加利福尼亞兔和新西蘭兔UCP1 基因第5外顯子多態(tài)性和生物信息學(xué)研究
2017-05-17 08:01:12李維林家棟劉若余謝海強(qiáng)李思杜雪
江蘇農(nóng)業(yè)科學(xué) 2016年1期
李維 林家棟 劉若余 謝海強(qiáng) 李思 杜雪琴 肖超能


摘要:本試驗(yàn)以加利福尼亞兔和新西蘭兔為研究對(duì)象,分別用磁珠法動(dòng)物基因組DNA抽提試劑盒提取新西蘭兔和加利福尼亞兔的DNA,通過(guò)構(gòu)建DNA池,擴(kuò)增UCP1基因的第5外顯子及第4內(nèi)含子部分序列。擴(kuò)增產(chǎn)物進(jìn)行雙向測(cè)序,利用DNAStar和BLAST分析測(cè)序結(jié)果。結(jié)果發(fā)現(xiàn):在加利福尼亞兔中沒(méi)有發(fā)現(xiàn)SNP位點(diǎn),在新西蘭兔中有4個(gè)多態(tài)性位點(diǎn),分別為G743T、A744G、C819A和G849T,其中G743T為同義突變,A744G為錯(cuò)義突變,導(dǎo)致編碼的蘇氨酸(Thr)變?yōu)楸彼幔ˋla);C819A、G849T突變位點(diǎn)位于內(nèi)含子區(qū)。多態(tài)位點(diǎn)的出現(xiàn)對(duì)UCP1基因的RNA二級(jí)結(jié)構(gòu)產(chǎn)生的影響表現(xiàn)在其最小自由能由-1.93 MJ/mol變?yōu)?1.932 MJ/mol,同時(shí)SNPs位點(diǎn)對(duì)UCP1基因蛋白結(jié)構(gòu)也有一定影響。
關(guān)鍵詞:UCPl基因;新西蘭兔;加利福尼亞兔;SNPs;RNA二級(jí)結(jié)構(gòu)
中圖分類號(hào):S829.12 文獻(xiàn)標(biāo)志碼:A 文章編號(hào):1002—1302(2016)01—0041—03
解偶聯(lián)蛋白(uneouplingprotein,UCPs)是褐色脂肪細(xì)胞線粒體內(nèi)膜上存在的一種質(zhì)子轉(zhuǎn)運(yùn)蛋白,它存在于動(dòng)物的各組織中,在能量平衡、維持體溫、機(jī)體產(chǎn)熱和代謝等方面都起著巨大的作用。比如,它激活后可以產(chǎn)生質(zhì)子電化學(xué)梯度降低,使呼吸鏈與ATP(三磷酸腺苷)合成過(guò)程解耦聯(lián),使能量以熱的形式釋放;UCP1基因參與機(jī)體能量代謝,對(duì)機(jī)體能量平衡涉及的食物轉(zhuǎn)化效率、靜止代謝率和體重肥胖等性狀具有明顯的效應(yīng),對(duì)肌肉生長(zhǎng)和品質(zhì)的改良作用重大。目前,這個(gè)家族的5種蛋白質(zhì)在魚(yú)、禽類、哺乳動(dòng)物甚至在人類的不同組織線粒體的內(nèi)膜上都已發(fā)現(xiàn)。其中,UCP1是最早發(fā)現(xiàn)的成員。
