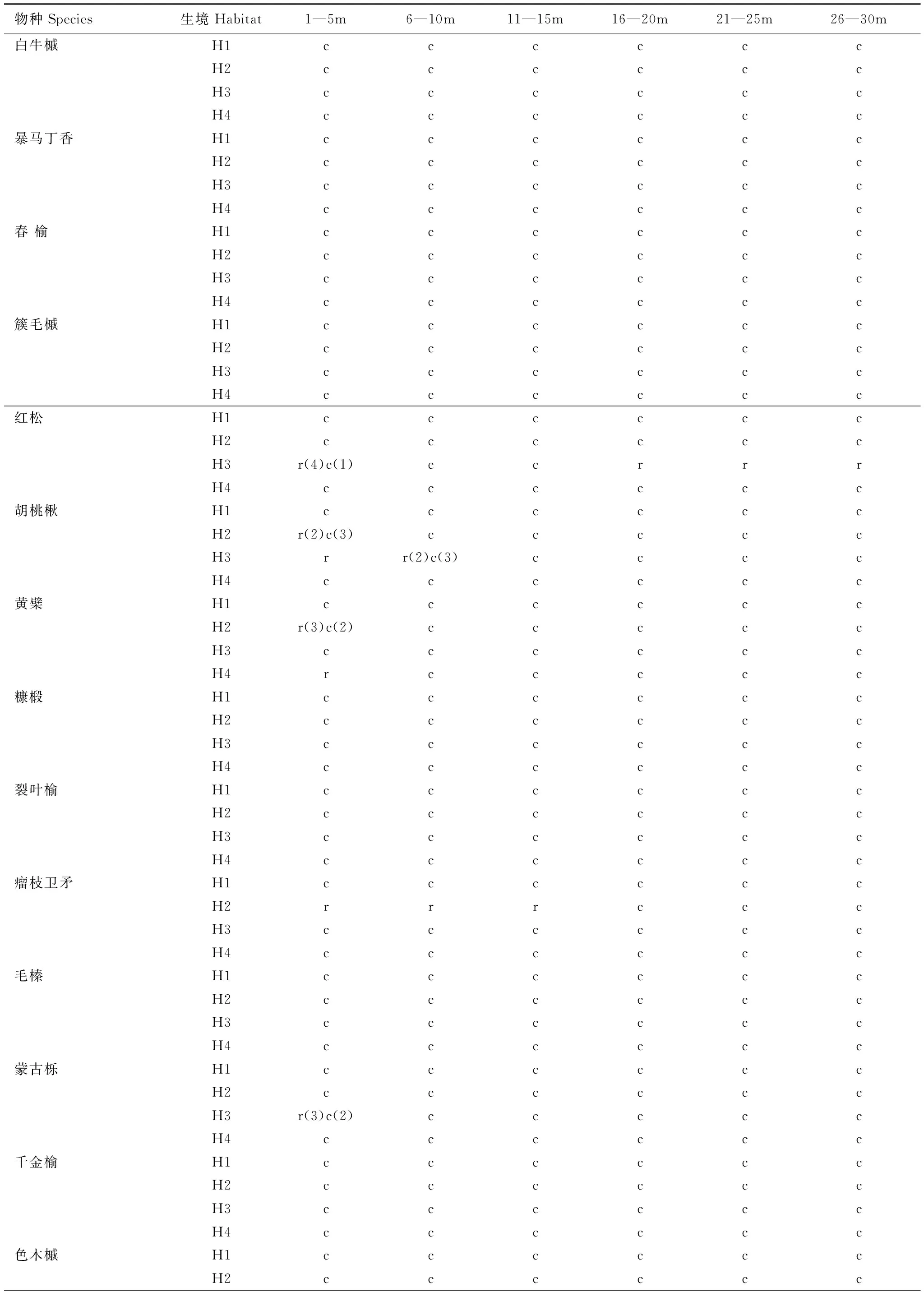
表2 不同生境中物種的空間位置分布特征
r表示隨機分布, c表物種示聚集分布, r(2)c(3)表示在相應的尺度范圍內有2個尺度為隨機分布、3個尺度為聚集分布; H1為生境型1, H2為生境型2, H3為生境型3, H4為生境型4
2.2 樹木生長的空間關聯性分析
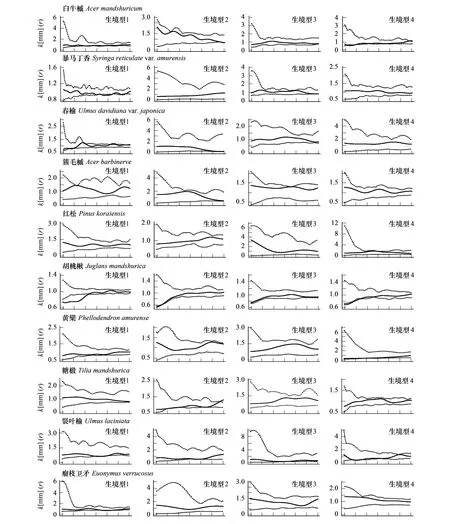
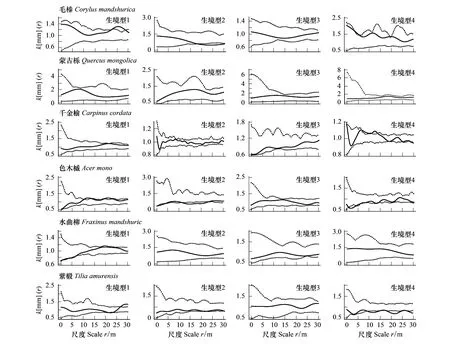
圖2 16個木本植物在不同生境中徑向生長空間關聯性分析Fig.2 Spatial correlations of diameter growth of 16 woody species in different habitats實線為利用觀測數據計算的k[mm]值, 虛線為95%置信區間
標記相關函數分析顯示不同樹種徑向生長的空間自相關特征不同;絕大多數樹種徑向生長的空間特征受生境類型影響,具有明顯的尺度依賴性(圖2)。紅松、黃檗、蒙古櫟和簇毛槭的徑向生長在全部研究尺度上均未檢測到顯著的空間關聯性。至少在一個生境中表現出正相關的物種及其特定的空間尺度為:春榆(生境型1:23—28m)、毛榛(Corylusmandshurica)(生境型1:23—28m)、色木槭(生境型1:11—13m、20—22m)、瘤枝衛矛(生境型4:4—6m)和千金榆(生境型4:7—13m)。至少在一個生境中表現出負相關的物種及其特定的空間尺度為:暴馬丁香(生境型1:1m—21m,26—28m)、胡桃楸(生境型1:1—11m;生境型2:1—3m;生境型4:1—6m)、裂葉榆(生境型1:15—20m;生境型4;4—8m)、瘤枝衛矛(生境型1:18—21m)、水曲柳(生境型1:4—7m)、紫椴(生境型1:22—23m;生境型2:20—23m;生境型4:10—30m)、糠椴(生境型2:5—11m)、毛榛(生境型2:18—30m)、色木槭(生境型2:1—3m,12—20m;生境型4:1—7m)和白牛槭(生境型4:1—5m)。
