光電子響應微生物的篩選鑒定及生長代謝特征研究
2017-05-19 09:40:01向沙劉明學張格格羅浪魏紅福董發勤
生物技術通報 2017年4期
向沙劉明學,張格格羅浪魏紅福董發勤
(1. 西南科技大學生命科學與工程學院,綿陽621010;2. 固體廢物處理與資源化教育部重點實驗室,綿陽621010)
光電子響應微生物的篩選鑒定及生長代謝特征研究
向沙1劉明學1,2張格格1羅浪1魏紅福2董發勤2
(1. 西南科技大學生命科學與工程學院,綿陽621010;2. 固體廢物處理與資源化教育部重點實驗室,綿陽621010)
篩選可良好響應半導體光電子的微生物菌株,在礦物與微生物交互作用研究中應用前景廣闊。從某鈾尾礦土壤中,利用含有半導體材料TiN營養成分1/2、1/4、1/8、1/10、1/50、1/100和1/1 000的培養基在光照條件下篩選出光電子響應的菌株;通過形態特征、生理生化和分子系統學對菌株進行鑒定,并用紫外-可見吸收光譜和三維熒光光譜測試分析該菌株在光電子作用下生長情況以及蛋白質、腐殖酸等代謝產物的變化。結果顯示,從土壤中篩選出一株對光電子具有較強響應效果的菌株Y-5,經鑒定為糞產堿桿菌(Alcaligenes faecalis)。該菌株在1/10底物濃度中由于營養豐富光電子作用并不明顯,在1/50,1/100較低底物濃度中營養貧乏,光電子作用加速了該菌生長速率,代謝產物蛋白質、腐殖酸也相應增加。從土壤篩選得到的光電子響應微生物菌株糞產堿桿菌,光電子能促進其生長代謝。該方法在定向篩選光電子響應的菌株方面具有一定的可行性。
光電子;糞產堿桿菌;生長代謝;蛋白質;腐殖酸
長期以來,人們將微生物按能量代謝的途徑分為兩大類:光能營養微生物和化能營養微生物,并且認為非光合型的微生物只能利用無機物、無機成因或者生物成因的有機物來獲得能量,不能直接利用太陽光能。但是隨著礦物和微生物交互作用研究的發展,發現了地表廣泛存在的半導體礦物吸收太陽光能后分離的光電子-空穴對,亦可對微生物產生作用。例如,微生物可通過細胞外膜上的一些分子(如血紅素)與鐵錳礦物晶體結構中的變價元素Fe、Mn進行價電子交換以此獲取能量[1-4];半導體礦物光催化作用可以顯著降低紅壤微生物群落多樣性,改變紅壤微生物群落結構[5]。天然閃鋅礦產生的光電子促進嗜酸性氧化亞鐵硫桿菌生長[6];一定范圍內的光電子能量可促進糞產堿桿菌生長[7];半導體礦物CdS光催化產生的光電子能夠作用于從異養產堿桿菌菌體中提取的NAD輔酶,使其還原為NADH[8]。這意味著半導體礦物光電子可以作為部分微生物生長代謝所需的重要能量來源,揭示了一種在自然界中可能廣泛存在的非光合微生物利用太陽能的新途徑[1]。自然界土壤中廣泛存在的還原性物質(腐殖酸、抗壞血酸、還原態硫等)[9-12],能夠捕獲半導體產生光電子的空穴,并擔當電子穿梭體參與微生物在地表過程中的物質循環與能量代謝[13-15]。綜上所述,半導體礦物光生電子作為土壤微生物的外源電子來源之一,理應對土壤微生物的生長代謝行為產生一定的影響[1]。