UCP1是褐色脂肪組織(brown adipose tissue,BAT)中表達(dá)的標(biāo)記基因,它具有解離氧化磷酸化耦聯(lián)功能,如機(jī)體通過(guò)在線粒體中產(chǎn)生能量,三羧酸循環(huán)產(chǎn)生的還原當(dāng)量通過(guò)電子將能量釋放,使H+從線粒體基質(zhì)轉(zhuǎn)移至內(nèi)膜面,形成1個(gè)跨線粒體內(nèi)膜的質(zhì)子電化學(xué)梯度,當(dāng)線粒體的ATP合酶將H+從內(nèi)膜面順梯度運(yùn)回至基質(zhì)面時(shí),其能量可推動(dòng)二磷酸腺苷與磷酸結(jié)合,促使生成ATP。UCP1基因的表達(dá)增加可能是受到去甲腎上腺素促進(jìn),使其與G偶聯(lián)蛋白相關(guān)的環(huán)磷酸腺苷合成,后者又可使蛋白激酶激活,蛋白激酶作用的靶蛋白包括激素敏感性脂肪酶,這種酶一旦被磷酸化激活就可以抑制三酰甘油的貯存,使脂肪分解,非脂化脂肪酸增加,促進(jìn)產(chǎn)熱作用。此外,UCP1表達(dá)增加還有可能受腎上腺素和甲狀腺素等的刺激,從而增加棕色脂肪組織所消耗脂肪酸的活性,因此UCPl被認(rèn)為是2型糖尿病和抵抗肥胖的重要基因。
Clark等發(fā)現(xiàn),UCP1基因的表達(dá)水平對(duì)羔羊出生后的成活率起著重大作用。Bassett等發(fā)現(xiàn),UCP1基因的表達(dá)對(duì)初生羔羊BAT的含量影響重大。然而對(duì)兔線粒體中UCP1基因方面的研究很少,本試驗(yàn)以加利福尼亞兔和新西蘭兔構(gòu)建其DNA池,對(duì)UCP1基因第5外顯子及第4內(nèi)含子部分序列進(jìn)行擴(kuò)增,BLAST分析其測(cè)序結(jié)果篩選多態(tài)位點(diǎn),由DNAStar軟件進(jìn)行序列拼接、校正,并預(yù)測(cè)其對(duì)RNA結(jié)構(gòu)影響和等位基因頻率估算。
1材料與方法
1.1樣品的來(lái)源
從貴州貴陽(yáng)白云牛場(chǎng)大林農(nóng)牧專業(yè)合作社種兔場(chǎng)和貴州貴陽(yáng)修文盛鑫兔業(yè)開(kāi)發(fā)有限公司種兔場(chǎng)采取加利福尼亞兔的組織樣48個(gè)和新西蘭兔組織樣48個(gè),將采取的樣品放在已編好序號(hào)的自封袋中于-20℃冰箱中保存。
1.2構(gòu)建DNA池和引物設(shè)計(jì)、合成
利用磁珠法動(dòng)物基因組(組織)DNA抽提試劑盒提取DNA,用1.0%瓊脂糖凝膠電泳檢測(cè)所提的DNA,取5μL構(gòu)建DNA池。以兔UCPl基因(其GenBank登錄號(hào)為:NC_013683.1)DNA序列利用Primer-BLAST設(shè)計(jì)1對(duì)引物來(lái)擴(kuò)增UCPl基因第5外顯子和第4內(nèi)含子的部分序列,將所設(shè)計(jì)的引物送往生工生物工程(上海)股份有限公司合成。物序列為F:GAGGTAAGTCCATCCCCACG,R:ACTCTTCTA-ACGATGTAGTGTC;退火溫度為59℃;目的片段大小為809 bp。
1.3 DNA的擴(kuò)增和序列的分析
反應(yīng)體系為25μL:2×Taq PCR Master Mix試劑12.5μL,基因組DNA 2.5μL,三蒸水7μL,上、下游引物10 pmol/μL各1.5μL。PCR擴(kuò)增條件為:94℃預(yù)變性5 min;95℃變性30 s,59℃退火30 s,72℃延伸30 s,35個(gè)循環(huán);72℃延伸10 min。用1%瓊脂糖凝膠電泳檢測(cè)PCR產(chǎn)物,并用凝膠成像系統(tǒng)觀察結(jié)果,最后拍照保存。