統計不同物種的林木徑向生長空間自相關類型比例(圖3),生境型1中林木徑向生長呈正相關和負相關的物種比例差異最小,且正相關比例明顯高于其它生境;生境型2中未檢測到正相關,超過37%的物種的徑向生長呈負相關;生境型3中未檢測到徑向生長呈顯著正、負相關的物種;生境型4與生境型1的比例結構類似,但生境型4中正相關的比例相對更小。
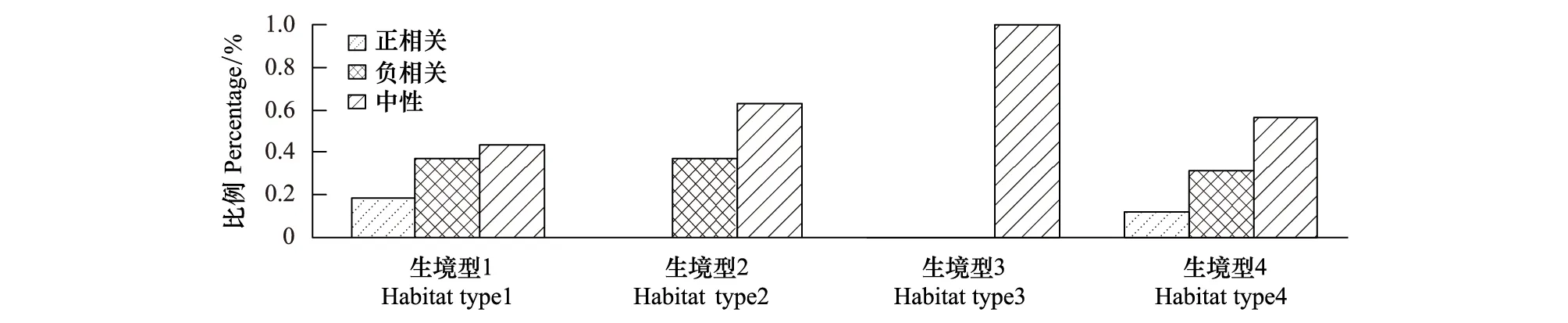
圖3 不同生境型中物種徑向生長的空間關聯性統計Fig.3 Proportion of the intra-specific relationship for all studied species in different habitats
3 討論
熱帶雨林和亞熱帶常綠闊葉林的木本植物空間格局主要表現為聚集性分布,聚集程度隨著空間尺度而變化[21- 23],溫帶森林的物種空間格局也具有類似結構[12]。物種空間分布通常受生境異質性[24]、種子擴散限制[25- 26]、植物種內和種間作用[4, 11]等諸多因素綜合影響。本文以研究樣地生境型劃分結果為基礎[14],在環境條件相對一致的生境型內討論問題,有助于更好地區分生境異質性和樹木之間的相互作用對群落格局構建的影響。
生境條件顯著地影響著樹木生長的空間自相關格局。生境型1位于樣地內地勢平緩的低海拔區域,該區域光照和土壤條件相對較好、水分充足,因此生境型1中正相關比例最高。生境型2和生境型3是樣地內低海拔到高海拔的過渡區域。生境型2位于半陰坡并且坡度較陡,生境型2內主要表現為種內生長的空間負相關,表明該區域種內競爭激烈,可能與其生境條件相對惡劣、土壤水分和光照條件較差有關。生境型3位于半陽坡,坡度較緩、光照條件相對較好,且生境型3擁有更多頂級樹種、群落結構相對更加穩定,因此全部物種的徑向生長均表現為中性關系。生境型2和生境型4都位于樣地內坡度較大的區域,樹種組成頗為相似,但生境型4的海拔相對較高,Coomes 等[7]發現隨著海拔升高植物對光照的競爭會減弱。生境型4中種內生長的負相關比例明顯減少、正相關比例明顯增加,這可能是光照競爭被削弱導致的。
