溫度馴化對(duì)尖頭熱耐受特征的影響
2017-05-16 09:07:21丹沈中源唐瓊英劉煥章
水生生物學(xué)報(bào) 2017年3期
俞 丹沈中源,張 智,張 晨,唐瓊英劉煥章
(1. 中國(guó)科學(xué)院水生生物研究所, 水生生物多樣性與保護(hù)重點(diǎn)實(shí)驗(yàn)室, 武漢 430072; 2. 中國(guó)科學(xué)院大學(xué), 北京 100049)
俞 丹1沈中源1,2張 智1,2張 晨1,2唐瓊英1劉煥章1
(1. 中國(guó)科學(xué)院水生生物研究所, 水生生物多樣性與保護(hù)重點(diǎn)實(shí)驗(yàn)室, 武漢 430072; 2. 中國(guó)科學(xué)院大學(xué), 北京 100049)
為了研究不同馴化溫度對(duì)尖頭(Rhynchocypris oxycephalus)熱耐受特征的影響, 本研究設(shè)置4組水溫(14℃、19℃、24℃和29℃), 對(duì)尖頭馴化兩周, 采用臨界溫度法觀察尖頭的耐受溫度。結(jié)果顯示: 尖頭的熱耐受性受到溫度馴化的影響, 表現(xiàn)為高溫馴化可以升高最大臨界溫度(CTmax), 4個(gè)馴化組的平均CTmax分別為32.29℃、33.23℃、33.40℃和35.71℃; 低溫馴化可以降低最小臨界溫度(CTmin), 平均CTmin分別為0.00、0.10℃、2.10℃和5.27℃; 在適中的溫度(19℃)馴化條件下具有最高的溫度耐受范圍(33.13℃)。在高溫條件下的溫度馴化具有較高的馴化反應(yīng)率, 最大值出現(xiàn)在24—29℃內(nèi)(0.46); 低溫馴化反應(yīng)率最大值出現(xiàn)在29—24℃內(nèi), 為0.63。尖頭在本研究的馴化區(qū)間(14—29℃)內(nèi)的熱耐受區(qū)域面積為478.98℃2, 與溫水性魚類的溫度耐受性相當(dāng), 說明尖頭具有較強(qiáng)的溫度適應(yīng)能力。
尖頭; 熱耐受; 馴化溫度
溫度是影響生物各項(xiàng)生命活動(dòng)的重要環(huán)境因子[1,2]。生物對(duì)環(huán)境溫度變化具有一定的耐受范圍。魚類屬于變溫動(dòng)物, 其生命活動(dòng)嚴(yán)格受其棲息水環(huán)境的限制, 如果水溫超過了魚類的耐受范圍,魚類很可能會(huì)死亡。目前, 全球氣候變化的一個(gè)趨勢(shì)是全球變暖, 氣候變暖將導(dǎo)致河流、湖泊和海洋溫度升高, 這勢(shì)必將對(duì)魚類的分布、形態(tài)、生理和行為等產(chǎn)生重大影響[3]。因此, 對(duì)魚類進(jìn)行熱耐受特征研究具有重要意義。大量的研究表明溫度馴化可以改變魚類的熱耐受性特征[4—6]。對(duì)北美116種淡水魚類的熱耐受研究綜述發(fā)現(xiàn), 其臨界溫度或初始致死溫度與馴化溫度均成顯著線性相關(guān)[7]。目前, 國(guó)內(nèi)淡水魚類的熱耐受特征研究仍較為少見,已經(jīng)開展了南方鲇(Silurus meridionalis)、點(diǎn)籃子魚(Siganus guttatus)等部分魚類的熱耐受研究[8,9]。魚類的熱耐受特征研究對(duì)于魚類的生態(tài)與分布具有重要意義, 也可以為物種保護(hù)措施的制定提供科學(xué)指導(dǎo), 因此, 需要對(duì)更多魚類進(jìn)行熱耐受性和溫度適應(yīng)性的研究[10,11]。近年來, 對(duì)魚類的熱耐受性特征研究普遍采用臨界溫度法 (Critical thermal methodology, CTM)。比起傳統(tǒng)的突變轉(zhuǎn)移法獲得的起始致死溫度 (Incipient Lethal Temperature technique, ILT), CTM法具有實(shí)驗(yàn)魚用量少, 無需致死實(shí)驗(yàn)魚, 操作方便等優(yōu)點(diǎn)[7,11,12]。
1 材料與方法
1.1 實(shí)驗(yàn)魚的來源與馴化
1.2 實(shí)驗(yàn)方案
1.3 數(shù)據(jù)處理
臨界溫度(CTmax和CTmin)為10尾實(shí)驗(yàn)魚失去平衡時(shí)各監(jiān)測(cè)溫度的平均值。
馴化反應(yīng)率(Acclimation response ratio, ARR)計(jì)算公式如下: ARR=ΔCTm/ΔTm。
ΔCTm是指不同馴化溫度條件下的臨界溫度ΔCTmax和ΔCTmin平均值的差值, ΔTm即不同馴化溫度之間的差值。高溫馴化反應(yīng)率=ΔCTmax/ΔTm, 低溫馴化反應(yīng)率=ΔCTmin/ΔTm。