半導體材料納米TiN作為催化及材料領域的一類重要的半導體化合物[16,17],與TiO2相比,氮摻雜晶格結構中,電荷密度分布改變形成缺陷或改變晶格類型,其能帶結構改變,光催化性能改善[18-20],可見光響應能力強,是一種優良的可見光催化劑[14,15],所以研究采用TiN作為提供光電子的半導體材料。本研究利用半導體材料納米TiN提供光電子從某鈾尾礦區土壤里直接篩選出能夠較強響應半導體礦物光電子的微生物,探討該微生物響應光電子過程中代謝產物的變化規律,旨為闡明微生物與礦物光電子相互作用微觀機制奠定理論基礎。
1 材料與方法
1.1 材料
1.1.1 材料與試劑 土壤樣品采集于某鈾尾礦區。TiN懸液:稱取0.003 g TiN將其倒入600 mL的超純水,超聲振蕩30 min使其均勻分散后,121℃滅菌20 min,保存備用。培養基:酵母粉5.0 g,蛋白胨10.0 g,NaCl 5.0 g,瓊脂20.0 g,抗壞血酸12 g(空穴捕獲劑),蒸餾水1 000 mL,pH7.0-7.2(除NaCl以外的成分按1∶2、1∶4等比例稀釋,配制相應的1/2、1/4培養基)。馬斯亮藍G-250染料試劑:稱取10 mg考馬斯亮藍G-250,加入95%乙醇5 mL和85%磷酸10 mL,定容至100 mL。
1.2 方法
1.2.1 鈉燈照射TiN材料光電子性能分析 取5 mg/L的50 mL TiN懸液混勻至石英電解池,插入鈦片工作電極、飽和甘汞參比電極、鉑絲對電極,在鈉燈光照100 s、無光照100 s、光照200 s和無光照200 s交替作用條件下,設置電壓0 V測定樣品電流。
1.2.2 光電子響應微生物的篩選 分別取上述土壤樣品5 g,加入100 mL濃度為5 mg/L TiN的1/2液體培養基,放在鈉燈照射下的振蕩培養箱培養2-d(條件:轉速120 r/min,溫度30℃),取1 mL培養液至新鮮的濃度為5 mg/L TiN的1/4液體培養基,鈉燈照射下培養2-3 d。取0.5 mL涂布在1/8固體培養基(TiN濃度為5 mg/L),鈉燈照射下培養2-3 d。接種到1/50固體培養基(TiN濃度為5 mg/L)反復劃線分離純化,得到較強響應光電子的Y-5菌株,接種在1/100和1/1 000固體培養基平板(TiN濃度分別為5 mg/L和0 mg/L),并觀察菌落生長狀況。
1.2.3 菌株Y-5的鑒定
