TcLr35小麥中抗病相關(guān)基因S2A2的抗葉銹性分析
2017-05-17 10:05:45張艷俊張家瑞栗小英王海燕劉大群
江蘇農(nóng)業(yè)科學(xué) 2016年1期
張艷俊 張家瑞 栗小英 王海燕 劉大群


摘要:NBS-LRR是已克隆植物抗病基因的高度保守氨基酸區(qū)域。前期工作中,筆者成功克隆獲得了1個(gè)通讀的NBS類抗病同源基因S2A2的cDNA序列,該序列含有NB-ARC保守結(jié)構(gòu)域和多個(gè)LRR結(jié)構(gòu)域,且在小麥葉片中為低豐度組成型表達(dá)。進(jìn)一步根據(jù)S2A2基因在TcLr35與Thatcher中擴(kuò)增獲得的基因序列差異位點(diǎn)設(shè)計(jì)特異性引物,分別以TcLr35、Thatcher為模板進(jìn)行擴(kuò)增,篩選出了具有較高穩(wěn)定性和可重復(fù)性的3對引物。利用3個(gè)多態(tài)性引物對TcLr35、Thatcher及其F2代群體進(jìn)行擴(kuò)增和遺傳性分析,并用Mapmanager軟件計(jì)算分子標(biāo)記與抗葉銹病基因之間的遺傳距離,結(jié)果發(fā)現(xiàn)這3對引物獲得的標(biāo)記與Lr35基因遺傳距離較遠(yuǎn)。利用這3個(gè)多態(tài)性引物擴(kuò)增33個(gè)不同小麥抗葉銹病近等基因系材料,并回收測序,結(jié)果表明該基因序列在不同近等基因系材料中廣泛存在。
關(guān)鍵詞:NBS-LRR;抗病基因;小麥葉銹病;分子標(biāo)記
中圖分類號(hào):S435.12 文獻(xiàn)標(biāo)志碼:A 文章編號(hào):1002—1302(2016)01—0148—04
小麥?zhǔn)鞘澜缟戏植挤秶顝V,栽培面積最大,總產(chǎn)量最高的糧食作物。病害是威脅小麥穩(wěn)產(chǎn)高產(chǎn)的主要因素之一。由專性寄生真菌——小麥葉銹菌(Puccinia triticina)引起的小麥葉銹病是最嚴(yán)重的小麥葉部病害之一,在世界各產(chǎn)麥區(qū)均有發(fā)生。在北非、東南亞、中亞、東歐和南北美洲等廣大地區(qū)較為嚴(yán)重。該病也是威脅我國小麥生產(chǎn)的一種常發(fā)病害,20世紀(jì)70年代,北方麥區(qū)曾發(fā)生3次中度以上流行(1973年、1975年和1979年),給我國小麥生產(chǎn)造成嚴(yán)重的損失。過去幾年中我國雖然沒有葉銹病大規(guī)模暴發(fā)的相關(guān)報(bào)道,但在華北及黃淮麥區(qū),葉銹病經(jīng)常對小麥生產(chǎn)造成危害,近年來該病害呈上升趨勢。開發(fā)利用寄主本身的抗病性是解決病害問題的根本途徑,但由于葉銹菌毒性基因的高度變異性和抗銹品種的不合理使用,導(dǎo)致傳統(tǒng)抗病品種的抗性喪失。因此,利用包括DNA分子標(biāo)記技術(shù)在內(nèi)的多種手段提高抗病基因和抗病類型的豐富度、保持品種抗銹性的持久有效,是一項(xiàng)十分緊迫的任務(wù)。