熱耐受區(qū)域(Thermal tolerance polygon)面積是指不同馴化溫度下由CTmax和CTmin所繪出的多邊形面積[15]。
在SPSS 19.0中采用單因素方差分析(ANOVA)統(tǒng)計(jì)溫度處理對(duì)CTmax和CTmin的影響, 采用DUNCAN多重比較CTmax和CTmin有差異馴化組, 顯著性水平P<0.05。在Excel 2013中繪制熱耐受區(qū)域面積。
2 結(jié)果
各馴化溫度下(14℃、19℃、24℃和29℃)的最大臨界溫度(CTmax)和最小臨界溫度(CTmin)分別為32.29℃、33.23℃、33.40℃、35.71℃和0.00、0.10℃、2.10℃、5.27℃(表 1, ANOVA, P<0.05)。由此可見, 尖頭的最大臨界溫度會(huì)隨著馴化溫度的升高而升高, 最小臨界溫度會(huì)隨著馴化溫度的降低而降低。
14—19℃、19—24℃和24—29℃區(qū)間內(nèi)對(duì)應(yīng)的ΔTmax值分別為0.94℃、0.17℃和2.31℃, ΔTmin值分別為0.10℃、2.00℃和3.17℃。因此, 尖頭的高溫耐受馴化反應(yīng)率為0.19 (14—19℃)、0.03 (19—24℃)、0.46 (24—29℃), 低溫耐受馴化反應(yīng)率為0.63 (29—24℃)、0.40 (24—19℃)、0.02 (19—14℃)(表 2)。馴化反應(yīng)率與馴化溫度的關(guān)系為CTmax(y=0.21x+29, R2=0.8590)和CTmin(y=0.36x–5,R2=0.8694), 高溫和低溫的馴化率最大值均出現(xiàn)在24—29℃內(nèi)。
表 1 尖頭在不同馴化溫度條件下的熱耐受范圍Tab. 1 Thermal tolerance of R. oxycephalus at different acclimation temperatures
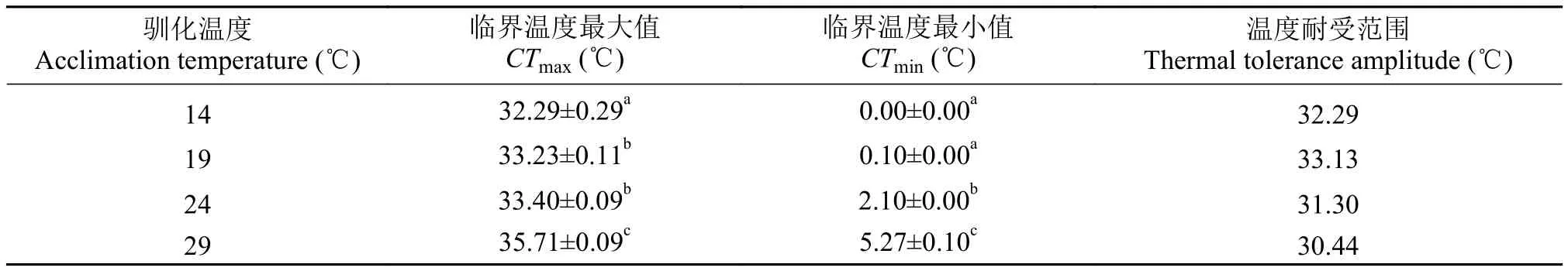
表 1 尖頭在不同馴化溫度條件下的熱耐受范圍Tab. 1 Thermal tolerance of R. oxycephalus at different acclimation temperatures
注: 數(shù)據(jù)為平均值±標(biāo)準(zhǔn)誤, 同一列數(shù)據(jù)上標(biāo)英文字母不同表示存在顯著差異(ANOVA, P<0.05)Note: Data are presented by Mean±SE. Values with different superscript letters within a column indicates significant difference (ANOVA, P<0.05)
溫度耐受范圍Thermal tolerance amplitude (℃) 14 32.29±0.29a0.00±0.00a32.29 19 33.23±0.11b0.10±0.00a33.13 24 33.40±0.09b2.10±0.00b31.30 29 35.71±0.09c5.27±0.10c30.44馴化溫度Acclimation temperature (℃)臨界溫度最大值CTmax(℃)臨界溫度最小值CTmin(℃)
表 2 尖頭不同馴化溫度條件下的馴化反應(yīng)率Tab. 2 Acclimation response ratios of R. oxycephalus at different acclimation temperatures
參數(shù)Parameter馴化溫度范圍Acclimation temperature range (℃)馴化溫度差值ΔT (℃)臨界溫度差值ΔCTm(℃)馴化反應(yīng)率Acclimation response ratio CTmax 14—19 5 0.94 0.19 19—24 5 0.17 0.03 24—29 5 2.31 0.46平均值 15 3.42 0.23 CTmin 29—24 5 0.10 0.63 24—19 5 2.00 0.40 19—14 5 3.17 0.02平均值 15 5.27 0.35
在各馴化溫度(14℃、19℃、24℃和29℃)條件下, 尖頭的溫度耐受范圍(CTmax–CTmin)分別為32.29℃、33.13℃、31.30℃和30.44℃(表 1)。其中最大溫度耐受范圍為33.13℃, 出現(xiàn)在19℃馴化條件下, 對(duì)應(yīng)的臨界溫度最大值和最小值分別為33.23℃和0.1℃。在14—29℃區(qū)間內(nèi), 尖頭的熱耐受區(qū)域面積為478.98℃2(圖 1)。
3 討論