將擴(kuò)增的產(chǎn)物送往立菲生物技術(shù)有限公司進(jìn)行雙向測(cè)序。用DNAStar軟件校正測(cè)序結(jié)果,通過(guò)BLAST軟件對(duì)SNPs進(jìn)行確定。
1.4測(cè)量峰高及等位基因頻率估算
利用MWSnap軟件對(duì)SNP位點(diǎn)等位基因的相應(yīng)峰高進(jìn)行測(cè)量,可依據(jù)如下公式fi=hi/(h1+h2),i=1,2,對(duì)等位基因頻率進(jìn)行估算。fi表示SNP位點(diǎn)某等位基因頻率,h1表示SNP等位基因1峰的高度,h2表示SNP等位基因2峰的高度。
1.5 RNA二級(jí)結(jié)構(gòu)預(yù)測(cè)和蛋白質(zhì)二級(jí)結(jié)構(gòu)預(yù)測(cè)
通過(guò)在線預(yù)測(cè)軟件:http:∥rna.tbi.univie.ac.at/cgi-bin/RNAfold.cgi,對(duì)UCPl基因突變前后不同DNA序列進(jìn)行RNA二級(jí)結(jié)構(gòu)變化的預(yù)測(cè),通過(guò)蛋白質(zhì)二級(jí)結(jié)構(gòu)在線預(yù)測(cè)軟件:https:∥npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html,對(duì)UCPl基因突變前后不同蛋白氨基酸序列進(jìn)行蛋白質(zhì)二級(jí)結(jié)構(gòu)預(yù)測(cè)。
2結(jié)果與分析
2.1 PCR產(chǎn)物擴(kuò)增
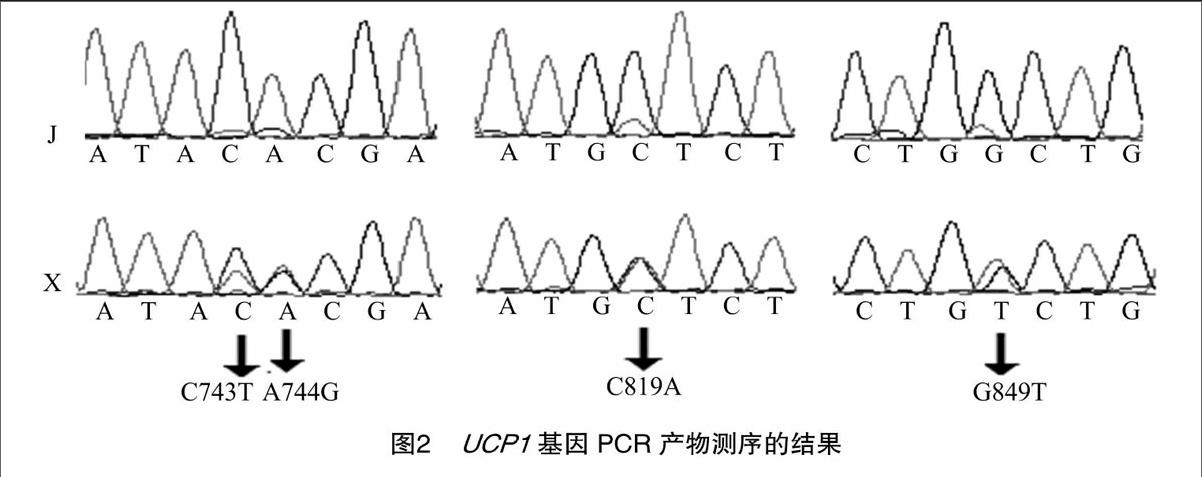
設(shè)計(jì)1對(duì)引物分別擴(kuò)增加利福尼亞兔和新西蘭兔的UCPl基因第5外顯子及第4內(nèi)含子部分序列。PCR擴(kuò)增(圖1)后送往立菲生物技術(shù)有限公司進(jìn)行雙向測(cè)序,BLAST分析共發(fā)現(xiàn)4個(gè)SNPs(圖2)。NCBI中SNP數(shù)據(jù)庫(kù)尚無(wú)該基因SNP信息,本試驗(yàn)所篩查的SNPs均為新發(fā)現(xiàn)多態(tài)性位點(diǎn),以UCP1基因第1外顯子第1位為+1位,SNPs位點(diǎn)分別為G743T、A744G、C819A、G849T,其中G743T為同義突變,A744G為錯(cuò)義突變,導(dǎo)致編碼的蘇氨酸(Thr)變?yōu)楸彼幔ˋla);C819A、G849T突變位點(diǎn)位于內(nèi)含子區(qū)。
2.2估算SNPs等位基因頻率
通過(guò)MWSnap軟件對(duì)加利福尼亞兔和新西蘭兔SNPs等位基因峰高進(jìn)行測(cè)量,根據(jù)公式估算4個(gè)SNPs位點(diǎn)等位基因頻率(表1)。由表1可知,內(nèi)含子C819A和外顯子A744G在加利福尼亞兔和新西蘭兔中有較大差異,而外顯子G743T和內(nèi)含子G849T在2種兔品種中差異較小,且加利福尼亞兔的4個(gè)位點(diǎn)基因頻率均高于新西蘭兔。
2.3預(yù)測(cè)UCP1基因的RNA二級(jí)結(jié)構(gòu)
預(yù)測(cè)突變前后UCP1基因的RNA二級(jí)結(jié)構(gòu),結(jié)果表明,SNPs導(dǎo)致RNA二級(jí)結(jié)構(gòu)發(fā)生改變,并導(dǎo)致RNA二級(jí)結(jié)構(gòu)最小自由能發(fā)生變化,由-1.934 MJ/mol變?yōu)?1.932 MJ/mol,這不僅可能影響RNA二級(jí)結(jié)構(gòu)的穩(wěn)定性,還可能影響后續(xù)蛋白質(zhì)翻譯過(guò)程(圖3)。