1.2.3.1 形態學鑒定 將篩選到的菌株劃線接種于固體培養基平板上,30℃培養2-3 d,觀察菌落形態;取穩定期菌夜涂布至蓋玻片,自然風干,用2.5%戊二醛固定8 h后倒掉戊二醛,分別用30%、50%、70%、90%和100%乙醇梯度洗脫(每次15-20 min),自然風干后電子顯微鏡掃描菌體形態。參照《伯杰氏細菌鑒定手冊(第9版)》[21]與《常見細菌系統鑒定手冊》[22]對獲得的菌株進行革蘭氏染色、氧化酶等生理生化實驗。確定菌株類型。
1.2.3.2 系統發育學分析 用生工生物工程股份有限公司提供的Ezup柱式細菌基因組DNA抽提試劑盒提取出該菌株的DNA,以7F 1540R和27F 1492R為引物,利用PCR技術擴增出16S r DNA區段,并將擴增片段用1%的瓊脂糖凝膠進行電泳[23],純化PCR產物在上海生工生物工程有限公司測序,通過Blast 比對分析所得序列與GenBank 數據庫中已有的細菌 16S rRNA 序列的相似性,運用MEGA 5.1 軟件構建系統發育樹。
1.2.4 菌株Y-5生長對光電子的響應 分別配制5 mg/L TiN的1/10、1/50、1/100液體培養基(按照每個濃度設置光照組和錫箔紙包裹的黑暗組即無光電子組),然后加入穩定期的菌種1 mL,放在鈉燈照射下的振蕩培養箱培養(條件:轉速180 r/min,溫度30℃),利用紫外-可見分光光度計在0 h、6 h、10 h、12 h、16 h、24 h、36 h、48 h、60 h和72 h時段取菌液3 mL于波長600 nm下測OD值,每組處理設置3個重復。收集培養72 h的菌體,生理鹽水清洗菌體后,置于0.2 mol/L NaOH中沸水浴10 min,得到微生物蛋白液,取待測蛋白液用0.9%生理鹽水按一定比例稀釋至2.4 mL,加入0.6 mL G-250(100 mg/L)于比色管中混合搖勻。使用紫外分光光度計于峰位595 nm,以生理鹽水作參比,測定吸光度,通過制作標準曲線,計算總蛋白的量[6,24,25]。
建設項目壓覆礦產資源的評估還應進行可否壓覆的評價。評估的重點,是以被壓覆資源近幾年平均價格水平上的經濟價值,與該工程項目產生的社會、經濟、生態效益進行對比,評估壓覆礦產資源的利弊,并給出建設項目是否值得壓覆的建議,為行政主管部門批復項目工程提供依據。
1.2.5 菌株Y-5代謝對光電子響應 按照1.2.4方法和條件將培養到穩定期的菌液在4℃、10 000 r/min條件下冷凍離心,收集上清液,利用F-7000熒光光度計進行三維熒光分析。儀器參數:激發和發射波長是從200 nm開始掃描,到800 nm結束;掃描速度為12 000 nm/min;狹縫10 nm,步徑10 nm,PMT電壓為700 V[26]。
2 結果
2.1 鈉燈照射TiN材料光電子性能分析
為驗證半導體材料TiN產生光電子性能,測定了TiN在鈉燈照射下恒電位時間-電流(I-t)曲線。結果(圖1)表明在有光照時TiN懸液電流值大約在1.0 uA,在無光照時電流值大約在0 uA,光照時段電流大于無光照時段電流。說明該濃度TiN,在鈉燈下可以產生一定量的光電子。