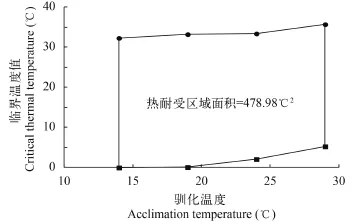
圖 1 尖頭的熱耐受區(qū)域面積Fig. 1 Thermal tolerance polygon of R. oxycephalus
魚類的溫度耐受范圍是指魚類可以耐受的高溫上限和低溫下限之間的范圍, 它是評(píng)價(jià)魚類熱耐受特征的重要指標(biāo)。在未來全球氣候變暖的情況下, 水溫升高會(huì)造成魚類的死亡率和分布范圍發(fā)生改變[3]。根據(jù)魚類對(duì)不同水溫的適應(yīng)情況, 通常將魚類劃分為3類: 冷水性魚類、溫水性魚類和暖水性魚類。通過比較不同環(huán)境溫度條件下魚類的溫度耐受范圍, 發(fā)現(xiàn)溫水性魚類的溫度耐受范圍一般比冷水性魚類和暖水性魚類大。例如, 冷水性魚類虹鱒(Oncorhynchus mykiss)的最大溫度耐受范圍為28.9℃, 其對(duì)應(yīng)的CTmax和CTmin分別為29.1℃和0.2℃[16]; 溫水性魚類南方鲇的最大溫度耐受范圍可達(dá)33.3℃, 對(duì)應(yīng)的CTmax和CTmin分別為38.2℃和5.9℃[8]; 暖水性魚類南亞野鯪(Labeo rohita)的最大溫度耐受范圍僅為27.4℃, 其對(duì)應(yīng)的CTmax和CTmin分別為41.6℃和14.2℃[17]。在本研究中尖頭的最大溫度耐受范圍出現(xiàn)在19℃馴化條件下, 其值為33.1℃, 對(duì)應(yīng)的CTmax和CTmin分別為33.2℃和0.1℃。由此可見, 尖頭與溫水性魚類的最大溫度耐受范圍最為相近, 但最大、最小臨界溫度均小于溫水性魚類。尖頭的最大溫度耐受范圍出現(xiàn)在一個(gè)適中的溫度馴化條件下(19℃), 與其他魚類的熱耐受研究結(jié)論一致[7]。因?yàn)樵谳^低的馴化溫度下, 魚類容易具有較低的低溫耐受值, 但相應(yīng)的高溫耐受值偏低, 反之, 在較高的馴化溫度下, 魚類容易具有較高的高溫耐受值, 但相應(yīng)的低溫耐受值偏高, 所在適中的馴化溫度下, 魚類反而可以獲得更大的溫度耐受范圍。
除溫度耐受范圍以外, 熱耐受區(qū)域面積是評(píng)價(jià)魚類熱耐受特征的另一個(gè)重要指標(biāo), 是指不同馴化溫度下由CTmax和CTmin所繪出的多邊形面積, 單位為℃2。通常來說, 溫水性魚類的熱耐受區(qū)域面積比冷水性魚類和暖水性魚類的大。例如, 暖水性魚類南亞野鯪在14—29℃馴化條件下的熱耐受區(qū)域面積約為399.80℃2, 溫水性魚類南方鲇在相同條件下的熱耐受區(qū)域面積約為470.89℃2。在本研究中尖頭在14—29℃的熱耐受區(qū)域面積為478.98℃2,與溫水性魚類南方鲇的熱耐受區(qū)域面積相近。由此可見, 雖然尖頭分布的自然棲息環(huán)境屬于山區(qū)冷水性溪流, 但是在實(shí)驗(yàn)室條件下, 它依然具有與溫水性魚類相當(dāng)?shù)臏囟饶褪芊秶蜔崮褪軈^(qū)域面積, 這說明尖頭具有較強(qiáng)的溫度適應(yīng)能力。在未來氣候變暖的情況下, 較強(qiáng)的溫度適應(yīng)能力有助于尖頭的生存與擴(kuò)散。
馴化反應(yīng)率(ARR)是指單位馴化溫度(升高或降低1℃)對(duì)魚類臨界溫度(CT)作用的效率, 也是評(píng)價(jià)魚類對(duì)溫度變化生理反應(yīng)的常用指標(biāo), 反映了魚類對(duì)馴化溫度敏感程度的大小。馴化反應(yīng)率越大,表明馴化過程對(duì)魚類耐受溫度范圍的影響作用就越大。在絕大多數(shù)馴化溫度條件下, 低溫馴化反應(yīng)率均大于高溫馴化反應(yīng)率, 在馴化溫度20—30℃條件下, 暖水性魚類大口黑鱸(Micropterus salmoides)的高、低溫馴化反應(yīng)率分別為0.32和0.76[16]; 溫水性魚類南方鲇在相同馴化溫度下的高、低溫馴化反應(yīng)率分別為0.12和0.39[8]; 冷水性魚類虹鱒在馴化溫度10—20℃條件下的馴化反應(yīng)率分別為0.18和0.36[16]。在14—29℃馴化條件下, 尖頭的高、低溫馴化反應(yīng)率分別為0.23和0.35, 該結(jié)果與上述研究的結(jié)論基本一致, 較高的低溫馴化反應(yīng)率表明尖頭對(duì)低溫馴化較為敏感, 暗示尖頭對(duì)低溫環(huán)境可能有較強(qiáng)的適應(yīng)能力。在24—29℃區(qū)間內(nèi), 尖頭的高溫馴化反應(yīng)率為0.46, 是高溫馴化反應(yīng)率中的最大值; 在29—24℃時(shí)的低溫馴化反應(yīng)率為0.63,也是低溫馴化反應(yīng)率中的最大值, 表 3中的數(shù)據(jù)表明尖頭在這兩個(gè)區(qū)間內(nèi)的馴化反應(yīng)率明顯大于其他區(qū)間, 表明溫度馴化對(duì)尖頭的熱耐受性影響程度存在不同。因此, 一定溫度范圍內(nèi)的馴化可以提高尖頭的溫度適應(yīng)性。