許多研究表明除生境條件外,林分水平上的樹木位置分布特征會對樹木生長的空間關聯性產生影響[1, 6, 27];當樹木呈聚集分布時,樹木的生長會表現出明顯的負相關,負相關強度會隨著聚集強度的增加而增加[27]。本文結果顯示:雖然林木位置在空間上呈聚集分布,但大多數物種的生長空間關聯格局卻是中性的。Law 等[5]研究發現樹木生長的空間關聯格局和樹木位置的空間分布格局在存在一定程度上的獨立性。樹木的空間位置格局主要由出生和死亡等過程決定的,樹木生長則主要由能量流入與養分獲取等過程決定。如果這些潛在生態學過程在一定程度上解耦,就會引起樹木生長的空間自相關性格局與樹木位置的空間分布格局相互獨立[5]。但事實上這兩個過程又常常密不可分:森林中生長發育長期受到抑制的樹木最終會死亡,生長上的負相關性導致樹木格局由聚集分布變成均勻分布[11]。Kubota 等[4]分析針闊混交林在10年間的空間動態,發現樹木聚集分布是由具有空間依賴性的更新過程引起的,并在持續的競爭作用下會逐漸變成均勻分布。Getzin 等[24]研究表明生境異質性導致物種聚集分布,但同時物種對于特定生境的依賴和偏好會削弱聚集分布引起的競爭,從而使樹木生長的空間自相關格局表現為中性。Das 等[28]研究發現雖然競爭對于樹木的生長和死亡存在持續的影響,但這種影響往往比較微弱,只在特定的環境中作用顯著。因此要全面探究群落中樹木生長格局的構建機制,有必要在研究樹木生長自相關格局的基礎上分析生境偏好對植物生長的影響以及植物生長與生境之間的關聯性,這也是我們未來的研究重點。
[1] W?lder K, W?lder O. Analysing interaction effects in forests using the mark correlation function. iForest-Biogeosciences and Forestry, 2008, 1(1): 34- 38.
