鉛鋅礦區(qū)耐砷細(xì)菌的分離、鑒定及性質(zhì)研究
2017-05-19 07:51:56吳丹張志鵬馬玉超
生物技術(shù)通報(bào) 2017年5期
吳丹 張志鵬 馬玉超
(北京林業(yè)大學(xué)生物科學(xué)與技術(shù)學(xué)院,北京 100083)
鉛鋅礦區(qū)耐砷細(xì)菌的分離、鑒定及性質(zhì)研究
吳丹 張志鵬 馬玉超
(北京林業(yè)大學(xué)生物科學(xué)與技術(shù)學(xué)院,北京 100083)
旨在從湖南康家灣鉛鋅礦區(qū)的重金屬污染土壤樣品中篩選出耐高濃度砷的細(xì)菌菌株。用稀釋涂布法分離耐砷細(xì)菌;根據(jù)16S rRNA基因系統(tǒng)發(fā)育分析鑒定分離得到的砷高耐受性菌株并檢測菌株內(nèi)含有的砷耐受性相關(guān)基因;用砷鉬藍(lán)法測定耐砷細(xì)菌的砷氧化還原能力;并通過吲哚乙酸(IAA)定量實(shí)驗(yàn)檢測優(yōu)勢菌株產(chǎn)IAA的能力。結(jié)果顯示,從土壤樣品中共分離出152株耐砷細(xì)菌,其中6株細(xì)菌對As5+和As3+的耐受性值分別高達(dá)800 mmol/L和20 mmol/L;并且這6株耐砷細(xì)菌分屬于5個不同的屬:假單胞菌屬、蒼白桿菌屬、芽孢桿菌屬、威廉氏菌屬和節(jié)細(xì)菌屬;菌株Tw31、Tw133、Sw149和Tw222中存在砷還原酶基因arsC,Bw218和Tw222中存在砷離子外排基因arsB/ACR3(2);在72 h之內(nèi),菌株Tw133和Tw222的As3+氧化率(約17 %)和As5+還原率(約35 %)均高于其他菌株;尤其是菌株Tw133在144 h具有48.66 %的As5+還原率;且這兩株菌分別能產(chǎn)生42.86 μg/mL和24.36 μg/mL的IAA。篩選出的Tw133和Tw222菌株在砷耐受性、砷氧還能力和產(chǎn)IAA能力等方面展現(xiàn)出了較明顯的優(yōu)勢,為深入研究細(xì)菌的砷耐受性機(jī)制提供了實(shí)驗(yàn)材料。
耐砷細(xì)菌;耐砷基因;砷氧化還原;IAA
砷(As)廣泛存在于巖石圈、水圈和生物圈。長期接觸砷可以導(dǎo)致各種癌癥的發(fā)生,如膀胱癌、肺癌、皮膚癌等。目前,世界上包括孟加拉、印度、阿根廷、美國和中國等許多國家面臨著嚴(yán)重的砷污染威脅。有報(bào)道稱中國土壤中砷濃度的平均值為11.2 mg/kg,約為世界平均值(6 mg/kg)的2倍[1]。因此砷污染的治理迫在眉睫。