成株抗葉銹病基因Lr35最初來源于擬斯卑爾脫山羊草(Aegilops spehoides),通過與二倍體的Triticum monococcum回交轉(zhuǎn)到小麥中,定位在2B染色體上,與Sr39緊密連鎖。國內(nèi)外至今尚未發(fā)現(xiàn)對它表現(xiàn)毒性的菌株存在,是一個(gè)應(yīng)用潛力很大的由主效基因控制的抗病基因。目前人們已經(jīng)找到了與Lr35緊密連鎖的BCD260和UBC836這2個(gè)分子標(biāo)記,并分別轉(zhuǎn)化為更為穩(wěn)定的STS和SCAR特異性標(biāo)記。筆者所在實(shí)驗(yàn)室王海燕等在TcLr35小麥中獲得$2A2基因全長,序列比較結(jié)果表明,與Thatcher中獲得的該基因序列差異明顯,含有多個(gè)插入/缺失或者SNP位點(diǎn),為了能快速且恰當(dāng)?shù)脑u價(jià)該基因的功能及其與小麥葉銹病的抗病相關(guān)性,本研究利用特異性引物篩選抗感材料和不同近等基因系材料差異,以期明確S2A2基因抗葉銹性。
1材料與方法
1.1材料
供試小麥葉銹菌致病菌株99-8-11-5-3由河北農(nóng)業(yè)大學(xué)小麥葉銹病研究中心提供。小麥抗葉銹病近等基因系材料TcLr35、感病親本Thatcher和鄭州5389及其他抗葉銹病近等基因系均來自河北農(nóng)業(yè)大學(xué)小麥葉銹病研究室。
1.2特異性引物的篩選
圍繞Thatcher和TcLr35中獲得的序列差異位點(diǎn)設(shè)計(jì)特異性引物(表1),將其自由組合成不同引物組合,并以TcLr35和Thatcher為模板進(jìn)行篩選。引物篩選的24μL反應(yīng)體系為:1.0μL模板,18.7μL ddH2O,2.5μL 10×buffer,0.5μLdNTP,正反向引物各0.5μL,0.3μL Taq酶。反應(yīng)程序?yàn)?4℃預(yù)變性1 min;94℃變性30 s,58℃退火1 min,72℃延伸2 min,30個(gè)循環(huán);最后72℃延伸10 min。擴(kuò)增產(chǎn)物用1.0%瓊脂糖凝膠電泳檢測,用凝膠成像分析系統(tǒng)進(jìn)行拍照及分析。
1.3抗葉銹病分析
Lr35基因?yàn)槌芍昕谷~銹病基因,待供試小麥生長至4葉期,采用撒粉法接種新鮮小麥葉銹菌種。接種后15 d左右,待發(fā)病充分時(shí)進(jìn)行抗感鑒定,按0、;、1、2、3、4等6級(jí)標(biāo)準(zhǔn)調(diào)查記載表現(xiàn)型,0~2級(jí)為抗病,3~4級(jí)為感病。同時(shí)利用Lr35的SCAR引物進(jìn)行抗病性鑒定。24μL反應(yīng)體系為:模板1.0μL,ddH20 18.7μL,10×buffer 2.5μL,dNTF0.5μL,正反向引物各0.5μL,Taq酶0.3μL。反應(yīng)程序?yàn)椋?4℃預(yù)變性1 min;94℃變性30 s,64℃退火1 min,72℃延伸2 min,35個(gè)循環(huán);最后72℃延伸10 min。擴(kuò)增產(chǎn)物用1.0%瓊脂糖凝膠電泳檢測,用凝膠成像分析系統(tǒng)進(jìn)行拍照及分析。