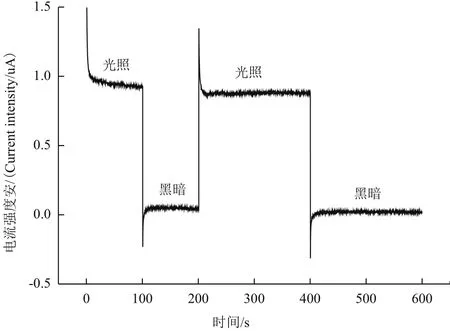
圖1 TiN恒電位I-t曲線
2.2 光電子響應微生物的篩選
經初步富集、篩選分離得到光電子響應微生物Y-5菌株。如圖2所示,菌株在5 mg/L TiN的1/100固體培養基上出現生長現象,在無TiN的1/100固體培養基和5 mg/L TiN的1/1 000固體培養基上無生長現象,光電子對該菌的生長產生了一定的促進作用。
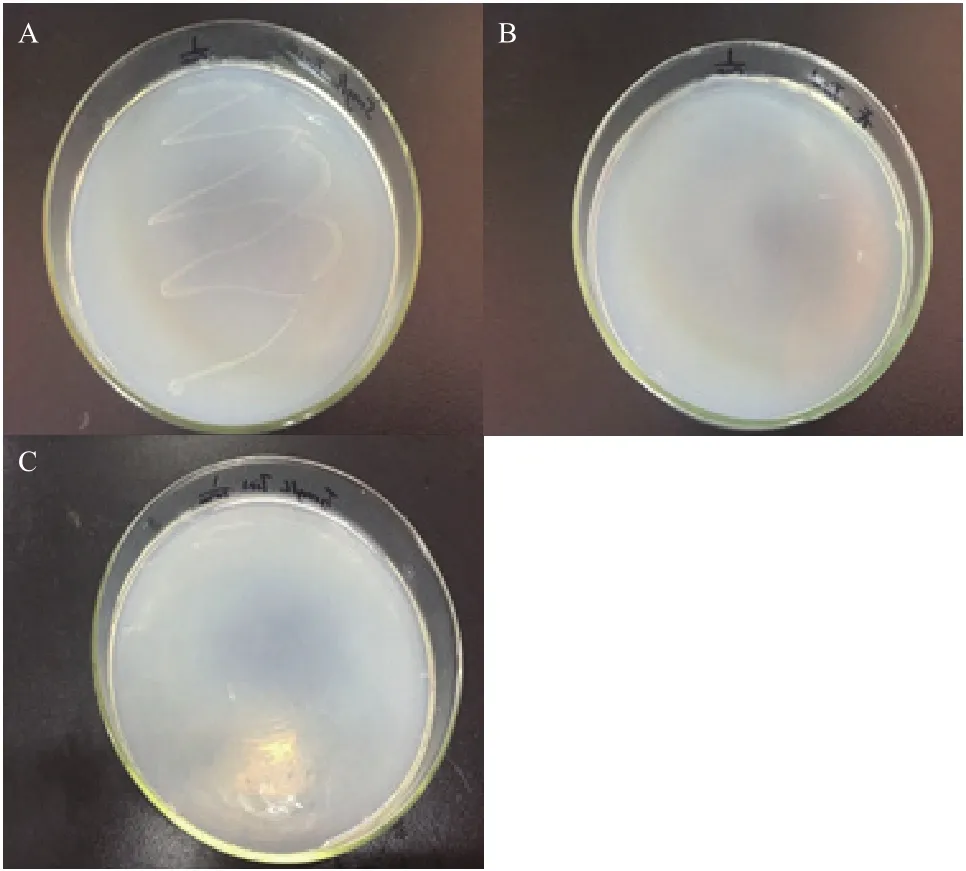
圖2 光電子響應微生物篩選結果
2.3 菌株的鑒定
2.3.1 菌株的形態學觀察 在完全營養成分的固體培養基上,Y-5菌株顏色為半透明到不透明,形態為扁平或略凸起,光滑(有時為模糊或粗糙)和邊緣完整,大多數菌株可蔓延成不規則邊緣(圖3,圖4)。通過掃描電鏡發現菌株大小約為0.7-2 μm,為桿狀。菌株Y-5常規生理生化特征分析結果見表1,結果表明菌株Y-5為革蘭氏陰性細菌,不能利用糖類物質,可以利用檸檬酸鹽,在肉湯培養基中pH值上升到8.6左右,與產堿桿菌屬糞產堿桿菌相似。
圖3 Y-5菌落形態

圖4 Y-5 SEM圖像
2.3.2 系統發育學分析 將菌株Y-5的16S rRNA序列(擴增長度為1 415 bp),通過 GenBank 進行序列分析和同源性比對。結果(圖5)表明,菌株Y-5序列與糞產堿桿菌(Alcaligenes faecalis)的16S rRNA 序列同源性為98%。
2.4 糞產堿桿菌生長對光電子的響應
為了突出不同條件下實驗效果差別,給細菌一定的生長壓力,在生長代謝實驗中分別采用稀釋10倍、50倍和100倍的培養基培養。如圖6所示,分別為菌株Y-5在5 mg/L TiN的1/10、1/50和1/100液體培養基的生長曲線,在接種培養初期(即最初約5 h),細菌生長處于遲緩期,光照實驗組與黑暗對照組無差別;5 h后,開始進入對數生長期,細菌大量繁殖,細胞濃度大幅增加。在1/10培養基中,對數期光照實驗組細胞濃度與黑暗對照組幾乎無差別,在1/50和1/100培養基中生長速率和生長數量均大于相應濃度的黑暗對照組,表明光催化提高了細菌生長速率和數量。隨后細菌生長進入衰亡期,在1/10培養基中,光照實驗組衰亡速率與黑暗對照組衰亡速率差別不明顯;在1/50和1/100培養基中光照實驗組衰亡速率明顯小于黑暗對照組,表明光催化延長了細菌的衰亡時間。定量蛋白測試結果(圖7)顯示1/10、1/50和1/100培養基有光電子菌體總蛋白含量大于無光電子對照組綜上所述1/50和1/100液體培養基中TiN產生的光電子促進了菌株Y-5生長。