致謝:
感謝周卓誠(chéng)在采樣過程中給予的幫助, 感謝王劍偉、張偉偉、羅思在實(shí)驗(yàn)設(shè)備上提供的幫助。
[1]Fry F E J. Effects of the environment on animal activity [J]. University of Toronto Studies, Biological Series No. 55, Publications of Ontario Fisheries Research Laboratory, 1947, 68: 1—62
[2]Willmer P J, Stone G N, Johnston I A. Environmental Physiology of Animals [M]. Oxford: Blackwell Publishing. 2000
[3]Roessig J M, Woodley C M, Cech J J, et al. Effects of global climate change on marine and estuarine fishes and fisheries [J]. Reviews in Fish Biology and Fisheries, 2004, 14(2): 251—275
[4]Heath W G. Thermoperiodism in sea-run cutthroattrout (salmo clarki clarki) [J]. Science, 1963, 142(3591): 486—488
[5]Kowalski K T, Schubauer J P, Scott C L, et al. Interspecific and seasonal differences in the temperature tolerance of stream fish [J]. Journal of Thermal Biology, 1978, 3(3): 105—108
[6]Shafland P L, Pestrak J P. Lower lethal temperatures for fourteen non-native fishes in Florida [J]. Environmental Biology of Fishes, 1982, 7(2): 149—156
[7]Beitinger T L, Bennett W A, McCauley R W. Temperature tolerances of North American freshwater fishes exposed to dynamic changes in temperature [J]. Environmental Biology of Fishes, 2000, 58(3): 237—275
[8]Wang Y S, Cao Z D, Fu S J, et al. Thermal tolerance of juvenile Silurus Meridionalis Chen [J]. Chinese Journal of Ecology, 2008, 27(12): 2136—2140 [王云松, 曹振東, 付世建, 等. 南方鲇幼魚的熱耐受特征. 生態(tài)學(xué)雜志, 2008, 27(12): 2136—2140]
[9]Wang Y, Song Z M, Liu J Y, et al. Thermal tolerance of juvenile Siganus Guttatas [J]. Marine Fisheries, 2015, 37(3): 253—258 [王妤, 宋志明, 劉鑒毅, 等. 點(diǎn)籃子魚幼魚的熱耐受特征. 海洋漁業(yè), 2015, 37(3): 253—258]
[10]Cook A M, Duston J, Bradford R G. Thermal tolerance of a northern population of striped bass Morone saxatilis [J]. Journal of Fish Biology, 2006, 69(5): 1482—1490
[11]Eme J, Bennett W A. Critical thermal tolerance polygons of tropical marine fishes from Sulawesi, Indonesia [J]. Journal of Thermal Biology, 2009, 34(5): 220—225