利用小麥近等基因系尋找TcLr35的多態(tài)條帶,再用F2分離后代單株,驗(yàn)證該多態(tài)性條帶與目標(biāo)基因連鎖程度。用Mapmanager軟件對數(shù)據(jù)進(jìn)行連鎖分析,計(jì)算遺傳距離。
2結(jié)果與分析
2.1特異性引物的篩選
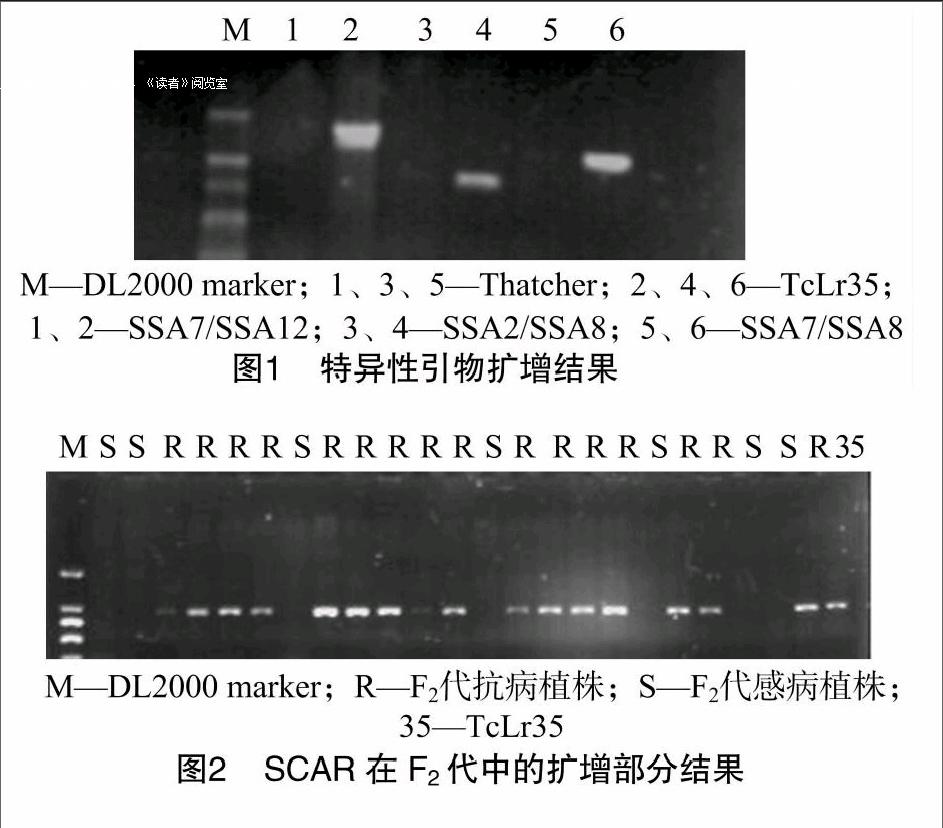
為了進(jìn)一步明確該基因與Lr35的關(guān)系,根據(jù)S2A2基因在TcLr35和Thatcher中序列的差異性位點(diǎn)設(shè)計(jì)引物組合進(jìn)行篩選,獲得3條特異性條帶,即3對引物組合能擴(kuò)增出小麥抗葉銹病近等基因系TcLr35特異性條帶,而在Thatcher中條帶缺失,這3對引物分別是SSA7/SSA8、SSA2/SSA8、SSAT/SSA12。引物對SSA7/SSA8能在TcLr35中穩(wěn)定擴(kuò)增出1條大小為1200 bp左右的條帶;引物對SSA2/SSA8能在TcLr35中穩(wěn)定擴(kuò)增出1條大小為890 bp左右的條帶;引物對SSA7/SSA12能在TcLr35中穩(wěn)定擴(kuò)增出1條大小為1300 bp左右的條帶(圖1)。經(jīng)多次試驗(yàn),擴(kuò)增結(jié)果具有很高的穩(wěn)定性和可重復(fù)性。
2.2小麥葉銹病的鑒定及群體遺傳分析
葉銹菌生理小種99-8-11-5-3接種TcLr35、感病親本Thatcher及雜交F1代10個(gè)單株。抗病鑒定結(jié)果表明,TcLr35和10個(gè)雜交F1代單株對生理小種99-8-11-5-3表現(xiàn)為高抗(反應(yīng)型為0),而Thatcher表現(xiàn)為高感(反應(yīng)型為4)。
成株期對192個(gè)F2代單株接種小麥葉銹菌生理小種99-8-11-5-3后,抗病146株,感病46株,用卡方測驗(yàn)(x2-test)檢驗(yàn)其適合度,其卡方值是0.11,檢驗(yàn)結(jié)果表明,該基因符合3:1的遺傳規(guī)律,說明F2代中的Lr35基因以單顯性基因方式存在。為保證鑒定結(jié)果的可靠性,用Lr35的SCAR引物對192株F2(TcLr35×Thatcher)代群體抗病性進(jìn)行鑒定,結(jié)果在抗病品種中擴(kuò)出了Lr35基因的特異條帶而在感病品種中沒有(圖2),結(jié)果與接菌鑒定一致。