2.4突變前后UCP1蛋白二級(jí)的預(yù)測(cè)
通過(guò)在線SOPMA服務(wù)器預(yù)測(cè)新西蘭兔UCP1基因突變前后蛋白質(zhì)二級(jí)結(jié)構(gòu)變化,結(jié)果表明:突變前后α螺旋由109變到110,而B(niǎo)轉(zhuǎn)角由39變成40,延伸鏈由86變到83,自由卷曲由72變到73(表2)。
3討論
大量的研究表明,UCP基因家族與畜禽肉質(zhì)關(guān)系密切。涂榮劍等對(duì)豬的UCP3基因研究發(fā)現(xiàn),突變可能導(dǎo)致相應(yīng)編碼氨基酸序列的改變,從而影響胴體、肉質(zhì)性狀。韓瑞華等對(duì)牛的UCP3基因進(jìn)行研究,發(fā)現(xiàn)UCP3基因不僅對(duì)肉牛胴體、肉質(zhì)性狀等方面具有重要影響,而且對(duì)肉牛的育種工作具有指導(dǎo)作用。王濤等研究了UCP基因?qū)﹄u肉質(zhì)和豬肉質(zhì)量的影響。肌內(nèi)脂肪沉積是兔肉肉質(zhì)標(biāo)記輔助選擇中重要的選擇指標(biāo),篩選出對(duì)家兔肌內(nèi)脂肪沉積有影響的基因有助于加快品種選育進(jìn)程和提高家兔肉質(zhì)性能。
目前對(duì)兔的UCP1基因多態(tài)性研究報(bào)道很少,本試驗(yàn)以加利福尼亞兔和新西蘭兔為研究,對(duì)UCP1基因第5外顯子及第4內(nèi)含子部分序列進(jìn)行擴(kuò)增,將擴(kuò)增的PCR產(chǎn)物進(jìn)行雙向測(cè)序,結(jié)果發(fā)現(xiàn)4個(gè)新的多態(tài)性位點(diǎn):G743T、A744G、C819A、G849T,其中G743T為同義突變,A744G為錯(cuò)義突變,導(dǎo)致編碼的蘇氨酸(Thr)變?yōu)楸彼幔ˋla);C819A、G849T突變位點(diǎn)位于內(nèi)含子區(qū)。比較加利福尼亞兔和新西蘭兔多態(tài)性位點(diǎn)等位基因頻率發(fā)現(xiàn),內(nèi)含子C819A和外顯子A744G在加利福尼亞兔和新西蘭兔中有較大差異,而外顯子G743T和內(nèi)含子G849T在2個(gè)兔品種中差異較小,且加利福尼亞兔的4個(gè)位點(diǎn)基因頻率均高于新西蘭兔。
突變前后UCP1編碼蛋白二級(jí)結(jié)構(gòu)也發(fā)生了改變,B轉(zhuǎn)角由39變成40,α螺旋由109變到110,延伸鏈由86變到83,自由卷曲由72變到73。突變前后UCPl基因RNA二級(jí)結(jié)構(gòu)發(fā)生改變,其最小自由能由-1.934 MJ/mol變?yōu)?1.932 MJ/mol。這說(shuō)明該位點(diǎn)可能影響2個(gè)品種的肌內(nèi)脂肪沉積等肉質(zhì)性狀差異,下一步工作將分析該多態(tài)性位點(diǎn)與兔肌內(nèi)脂肪沉積性狀的關(guān)聯(lián)性,就UCP1基因?qū)ν萌赓|(zhì)性能的調(diào)控作進(jìn)一步探究,從而為線粒體中相關(guān)基因?qū)ν眉?nèi)脂肪沉積影響提供一定的依據(jù)。