[2] del Río M, Condés S, Pretzsch H. Analyzing size-symmetric vs. size-asymmetric and intra-vs. inter-specific competition in beech (FagussylvaticaL.) mixed stands. Forest Ecology and Management, 2014, 325: 90- 98.
[3] Stoll P, Bergius E. Pattern and process: competition causes regular spacing of individuals within plant populations. Journal of Ecology, 2005, 93(2): 395- 503.
[4] Kubota Y, Kubo H, Shimatani K. Spatial pattern dynamics over 10 years in a conifer/broadleaved forest, northern Japan. Plant Ecology, 2007, 190(1): 143- 157.
[5] Law R, Illian J, Burslem D F R P, Gratzer G, Gunatilleke C V S, Gunatilleke I A U N. Ecological information from spatial patterns of plants: insights from point process theory. Journal of Ecology, 2009, 97(4): 616- 628.
[6] Getzin S, Wiegand K, Schumacher J, Gougeon F A. Scale-dependent competition at the stand level assessed from crown areas. Forest Ecology and Management, 2008, 255(7): 2478- 2485.
[7] Coomes D A, Allen R B. Effects of size, competition and altitude on tree growth. Journal of Ecology, 2007, 95(5): 1084- 1097.
[8] Tobler W R. Cellular geography // Gale S, Olsson G, eds. Philosophy in geography. Netherlands: Springer, 1979: 379- 386.
[9] Goodchild M F. The application of advanced information technology in assessing environmental impacts // Corwin D L, Loague K, eds. Applications of GIS to the Modeling of Non-Point Source Pollutants in the Vadose Zone. Madison: Soil Science Society of America, Inc., 1996: 1- 17.
[10] Parrott L, Lange H. Use of interactive forest growth simulation to characterise spatial stand structure. Forest Ecology and Management, 2004, 194(1/3): 29- 57.
[11] Picard N, Bar‐Hen A, Mortier F, Chadoeuf J. Understanding the dynamics of an undisturbed tropical rain forest from the spatial pattern of trees. Journal of Ecology, 2009, 97(1): 97- 108.
[12] Zhang C Y, Wei Y B, Zhao X H, Gadow K V. Spatial characteristics of tree diameter distributions in a temperate old-growth forest. PLoS ONE, 2013, 8(3): e58983.
[13] 方精云, 王襄平, 沈澤昊, 唐志堯, 賀金生, 于丹, 江源, 王志恒, 鄭成洋, 朱江玲, 郭兆迪. 植物群落清查的主要內容、方法和技術規范. 生物多樣性, 2009, 17(6): 533- 548.
[14] Zhang C Y, Zhao Y Z, Zhao X H, Gadow K V. Species-habitat associations in a northern temperate forest in China. Silva Fennica, 2012, 46(4): 501- 519.
[15] Ripley B D. The second-order analysis of stationary point processes. Journal of applied probability, 1976, 13(2): 255- 266.
[16] Diggle P J. Statistical Analysis of Spatial Point Patterns. 2nd ed. New York: Arnold Oxford University Press, 2003.
[17] Stoyan D, Stoyan H. Fractals, Random Shapes, and Point Fields: Methods of Geometrical Statistics. Chichester: Wiley, 1994.
[18] Stoyan D, Penttinen A. Recent applications of point process methods in forestry statistics. Statistical Science, 2000, 15(1): 61- 78.
[19] Schlather M, Ribeiro P J Jr, Diggle P J. Detecting dependence between marks and locations of marked point processes. Journal of the Royal Statistical Society: Series B (Statistical Methodology), 2004, 66(1): 79- 93.
[20] Lancaster J. Using neutral landscapes to identify patterns of aggregation across resource points. Ecography, 2006, 29(3): 385- 395.
[21] Hubbell S P. Tree dispersion, abundance, and diversity in a tropical dry forest. Science, 1979, 203(4387): 1299- 1309.
[22] He F L, Legendre P, LaFrankie J V. Distribution patterns of tree species in a Malaysian tropical rain forest. Journal of Vegetation Science, 1997, 8(1): 105- 114.
[23] Li L, Huang Z L, Ye W H, Cao H L, Wei S G, Wang Z G, Lian J Y, Sun I F, Ma K P, He F L. Spatial distributions of tree species in a subtropical forest of China. Oikos, 2009, 118(4): 495- 502.
[24] Getzin S, Wiegand T, Wiegand K, He F L. Heterogeneity influences spatial patterns and demographics in forest stands. Journal of Ecology, 2008, 96(4): 807- 820.