表1 菌株Y-5的生理生化特征
2.5 糞產堿桿菌光電子響應中代謝產物的變化
通過三維熒光技術測定有無光電子作用下Y-5菌株的代謝產物變化。如圖8所示,熒光圖譜主要在激發波長200-400 nm和發射波長200-500 nm范圍內,該區域主要在蛋白質熒光峰(激發波長位于260-290 nm,發射波長發射波長位于340-380 nm)和腐殖酸熒光峰(激發波長位于320-390 nm,發射波長位于400-480 nm)。讀取該區域內蛋白質熒光峰值(1/10、1/50和1/100培養基光照組和黑暗組分別是4 029和3 987、2 689和2 574、1 241和1 130)和腐殖酸熒光峰值(1/10、1/50和1/100培養基光照組和黑暗組分別是2 792和2 779、893和841、453和402)表明Y-5菌株在5 mg/L TiN的1/10、1/50和1/100培養基中光照實驗組的蛋白質熒光峰和腐殖酸熒光峰變化較明顯。
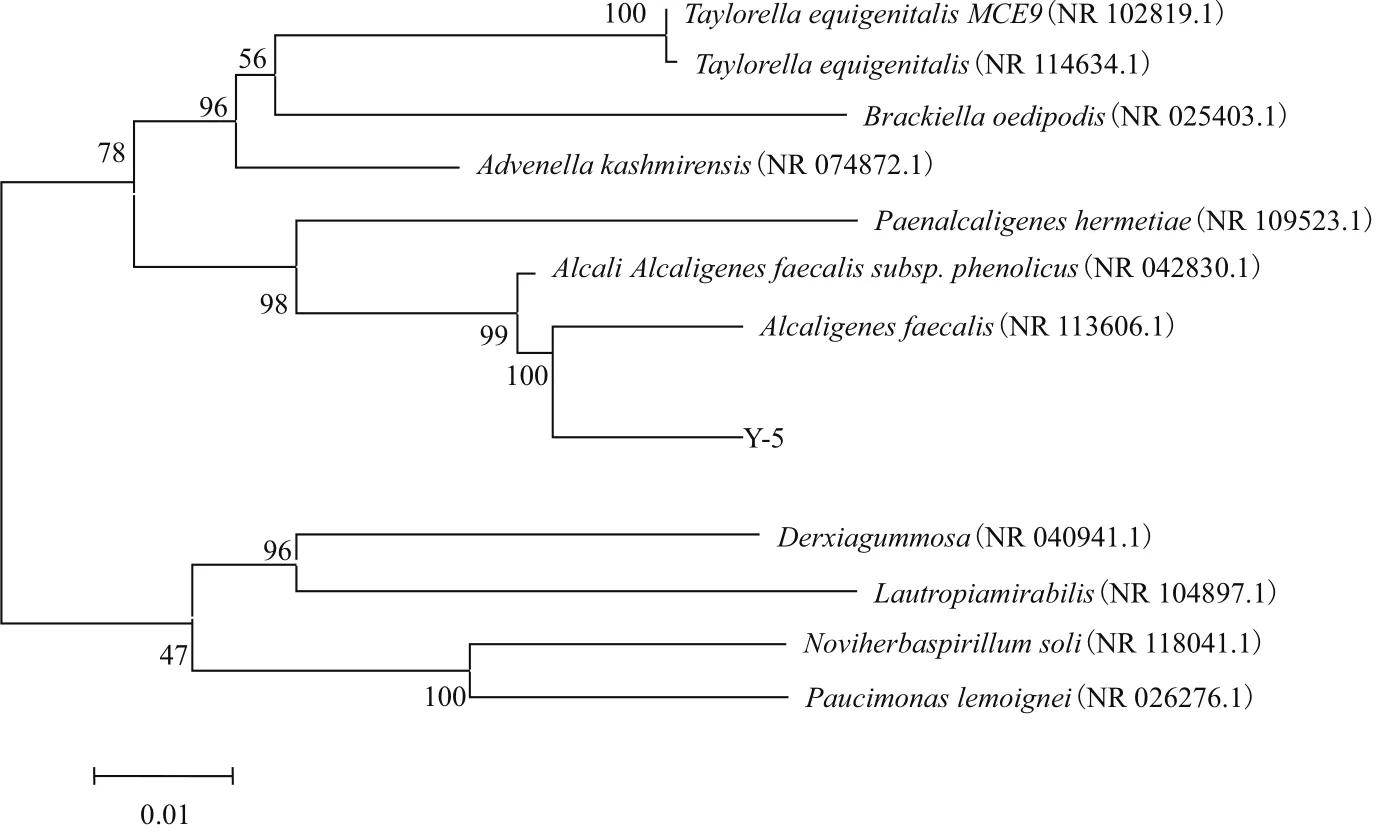
圖5 Y-5菌株和相似屬的一些種基于16S rDNA序列構建的系統發育樹
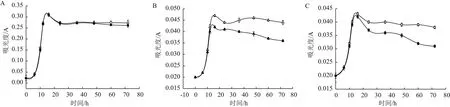
圖6 Y-5菌株生長曲線圖
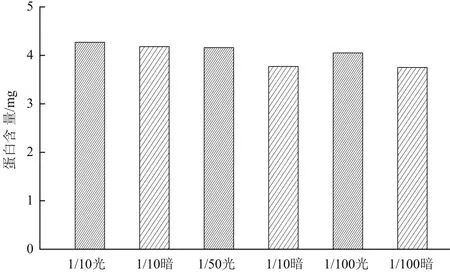
圖7 Y-5菌株與5 mg/L TiN的1/10、1/50、1/100培養基作用后總蛋白測定結果
分析蛋白質和腐殖酸結果如圖9所示,Y-5菌株在1/10培養基中的蛋白質和腐殖酸熒光峰光照實驗組與黑暗對照組差別較小,在1/50和1/100培養基中蛋白質和腐殖酸熒光峰光照實驗組與黑暗對照組差別較大,即在該條件下光照實驗組蛋白質和腐殖酸含量大于黑暗對照組。說明在1/50和1/100培養基中光電子作用下Y-5菌株代謝加快。結合生長曲線分析,Y-5菌株在低濃度底物下利用光電子效率更高。
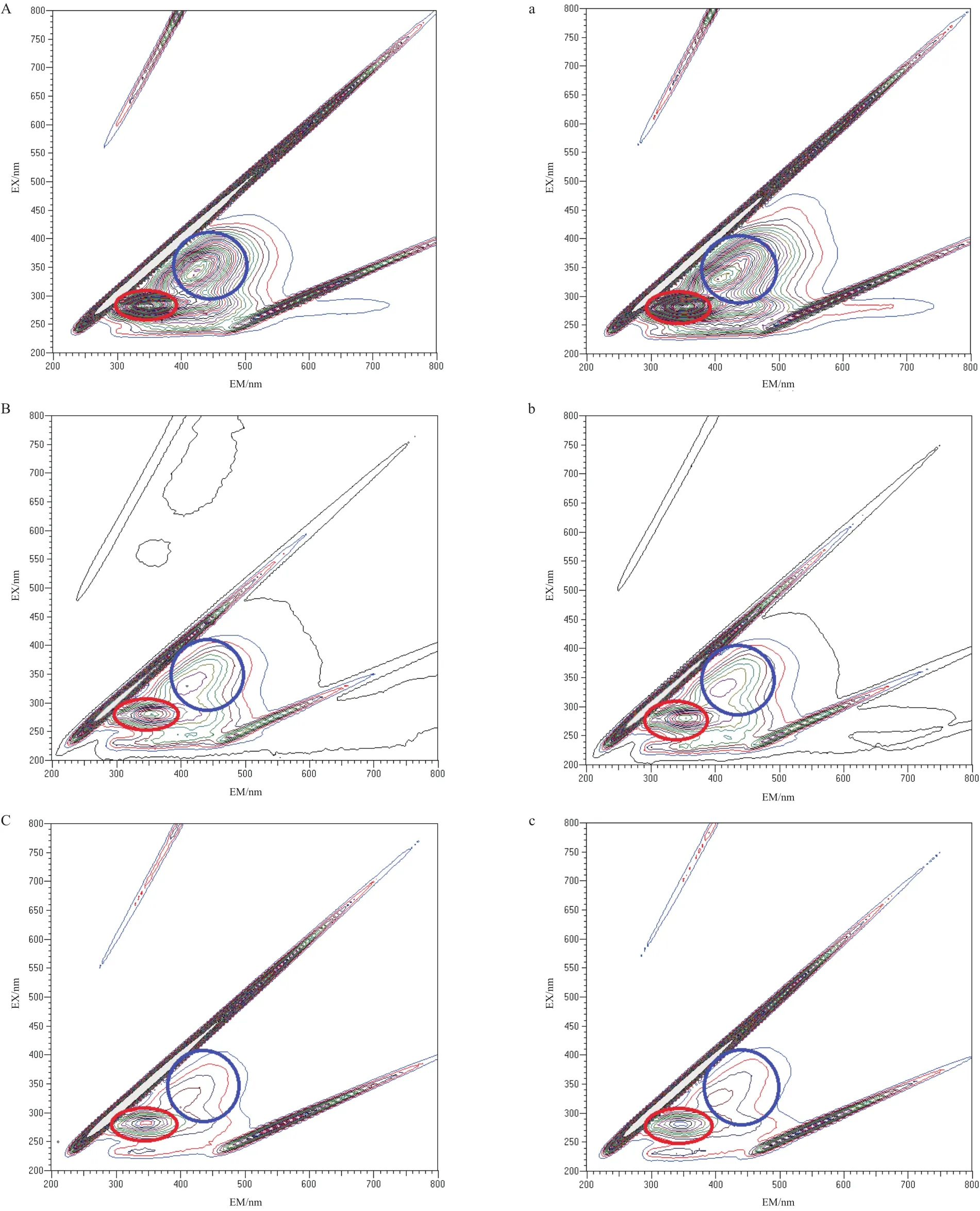
圖8 三維熒光圖譜
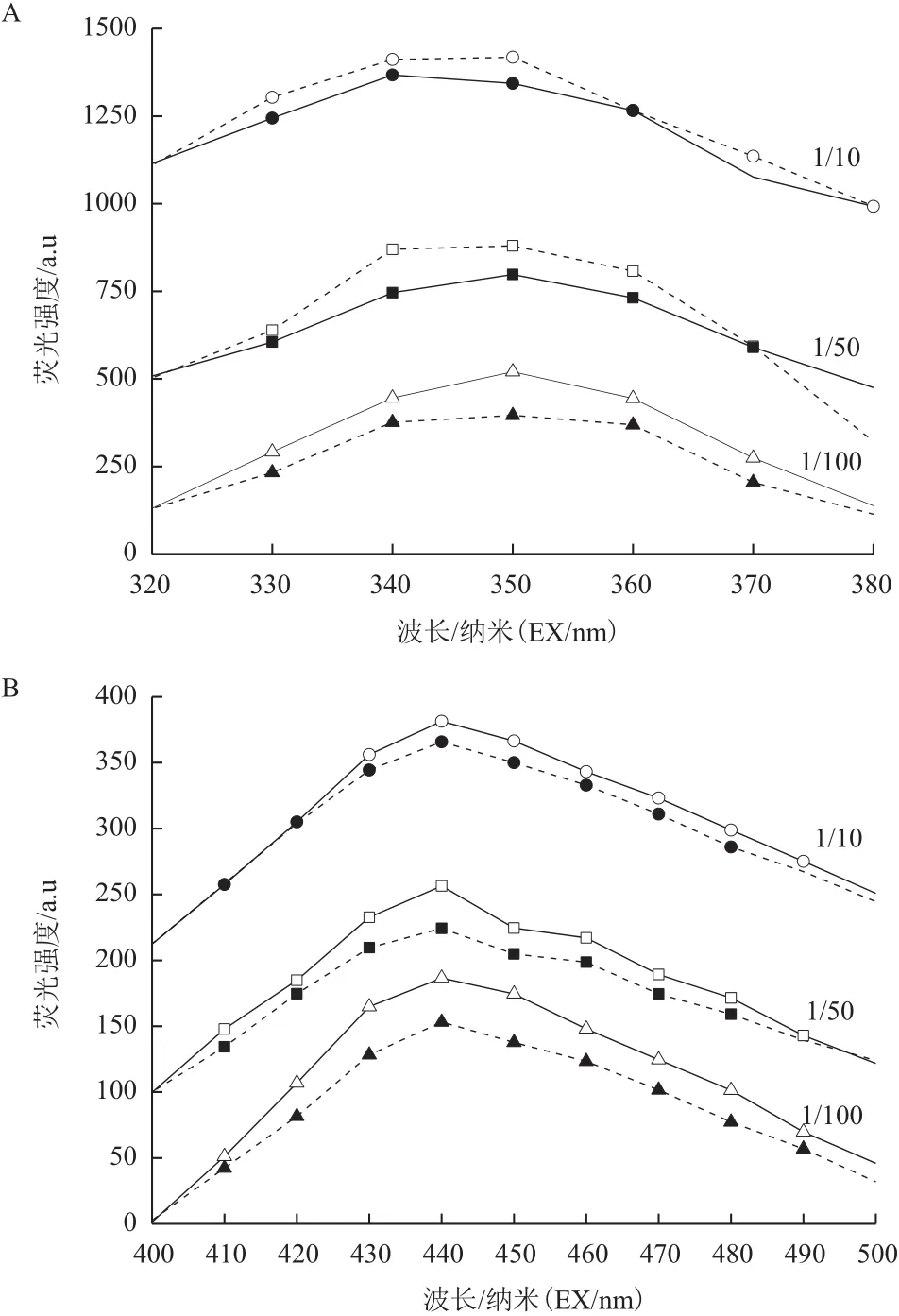
圖9 Y-5菌株與5 mg/L TiN的1/10、1/50、1/100培養基作用后蛋白質(A)和腐殖酸(B)的熒光強度變化分析圖
3 討論
土壤里的微生物是土壤生態系統結構和維持生態系統穩定性的重要構成[27],不僅可以代謝土壤中的動植物殘體、調節土壤有機質、土壤結構形成[28],而且與無機礦物的交互作用影響著地球表層物質循環與環境演化,包括礦物的形成與演化、養分循環與污染物的環境行為等[29-32]。隨著礦物與微生物交互作用研究領域的發展,微生物與礦物之間電子轉移和能量流動逐漸受到關注。近年來,國內外關于這方面研究也取得一些進展,不僅發現一些微生物可以利用光電子為自身提供能量[1],還探索了微生物利用光電子途徑受到光波長(光子能量)與光強(光子數量)兩方面的因素調控、氧化亞鐵硫桿菌利用光電子的機理以及利用效率等[2,33]。