[12]Cowles R B, Bogert C M. A preliminary study of the thermal requirements of desert reptiles [J]. Bulletin of The American Museum of Natural History, 1944, 83: 261—296
[13]Chen Y Y. Fauna Sinica, Osteichthyes, CypriniformesⅡ[M]. Beijing: Science Press. 1998, 86—87 [陳宜瑜.《中國(guó)動(dòng)物志-硬骨魚綱鯉形目》中卷, 北京: 科學(xué)出版社. 1998, 86—87]
[14]Yu D, Chen M, Zhou Z C, et al. Global climate changewill severely decrease potential distribution of the East Asian coldwater fish Rhynchocypris oxycephalus (Actinopterygii, Cyprinidae) [J]. Hydrobiologia, 2012, 700(1): 23—32
[15]Dalvi R S, Pal A K, Tiwari L R, et al. Thermal tolerance and oxygen consumption rates of the catfish Horabagrus brachysoma (Günther) acclimated to different temperatures [J]. Aquaculture, 2009, 295(1—2): 116—119
[16]Currie R J, Bennetf W A, Beitinger T L. Critical thermal minima and maxima of three freshwater game—fish species acclimated to constant temperatures [J]. Environmental Biology of Fishes, 1998, 51(2): 187—200
[17]Chatterjee N, Pal A K, Manush S M, et al. Thermal tolerance and oxygen consumption of Labeo rohita and Cyprinus carpio early fingerlings acclimated to three different temperatures [J]. Journal of Thermal Biology, 2004, 29(6): 265—270
[18]Rajaguru S. Critical thermal maximum of seven estuarine fishes [J]. Journal of Thermal Biology, 2002, 27(2): 125—128
[19]Bevelhimer M, Bennett W. Assessing cumulative thermal stress in fish during chronic intermittent exposure to high temperatures [J]. Environmental Science & Policy, 2000, 3: S211—S216
[20]Baker S C, Heidinger R C. Upper lethal temperature of fingerling black crappie [J]. Journal of Fish Biology, 1996, 48(6): 1123—1129
EFFECT OF TEMPERATURE ACCLIMATION ON THE THERMAL TOLERANCE OF RHYNCHOCYPRIS OXYCEPHALUS
YU Dan1, SHEN Zhong-Yuan1,2, ZHANG Zhi1,2, ZHANG Chen1,2, TANG Qiong-Ying1and LIU Huan-Zhang1
(1. The Key Laboratory of Aquatic Biodiversity and Conservation of Chinese Academy of Sciences, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan 430072, China; 2. University of Chinese Academy of Sciences, Beijing 100049, China)