自然環(huán)境中的砷主要以As5+和As3+兩種無機(jī)價態(tài)形式存在,其中后者毒性大約為前者的100倍[2]。砷酸鹽的毒性源于其作為磷酸鹽的結(jié)構(gòu)類似物,可以取代磷酸鹽,抑制氧化磷酸化過程;而亞砷酸鹽能夠與半胱氨酸的巰基結(jié)合,破壞或干擾蛋白和酶的功能[3],從而發(fā)揮毒性作用。目前砷污染的主要治理原理是通過物理、化學(xué)、生物等技術(shù)降低砷的毒性或增加其穩(wěn)定性。研究發(fā)現(xiàn)在砷污染較嚴(yán)重的地區(qū),一些特殊的“耐砷微生物”[4]在砷化合態(tài)的相互轉(zhuǎn)化過程中發(fā)揮著重要作用[5]。目前報(bào)道較多的耐砷微生物有芽孢桿菌屬、假單胞菌屬、紅球菌屬、蒼白桿菌屬、節(jié)細(xì)菌屬、放線菌屬、微桿菌屬等[6-9]。微生物的耐砷特性與砷吸收、氧化還原、甲基化和外排等過程密切相關(guān)[10]。微生物體內(nèi)主要通過ars操縱子介導(dǎo)砷代謝過程,其中砷酸鹽還原酶(ArsC)[11]催化As5+還原成As3+,而As3+又被亞砷酸鹽外排泵(ArsB、ACR3)[12]排出細(xì)胞外,還有其他的一些相關(guān)基因arsR和arsD(調(diào)節(jié)基因),arsA(ATP酶基因),arsH[13]、arsO等也參與了輔助代謝過程。另外一種機(jī)制是由亞砷酸鹽氧化酶AioBA[11]催化介導(dǎo)的,它可以將高毒的As3+氧化成低毒、較穩(wěn)定的As5+。而Qin等[14]的研究發(fā)現(xiàn),在細(xì)菌Rhodopseudomonas palustris CGA009中As3+可以被轉(zhuǎn)甲基化酶ArsM經(jīng)過不同程度的甲基化,最終轉(zhuǎn)變成氣態(tài)的三甲基砷排出細(xì)胞,從而達(dá)到耐砷效果。近幾年,由于微生物的耐砷機(jī)制而演化出利用微生物進(jìn)行砷污染的修復(fù)技術(shù)也受到了越來越多的關(guān)注。除此之外,有些耐砷微生物還有植物促生作用,可以促進(jìn)蜈蚣草[15]、粉葉蕨[16]、大葉井口邊草[17]等一些砷富集植物對砷的吸收,從而達(dá)到微生物—植物協(xié)同作用修復(fù)砷污染的目的。
在土壤砷污染的來源中,礦業(yè)開采是主要原因之一。湖南的康家灣鉛鋅礦是中國的第四大礦區(qū),至今已有100多年的開采歷史,在開采過程中,周圍環(huán)境被各種重金屬污染,如鉛、鋅、砷等,對人們的生活構(gòu)成很大的威脅。目前國內(nèi)外關(guān)于砷污染的環(huán)境中高耐砷性菌株分離的研究較少。本研究以該礦區(qū)作為研究對象,目的是分離篩選耐高濃度砷的細(xì)菌,并對其進(jìn)行鑒定,通過研究其對砷的氧化還原能力、砷耐性基因及其他的與砷耐性相關(guān)的特性,從而確定目標(biāo)菌株,進(jìn)行下一步砷代謝機(jī)制的研究,旨為土壤砷污染的生物修復(fù)奠定理論基礎(chǔ)。