為檢測目的基因位點(diǎn)與所獲得標(biāo)記位點(diǎn)的遺傳連鎖性,利用3個(gè)多態(tài)性引物檢測F2代群體單株,其結(jié)果顯示,引物對SSA7/SSA8在46株感病中有4株擴(kuò)出大小為1200 bp的差異性條帶,而在146株抗病中有4株沒有擴(kuò)出差異性條帶(圖3);引物對SSA7/SSA12在46株感病中有5株擴(kuò)出大小為1290 bp的差異性條帶,而在146株抗病中有3株沒有擴(kuò)出差異性條帶(圖4);引物對SSA2/SSA8在46株感病中由5株擴(kuò)出890 bp差異性條帶,而在146株抗病中有4株沒有擴(kuò)出差異性條帶(圖5),表明在后代群體中3個(gè)標(biāo)記交換值均較高。用Mapmanager軟件計(jì)算分子標(biāo)記與抗葉銹病基因之間的遺傳距離,結(jié)果發(fā)現(xiàn)這3對引物獲得的標(biāo)記與Lr35基因遺傳距離較遠(yuǎn)。
2.3特異性引物擴(kuò)增近等基因系
用3對特異性引物組合分別對33個(gè)小麥抗葉銹基因系材料進(jìn)行PCR擴(kuò)增(圖6、圖7、圖8),結(jié)果顯示,引物組合SSA7/SSA8、SSA7/SSA12除在TcLr14a、TcLrl4b、TcLr14ab、TcLr41、TcLr42和TcLrl8中沒有獲得擴(kuò)增條帶外,其余27個(gè)近等基因系中均擴(kuò)增出目的條帶;引物組合SSA2/SSA8除在TcLr14a、TcLr14b、TcLr14ab、TcLr41、TcLr42、TcLr11和TcLr18中沒有條帶外,其余26個(gè)近等基因系中均擴(kuò)增出目的條帶,說明3個(gè)特異性條帶在大多數(shù)近等基因系材料中存在,選取不同來源近等基因系材料中獲得的差異性條帶回收測序,所得序列與TcLr35的差異性條帶序列差異不大,表明獲得的序列在近等基因系材料中普遍存在。
3討論
抗葉銹病基因的利用,在小麥抗葉銹病育種中發(fā)揮了重要的作用。迄今為止,國際上已發(fā)現(xiàn)100余個(gè)小麥抗葉銹基因,正式命名編號(hào)至Lr72,其中大部分已分子作圖并獲得緊密連鎖或共分離的分子標(biāo)記(Lr1、Lr2c、Lr12、Lr25、Lr32、Lr37、Lr38、Lr41、Lr42、Lr48和LrZH84,但由于氣候條件的變化、生產(chǎn)上的局限性及新的葉銹菌生理小種的形成,導(dǎo)致許多抗銹品種反應(yīng)型由近免疫到了1或2型甚至4型。如Lr1基因?qū)π←湸蟛糠秩~銹菌生理小種已表現(xiàn)感病,Lr2c基因毒性頻率也下降等等。培育成株抗性品種將會(huì)成為控制小麥葉銹病經(jīng)濟(jì)與環(huán)保相結(jié)合的有效方法。小麥抗葉銹病基因Lr35是一個(gè)成株抗性基因,且尚未報(bào)道發(fā)現(xiàn)其毒性小種,是有效的成株抗性基因,吸引了大量的研究者。本實(shí)驗(yàn)室先后對其抗病機(jī)制、分子標(biāo)記、輔助育種等方面進(jìn)行了研究,并取得了一定的進(jìn)展。本研究旨在前期工作的基礎(chǔ)上進(jìn)一步明確S2A2基因與小麥抗葉銹性的關(guān)系。