Temperature is one of the most important environmental factors that impact the physiological activities of fishes. Fishes exhibit thermal tolerance, which are influenced by temperature acclimation. Global climate change is one potential factor affect the survival and growth of fish species. Therefore, investigation of thermal tolerance of various fish species is of great importance. Rhynchocypris oxycephalus is a typical cold-water fish that might be influenced by the future global warming. The current study investigated thermal tolerance of R. oxycephalus using critical thermal methodology by acclimating the fish to 14, 19, 24 and 29℃ temperature for two weeks. The results showed that thermal tolerance of R. oxycephalus regulated by temperature acclimation, and that the critical thermal maxima (CTmax) increased with the increase of temperature acclimation, and that the average CTmaxwere 32.29, 33.23, 33.40 and 35.71℃. The critical thermal minima (CTmin) decreased with the decrease of temperature acclimation, and the average CTminwere 0.00, 0.10, 2.10 and 5.27℃. At the moderate acclimation temperature (19℃), R. oxycephalus has the widest thermal tolerance up to 33.13℃. The higher acclimation temperatures produced higher acclimation response ratio (ARR). The maximum ARR (0.46) of high temperatures occurred in the range of 24—29℃, while the maximum ARR of low temperatures, (0.63) occurred in 29—24℃. A thermal tolerance polygon in the range of 14 to 29℃ showed an area of 478.98℃2, which was similar with eurythermal fishes. These results indicated that R. oxycephalus has strong ability for thermal adaption.
Rhynchocypris oxycephalus; Thermal tolerance; Temperature acclimation
Q178.1
A
1000-3207(2017)03-0538-05
10.7541/2017.69
2016-04-18;
2016-07-11
國(guó)家自然科學(xué)基金(31401968和31272306)資助 [Supported by the National Natural Science Foundation of China (31401968, 31272306)]
俞丹(1985—), 女, 浙江嵊州人; 博士; 主要從事魚類分子進(jìn)化研究。E-mail: yudan@ihb.ac.cn
劉煥章, E-mail: hzliu@ihb.ac.cn