1 材料與方法
1.1 材料
1.1.1 供試土壤 采自湖南康家灣水口山鉛鋅礦(26°33'57.9"N 112°36'23.7"E)周圍礦渣區(qū)10-20 cm的表層土壤。采樣點(diǎn)周圍覆蓋冶煉礦渣,重金屬含量較高,表層植物較少。去除土壤表面礦渣,現(xiàn)場測定pH值,用無菌鏟取約5 kg左右的新鮮表層土壤,置于無菌袋中,做好標(biāo)記,將無菌袋放入冰盒中空運(yùn)回實(shí)驗(yàn)室。取一部分新鮮樣品,立刻進(jìn)行細(xì)菌分離,剩余的樣品存于4℃冰箱中,用于化學(xué)分析,土壤的理化性質(zhì)見表1。

表1 供試土壤的基本性質(zhì)
1.1.2 培養(yǎng)基 LB液體培養(yǎng)基:酵母提取物5 g,氯化鈉10 g,胰蛋白胨10 g,蒸餾水1 000 mL,pH 7.2。CDM培養(yǎng)基[18]:溶液 A(0.081 2 mol/L七水硫酸鎂,0.187 mol/L氯化銨,0.07 mol/L無水硫酸鈉,0.574 mmol/L磷酸氫二鉀,4.57 mmol/L二水合氯化鈣和0.446 mol/L乳酸鈉),121℃滅菌20 min;溶液B(4.8 mmol/L七水硫酸亞鐵)和溶液C(0.95 mol/L碳酸氫鈉),用0.45 μm過濾器過濾除菌。分別配制上述3種溶液,1 L CDM液體培養(yǎng)基中含有100 mL溶液A、2.5 mL溶液B、10 mL溶液C和887.5 mL蒸餾水,調(diào)pH至7.2。上述培養(yǎng)基若制備固體培養(yǎng)基,加入14 g/L的瓊脂粉即可。
1.1.3 主要試劑和儀器 細(xì)菌基因組DNA提取試劑盒和購自天根生化科技有限公司,PCR擴(kuò)增和克隆相關(guān)試劑購自大連寶生物有限公司(TaKaRa),引物由上海英濰捷基公司合成,測序服務(wù)由生工生物工程(上海)股份有限公司提供。PCR儀和凝膠成像系統(tǒng)分別為Bio-Rad PCR儀和上海Tocan360凝膠成像系統(tǒng)。
1.2 方法
1.2.1 供試土壤理化性質(zhì)的測定 將新鮮土樣進(jìn)行烘干、過篩、消化(1∶1的HNO3-H2SO4)處理,用火焰原子吸收分光光度法[19]測定供試土壤中的重金屬含量。用總有機(jī)碳分析儀(TOC-VE,Shimadzu,日本)檢測有機(jī)碳含量;而總氮和磷的含量用傅里葉變換近紅外光譜儀(NIRLabN-200,BUCHI,瑞士)即可測得。