大量研究表明[1,34],半導體光電子能夠延長鐵細菌的電子傳遞鏈,作為新陳代謝的能量,實現光能-電能-化學能-生物能的轉換,但轉換效率遠遠低于植物轉換效率的10%;在外加電壓下光電子作用使氧化亞鐵硫桿菌細胞濃度由2.5×105/mL上升到到3.25×106/mL,增加12倍。曾翠萍等[35]加偏壓證實外來電流可促進糞產堿桿菌菌體在電極表面上的生長和附著;魯安懷等[2]利用光燃料電子雙室裝置體系模擬日光下礦物光電子證實光電子降低土壤里微生物群落多樣性,影響微生物群落結構。本研究利用半導體材料光生電子與土壤里微生物直接作用,篩選獲得Y-5菌株,經形態生理生化系統發育學鑒定為糞產堿桿菌。底物濃度的限制改變了微生物對底物利用效率[36],該菌在1/10底物濃度中光電子作用不明顯,在1/50,1/100較低底物濃度中光電子作用使總蛋白含量增加,菌體生長加速。說明糞產堿桿菌是土壤微生物中光電子響應較強的微生物,能夠利用光電子作為自身能量來源。
生物地球化學的觀點認為,腐殖質能夠擔當電子穿梭體的角色,參與微生物在地表過程中的物質循環與能量代謝[37-40];微生物能夠分泌腐殖質物質進行電子穿梭,參與能量代謝[41];余萍等[7]通過模擬天然半導體礦物所產生的光生電子證實,該模擬光電子作用下,糞產堿桿菌形成了大量具有電化學活性的代謝產物。本研究結果表明,光電子作用下糞產堿桿菌代謝產物蛋白質和腐殖酸增加較多,說明光電子作用下該菌生長代謝加快,增加的腐殖質可能參與電子傳遞。在后續的研究中將利用代謝組學手段對代謝產物進行分析,結合蛋白組學和轉錄組學[42]研究半導體礦物光電子在礦物和糞產堿桿菌活動界面上的傳遞過程與其生長代謝的微觀機制[43]。
4 結論
本研究主要利用含半導體材料TiN的1/2、1/4、1/8、1/10、1/50和1/100培養基分離純化得到一株對光電子較強響應的Y-5菌株,依據形態特征、生理生化和16S rRNA序列系統發育學分析,鑒定其為產堿桿菌屬的糞產堿桿菌(Alcaligenes faecalis)。菌株在不同濃度底物條件下,生長曲線不同;其不僅能夠在5 mg/LTiN光催化下的1/50,1/100低營養成分的培養基中穩定生長,且生長速率和代謝速率也相應的加快,表明該菌株具有響應光電子的能力,是一株“光電能微生物”。同時表明該篩選光電子響應微生物的方法具有可行性,可應用于進一步發現和豐富“光電能微生物”資源方面的研究。
[1]魯安懷, 李艷, 王鑫, 等. 半導體礦物介導非光合微生物利用光電子新途徑[J]. 微生物學通報, 2013, 40(1):190-202.