研究發(fā)現(xiàn)試圖應(yīng)用特異性引物在抗感材料間發(fā)現(xiàn)帶型上的差異不太可能。可能是單堿基的差異,而不是堿基數(shù)目的差異,這是不能用普通的電泳帶型區(qū)分的,這與已克隆的小麥抗葉銹病基因Lr34特點(diǎn)類似。本研究根據(jù)S2A2基因序列設(shè)計(jì)全長引物,在Thatcher和TcLr35基因組中分別獲得該基因擴(kuò)增條帶,進(jìn)行了全長測序,發(fā)現(xiàn)S2A2基因在抗感材料問序列差異明顯,尤其3′端差異顯著,含有多個(gè)較長的插入、缺失序列,而大多數(shù)基因的功能主要決定于3′端。
為了揭示$2A2基因與Lr35相關(guān)性,根據(jù)抗感材料中序列差異明顯區(qū)域,尤其圍繞2個(gè)內(nèi)元設(shè)計(jì)特異性引物,有3個(gè)引物能夠揭示TcLr35和Thatcher抗感親本間差異,而在F2代驗(yàn)證中感病單株也出現(xiàn)了與抗病親本相同的特異條帶,郭楠等和閆紅飛等分別在獲得Lr45、Lr38的分子標(biāo)記時(shí)也具有該現(xiàn)象,可能是由單核苷酸位點(diǎn)突變引起的重組突變或染色體交叉導(dǎo)致的,但其交換值較高,這些標(biāo)記與Lr35基因遺傳距離較遠(yuǎn)。當(dāng)然這些引物擴(kuò)增的僅是S2A2基因全長中的一部分序列,可能為遺傳背景中共同的遺傳信息,由于小麥抗葉銹病基因來源比較復(fù)雜,如Lr19來源于長穗偃麥草(Thinopyrum elongatum),Lr35來源于擬斯比爾脫山羊草(Ae-gilops speltoides),Lr38來源于中間偃麥草(T.interrnedium),Lr41和Lr42基因來源于粗山羊草(T.tauschii)等等,轉(zhuǎn)入普通小麥后,經(jīng)過6代回交,可能會(huì)產(chǎn)生一些共同的遺傳物質(zhì),如果篩選更多的特異性引物,可能會(huì)獲得與Lr35緊密連鎖或共分離的分子標(biāo)記。同時(shí)利用這些特異性引物在一部分小麥抗葉銹病近等基因系材料中可以獲得相同的序列,在一部分小麥抗葉銹病近等基因系材料中不能獲得有效擴(kuò)增片段,說明這些序列在多數(shù)近等基因系基因組中廣泛存在,而且這種差異能在抗感材料間表現(xiàn)。由于小麥基因組的復(fù)雜性,以及Lr35目的基因本身來源于外源導(dǎo)入片段,從以上的研究結(jié)果推測,S2A2基因可能不是Lr35目的基因,但其中的一些序列在多數(shù)小麥抗葉銹病近等基因系材料中普遍存在,而且可能與小麥抗葉銹性相關(guān),可能為多數(shù)小麥抗葉銹病反應(yīng)中的調(diào)控基因,該結(jié)果尚待在穩(wěn)定轉(zhuǎn)化中進(jìn)行進(jìn)一步的功能驗(yàn)證。