1.2.2 土壤耐砷菌株的分離和純化 取1 g新鮮土樣溶于99 mL無菌水中,錐形瓶中加入10粒左右的玻璃珠,150 r/min,28℃振蕩培養(yǎng)30 min。將土壤懸液連續(xù)稀釋至濃度10-3,10-4和10-5,然后取0.1 mL稀釋液涂布于含有40 mmol/L Na2HAsO4·7H2O的CDM平板上,28℃倒置培養(yǎng)3-7 d后劃線分離。為了進(jìn)一步篩選出對砷具有較高耐受性的菌株,分別用含不同梯度Na2HAsO4·7H2O(65、130和260 mmol/L)的CDM固體培養(yǎng)基對初篩分離的菌株進(jìn)一步分離培養(yǎng),純化。所有實(shí)驗(yàn)重復(fù)3次。
1.2.3 對砷和其他重金屬耐受性檢測 對篩選出來的耐砷能力較強(qiáng)的菌株進(jìn)行高濃度砷和其他重金屬的耐受性檢測。將分離純化的菌株接種于分別含有400、600、800 mmol/L Na2HAsO4·7H2O和 5、10、20 mmol/L NaAsO2的CDM瓊脂平板上,28℃培養(yǎng)3-7 d。對其他重金屬(鎘、汞、銅、鎳和鋇)的耐受性測定分別選用化合物Cd(NO3)2·4H2O、HgCl2、Cu(NO3)2·3H2O、NiCl2和BaCl2的濃度為1-20 mmol/L,共20個梯度,每兩個梯度間隔1 mmol/L,用平板劃線法將菌株接種于含有不同重金屬濃度的CDM固體培養(yǎng)基上,重復(fù)3次,28℃培養(yǎng)3-7 d。
1.2.4 耐砷菌株的鑒定和耐砷基因的檢測 采用16S rRNA基因的PCR擴(kuò)增和測序分析法對篩選出來的耐砷細(xì)菌進(jìn)行菌種鑒定。利用細(xì)菌基因組DNA提取試劑盒提取耐砷菌株的基因組DNA,并以其作為模板,以27F(5'-AGAGTTTGATCCTGGCTCAG-3')和 1 492R(5'-CGGCTACCTTGTTACGACTT-3')[20]為引物進(jìn)行50 μL 體系的PCR擴(kuò)增。擴(kuò)增程序:94℃變性4 min;94℃ 1 min,55℃ 1 min,72℃ 1.5 min,循環(huán)30次;72℃ 10 min。為了獲得幾乎全長的16S rRNA基因,將PCR擴(kuò)增產(chǎn)物用DNA純化試劑盒(TaKaRa)進(jìn)行純化,然后用切膠回收試劑盒(TaKaRa)回收純化DNA片段,并用pMD18-T克隆試劑盒(TaKaRa)將純化片段鏈接到pMD18-T載體中,10 μL的連接體系包含1 μL T4 DNA Ligase Buffer(10×),7.5 μL DNA,0.5 μL pMD-18T,1 μL T4 DNA 連接酶,16℃連接16 h,然后用氯化鈣法轉(zhuǎn)化到大腸桿菌感受態(tài)細(xì)胞(購自天根生化科技有限公司)中,挑取單克隆進(jìn)行菌落PCR檢測,篩選出陽性克隆,然后送到生工生物工程股份有限公司利用引物M13-47(5'-CGCCAGGGTTTTCCCAGTCACGAC-3')/RV-M(5'-AGCGGATAACAATTTCACACAGGA-3')進(jìn)行測序。