[2]Lu A, Li Y, Jin S, et al. Growth of non-phototrophic micro-organisms using solar energy through minernal photocatalysis[J]. Nature Communication, 2012, 3(4):768-775.
[3]Marsili E, Baron DB, Shikhare ID, et al. Shewanella secretes flavins that mediate extracellular electron transfer[J]. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(10):3968-3973.
[4]王項. 微生物表征水體污染狀況及還原解毒高鐵離子的應用初探[D]. 長沙:中南大學, 2012.
[5]Zeng CP, Lu A, Li Y, et al. Response of microbial community to sunlight catalysis of semiconductor minerals in red soil[J]. Geological Journal of China Universities, 2011, 17(1):101-106.
[6]王鑫, 李艷, 魯安懷, 等. 不同光波長下天然閃鋅礦光催化作用對Acidithiobacillus ferrooxidans生長的影響[J]. 礦物學報, 2012(s1):175-176.
[7]余萍, 李艷, 魯安懷, 等. 光電子作用下土壤微生物糞產堿桿菌反硝化性能研究[J]. 巖石礦物學雜志, 2013, 32(6):761-766.
[8]Shumilin IA, Nikandrov VV, Popov VO. Photogeneration of NADH under coupled action of CdS semiconductor and hydrogenase from Alcaligenes eutrophus, without exogenous mediators[J]. Febs Letters, 1992, 306(2-3):125-128.
[9]Smirnoff N, Wheeler GL. Ascorbic acid in plants:biosynthesis and function[J]. Critical Reviews in Biochemistry and Molecular Biology, 2004, 35(4):291-314.
[10]Hayase K, Tsubota H. Sedimentary humic acid and fulvic acid as surface active substances[J]. Geochimica et Cosmochimica Acta, 1983, 47(5):947-952.
[11]楊凌, 李松. 水稻土中異化鐵還原對Cr(Ⅵ)還原的環境化學效應[D]. 楊凌:西北農林科技大學, 2006.
[12]劉鄧. 不同厭氧微生物功能群對粘土礦物結構Fe(Ⅲ)的還原作用及其礦物轉變[D]. 武漢:中國地質大學, 2012.
[13]Lovley DR, Coates JD, Blunt-Harris EL, et al Humic substances as electron acceptors for microbial respiration[J]. Nature, 1996, 382(6590):445-448.
[14]Klüpfel L, Piepenbrock A, Kappler A, et al. Humic substances as fully regenerable electron acceptors in recurrently anoxic environments[J]. Nature Geoscience, 2014, 7(3):195-200.
[15]張亞萍. 腐殖質對U(Ⅵ)的吸附與腐殖質/腐敗希瓦氏菌還原U(Ⅵ)的試驗研究[D]. 衡陽:南華大學, 2012.
[16]王佳. 水熱法制備氧化鈦納米棒陣列結構及其光電性能研究[D]. 杭州:浙江大學, 2014.
[17]尹詩衡, 雷淑梅, 匡同春. 非金屬摻雜改性二氧化鈦光催化劑研究進展[J]. 廣東化工, 2007, 34(5):37-39.
[18]尹佳音, 李艷紅, 周婉媛, 等. 納米TiN制備可見光活性TiO2光催化劑的研究[J]. 環境科學與技術, 2014(s1):104-107.
[19]Randorn C, Irvine JTS. Synthesis and visible light photoactivity of a high temperature stable yellow TiO2photocatalyst[J]. Journal of Materials Chemistry, 2010, 20(39):8700 -8704.
[20]王金淑, 刑朋飛, 李莉莉, 等. 機械化學法N 摻雜納米TiO2的制備與表征[J]. 北京工業大學學報, 2006, 32(7):633-637.
[21] 布坎南RE, 吉本斯NE. 伯杰細菌鑒定手冊[M]. 第8版. 北京:科學出版社, 1984.
[22]東秀珠, 蔡妙英. 常見細菌系統鑒定手冊[M]. 北京:科學出版社, 2001.
[23]陳士華, 孫強, 孫莉云, 等. 一株高產油脂微生物菌種的篩選與鑒定[J]. 河南工業大學學報:自然科學版, 2013, 34(6):65-68.
[24]丁健. 鐵還原菌Sphingomonas sp. DJ的分離及其降解特性的研究[D]. 大連:大連理工大學, 2014.
[25]陳聰聰. 金屬還原地桿菌對偶氮染料的還原研究[D]. 大連:大連理工大學, 2012.
[26]傅平青, 劉叢強, 吳豐昌. 溶解有機質的三維熒光光譜特征研究[J]. 光譜學與光譜分析, 2005, 25(12):2024-2028.
[27]滕應, 黃昌勇. 重金屬污染土壤的微生物生態效應及修復研究進展[J]. 土壤與環境, 2002, 11(1):85-89.
[28]趙李寧. 污染土壤生態修復技術研究[J]. 資源節約與環保, 2015, 4:149-150.