[25] Dalling J W, Muller‐Landau H C, Wright S J, Hubbell S P. Role of dispersal in the recruitment limitation of neotropical pioneer species. Journal of Ecology, 2002, 90(4): 714- 727.
[26] Seidler T G, Plotkin J B. Seed dispersal and spatial pattern in tropical trees. PLoS Biology, 2006, 4(11): e344.
[27] Fraver S, D′Amato A W, Bradford J B, Jonsson B G, Jonsson M, Esseen P A. Tree growth and competition in an old‐growthPiceaabiesforest of boreal Sweden: influence of tree spatial patterning. Journal of vegetation science, 2014, 25(2): 374- 385.
[28] Das A, Battles J, Stephenson N L, van Mantgem P J. The contribution of competition to tree mortality in old-growth coniferous forests. Forest Ecology and Management, 2011, 261(7): 1203- 1213.
Spatial autocorrelation patterns of tree growth in a coniferous and broad-leaved mixed forest in Jiaohe of Jilin province
HAO Minhui1, ZHANG Zhonghui2, ZHAO Shanshan2, ZHANG Chunyu1,*, ZHAO Xiuhai1
1TheKeyLaboratoryforForestResources&EcosystemProcessesofBeijing,BeijingForestryUniversity,Beijing100083,China2AcademyofForestryResearchofJilinProvince,Changchun130033,China
This study aimed to determine the spatial autocorrelation patterns of tree growth and the mechanisms influencing habitat heterogeneity. A dataset of observed forest growth was used, with data between 2009 and 2014 from a 21.12 hm2stem-mapped mixed forest plot (temperate coniferous and broadleaved forest) in Jiaohe, Jilin Province. Based on habitat type classifications, Ripley′sL(r) function was adopted to evaluate the spatial distribution characteristics of tree locations. A mark correlation function (MCF) was used to evaluate the spatial autocorrelation characteristics of tree growth. Our results showed that (1)Pinuskoraiensis(habitat type 3: 1—5 m),Quercusmongolica(habitat type 3: 1—3 m),Juglansmandshurica(habitat type 2: 1—2 m; habitat type 3: 1—7 m),Phellodendronamurense(habitat type 2: 1—3 m; habitat type 4: 1—5 m),Fraxinusmandshurica(habitat type 3: 1—2 m; habitat type 4: 1—2 m), andEuonymusverrucosus(habitat type 2: 1—15 m) were randomly distributed at particular scales and for certain habitat types. Nonetheless, they mainly showed clumped distribution patterns. For ten other species, the spatial distribution of tree positions showed an aggregated pattern at a scale of 0—30 m. (2) The mark correlation function analysis indicated that the radial growth ofUlmusdavidianavar.japonica,Corylusmandshurica,Acermono,E.verrucosus, andCarpinuscordatawere positively autocorrelated for at least one of the habitat types. In contrast, the radial growth ofSyringareticulatavar.amurensis,J.mandshurica,Ulmuslaciniata,E.verrucosus,F.mandshurica,Tiliaamurensis,Tiliamandshurica,C.mandshurica,A.mono, andAcermandshuricumwere negatively autocorrelated at least in one of habitat types. Other species did not show any significant autocorrelations at any of the studied scales. Our results indicate that different species had different spatial autocorrelation characteristics of radial growth, as well as significantly different habitat dependencies.
radius growth; habitat differentiation; competition effect; habitat dependency; Ripley′sL(r) function
國家林業公益性行業科研專項項目(201204309); 教育部新世紀優秀人才支持計劃項目(NCET- 12-0781); 國家自然科學基金項目(31200315)
2015- 10- 27;
日期:2016- 08- 02
10.5846/stxb201510272172
*通訊作者Corresponding author.E-mail: zcy_0520@163.com
郝珉輝, 張忠輝, 趙珊珊, 張春雨, 趙秀海.吉林蛟河針闊混交林樹木生長的空間關聯格局.生態學報,2017,37(6):1922- 1930.
Hao M H, Zhang Z H, Zhao S S, Zhang C Y, Zhao X H.Spatial autocorrelation patterns of tree growth in a coniferous and broad-leaved mixed forest in Jiaohe of Jilin province.Acta Ecologica Sinica,2017,37(6):1922- 1930.
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
汽車觀察(2018年10期)2018-11-06 07:05:26