用DNAMAN軟件對測序結(jié)果進(jìn)行拼接,將得到的序列提交到http://www.eztaxon-e.ezbiocloud. net/[21]進(jìn)行序列分析,然后用MEGA 5.0[22]軟件中的鄰接法(Neighbor-Joining)構(gòu)建系統(tǒng)發(fā)育樹[23]。
對于砷耐受性相關(guān)基因(As3+氧化酶基因aioA、As5+還原酶基因arsC、外排基因arsB/ACR3(1)/ ACR3(2)、甲基化酶基因arsM)的檢測,我們分別選取了9對引物(表2)進(jìn)行PCR擴(kuò)增。25 μL PCR反應(yīng)體系包含 14.75 μL 2×Taq PCR Mix,上下游引物各0.2 mmol/L,25 ng DNA模板,克隆及測序方法同16S rRNA鑒定。
1.2.5 菌株氧化還原能力測定 采用砷鉬藍(lán)法[7]進(jìn)行測定。其原理為五價砷與鉬酸銨作用生成砷鉬酸絡(luò)合物,然后被抗壞血酸還原成鉬藍(lán),在沸水浴中顯色并在865 nm下吸收峰值達(dá)到最大。挑取單菌落接種于LB液體培養(yǎng)基中,28℃,150r/min 培養(yǎng)24 h。將菌液4 750 × g 離心10 min,用PIPES(20 mmol/L,pH 7.0)緩沖液洗滌菌體兩次,懸浮于分別含有1 mmol/L NaHAsO2和1 mmol/L Na2HAsO4·7H2O的PIPES中,并調(diào)整菌液濃度至OD600= 1.0,28℃,150 r/min 培養(yǎng)。分別在培養(yǎng)24、48和72 h時取1 mL菌液于1.5 mL離心管中,4 750 ×g離心10 min去沉淀,將上清用PIPES緩沖液稀釋10倍,用于測定As5+和As3+含量。具體測定方法:處理一:取300 μL稀釋后的上清液加入100 μL KIO3溶液(5 mmol/L KIO3溶于 50 mmol/L HCl),目的是將上清液中的As3+氧化成As5+;處理二:取300 μL稀釋后的上清液加入100 μL 25 mmol/L HCl,對樣品進(jìn)行酸化。兩份樣品均25℃處理10 min,再分別加入600 μL鉬藍(lán)試劑(每升含有6 g鉬酸銨、10.8 g抗壞血酸、0.136 g酒石酸銻鉀、67.3 mL濃硫酸)立刻放入水浴鍋中,78℃處理10 min,然后冰浴5 min,用分光光度計(jì)在865 nm下測定其吸光度值。所有處理均進(jìn)行3次重復(fù)。以砷(Na2HAsO4·7H2O)濃度(0、20、40、60、80和100 mmol/L)為橫坐標(biāo),吸光度OD865為縱坐標(biāo),繪制標(biāo)準(zhǔn)曲線。處理一和處理二分別測定的是樣品中總As和As5+濃度,而As3+的濃度則根據(jù)兩個處理的吸光度差值計(jì)算可得。