[29]O’Loughlin EJ, Gorski CA, Scherer MM, et al. Effects of oxyanions, natural organic matter, and bacterial cell numbers on the bioreduction of lepidocrocite(gamma-Fe OOH)and the formation of secondary mineralization products[J]. Environmental Science & Technology, 2010, 44(12):4570-4576.
[30]Lovley DR. Dissimilatory metal reduction. [J]Annual Review of Microbiology, 1993, 47(3):263-290.
[31]O’Loughlin EJ, Boyanov MI, Flynn TM, et al. Effects of bound phosphate on the bioreduction of lepidocrocite(γ-Fe OOH)and maghemite(γ-Fe2O3)and formation of secondary minerals[J]. Environmental Science & Technology, 2013, 47(16):9157-9166.
[32] 陳銘. Fe(Ⅲ)/腐殖質還原菌的分離鑒定及其還原特性研究[D]. 長沙:湖南農業大學, 2013.
[33]呂明, 魯安懷, 郝瑞霞, 等. 鐵細菌利用天然金紅石光生電子能量研究[J]. 巖石礦物學雜志, 2008, 27(3):212-220.
[34]顏云花, 李艷, 魯安懷, 等. 天然褐鐵礦的光電化學響應及對嗜酸性氧化亞鐵硫桿菌生長的影響[J]. 巖石礦物學雜志, 2009, 28(6):535-540.
[35]Zeng CP, Li Y, Lu A, et al Electrochemical interaction of a heterotrophic bacteria Alcaligenes faecalis, with a graphite cathode[J]. Geomicrobiology, 2012, 29(3):244-249.
[36]Zheng Z Y, Jin W L, Zhan X B, et al. Effect of metabolic structures and energy requirements on curdlan production by Alcaligenes faecalis[J]. Biotechnology and Bioprocess Engineering, 2007, 12(4):359-365.
[37]Lovley DR, Coates JD, Blunt-Harris EL, et al. Humic substances as electron acceptors for microbial respiration[J]. Nature, 1996, 382(6590):445-448.
[38]Klüpfel L, Piepenbrock A, Kappler A, et al. Humic substances as fully regenerable electron acceptors in recurrently anoxic environments[J]. Nature Geoscience, 2014, 7(3):195-200.
[39]Lovley DR, Fraga JL, Blunt-Harris EL, et al. Humic substances as a mediator for microbially catalyzed metal reduction[J]. Acta Hydrochimica et Hydrobiologica, 1998, 26(3):152-157.
[40]王秀娟. 腐殖質類物質對Se/Te及偶氮染料的介導還原研究[D]. 大連:大連理工大學, 2011.
[41]Nevin KP, Lovley DR. Mechanisms for accessing insoluble Fe(III)oxide during dissimilatory Fe(III)reduction by Geothrix fermentans[J]. Applied and Environmental Microbiology, 2002, 68(5):2294-2299.
[42]吳云當, 李芳柏, 劉同旭, 等. 土壤微生物—腐殖質—礦物間的胞外電子傳遞機制研究進展[J]. 土壤學報, 2016, 53(2):278-291.
[43]史其峰. 微生物多糖熱凝膠在CSTR反應器中的發酵與代謝特性研究[D]. 無錫:江南大學, 2008.
(責任編輯 狄艷紅)
Screening of Photoelectron-response Microbes as well as Their Growth and Metabolism
XIANG Sha1LIU Ming-xue1,2ZHANG Ge-ge1LUO Lang1WEI Hong-fu2DONG Fa-qin2
(1. Life Science and Engineering College,Southwest University of Science and Technology,Mianyang 621010;2. Key Laboratory of Waste Solid Treatment and Resource Recycle of Ministry of Education,Mianyang 621010)
Screening microbial strains that have promising response to semiconductor photoelectron brings broad application prospects in the study of interaction between minerals and microorganisms. Using the uranium tailing area soil,the microbial strains with photoelectronresponse were screened in the media of containing 1/2,1/4,1/8,1/10,1/50,1/100,and 1/1 000 semiconductor material TiN under illumination condition. Then the strain was identified through its morphological,physiological and biochemical characteristics,and the molecular phylogeny. Ultraviolet-visible absorption spectra and three-dimensional fluorescence spectra were used to analyze the growth of the strain and the changes of protein,humic acid,and other metabolites. The strain Y-5 screened from soil was identified to be Alcaligenes faecalis,which had strong response to photoelectron of TiN. The optoelectronic effect was not obvious due to rich nutrient in the 1/10 substrate concentration;but in 1/50 and 1/100 low substrate concentration,the growth rate was accelerated by photoelectron due to poor nutrient,and the protein and humic acid increased correspondingly. A photoelectron-response microbial strain A. faecalis was screened from an uranium tailing area soil,and the photoelectron promoted its growth and metabolism. This method demonstrated certain feasibility in screening the photoelectron-response.
photoelectron;Alcaligenes faecalis;growth and metabolism;protein;humic acid
10.13560/j.cnki.biotech.bull.1985.2017.04.027
2016-10-31
國家重點基礎研究發展計劃(“973”)項目(2014CB846003),國家自然科學基金項目(41272371)
向沙,女,碩士研究生,研究方向:環境微生物;E-mail:583567642@qq.com
劉明學,男,教授,研究方向:環境微生物;E-mail:dragonlmx@126.com
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
豬業科學(2018年4期)2018-05-19 02:04:38
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
浙江農業科學(2016年11期)2016-05-04 04:16:49
化工進展(2015年6期)2015-11-13 00:27:33
無機化學學報(2014年9期)2014-02-28 17:32:57