1.2.6 菌株產(chǎn)IAA能力測定 根據(jù)康貽軍等[29]的方法對砷氧還能力較強(qiáng)的菌株進(jìn)行產(chǎn)吲哚乙酸(IAA)能力定量測定。耐砷菌株產(chǎn)IAA濃度測定:用固體LB培養(yǎng)基活化耐砷菌株,然后接種于含有0.5 g/L色氨酸的LB液體培養(yǎng)基中,150 r/min 28℃ 培養(yǎng)48 h。5 000×g 離心15 min取2 mL上清液于10 mL離心管中,再加入等體積的Salkowshi比色液(每升10. 8 mol /L H2SO4含4.5 g FeCl3),室溫避光放置30 min。根據(jù)混合液的顏色可初步判斷產(chǎn)IAA量(若菌株產(chǎn)IAA,混合液則顯示紅色);然后用分光光度計(jì)測定OD530值,定量檢測IAA。以空白培養(yǎng)基作對照,重復(fù)3次。以純IAA濃度(0、20、40、60和80 μg/mL)為橫坐標(biāo),OD530為縱坐標(biāo)作標(biāo)準(zhǔn)曲線。根據(jù)IAA標(biāo)準(zhǔn)曲線的OD530值計(jì)算菌液中IAA濃度。
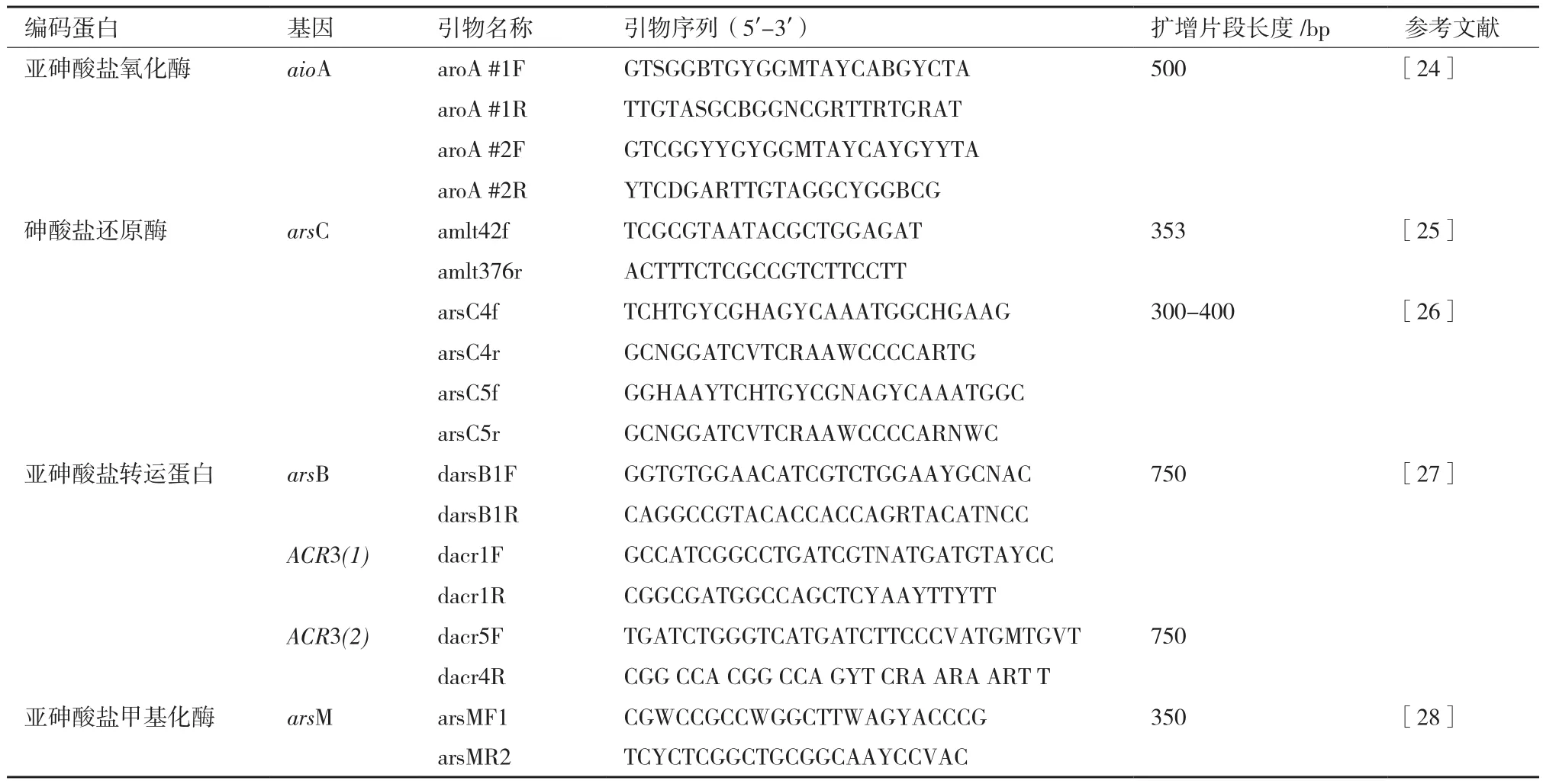
表2 擴(kuò)增與砷代謝相關(guān)基因所選用的引物
2 結(jié)果
2.1 抗砷菌株的分離和篩選
利用含有40 mmol/L Na2HAsO4·7H2O的CDM平板,從采集的土壤樣品中共初篩獲得152株耐砷細(xì)菌。然后用更高濃度的As5+對152株菌進(jìn)行了進(jìn)一步的篩選,在65、130和260 mmol/LNa2HAsO4·7H2O的CDM瓊脂培養(yǎng)基中分別篩選出了42、26和6株砷耐受性菌株(圖1)。
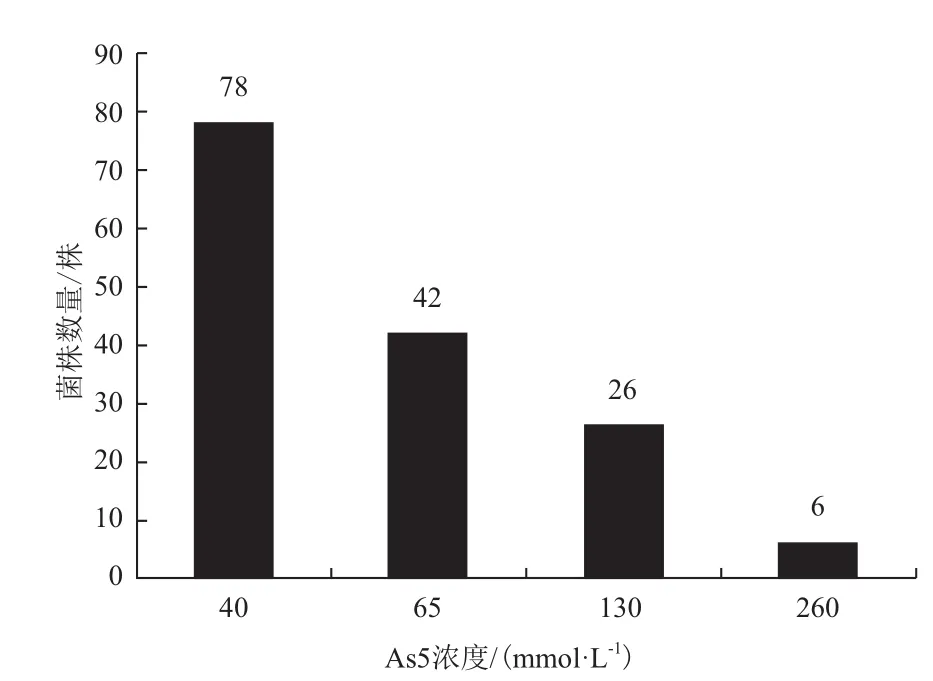
圖1 根據(jù)不同濃度的As5+篩選的菌株數(shù)量
2.2 菌株對砷和其他重金屬的耐受性測定
對篩選出的6株砷高耐受性菌株進(jìn)行砷的最低抑菌濃度和耐其他重金屬性能的測定。從表3可以看出大多數(shù)菌株對As5+的耐受性在600 mmol/L以上,甚至高達(dá)800 mmol/L(Tw133和Tw222);對As3+的耐受性除了Tw31(5 mmol/L)以外,也均在10 mmol/L以上。這些耐砷菌株還對多種重金屬具有耐受性,其中對鎳和銅耐受性較高,對汞和鋇產(chǎn)生中等程度的耐受性,而對低濃度的鎘較為敏感。
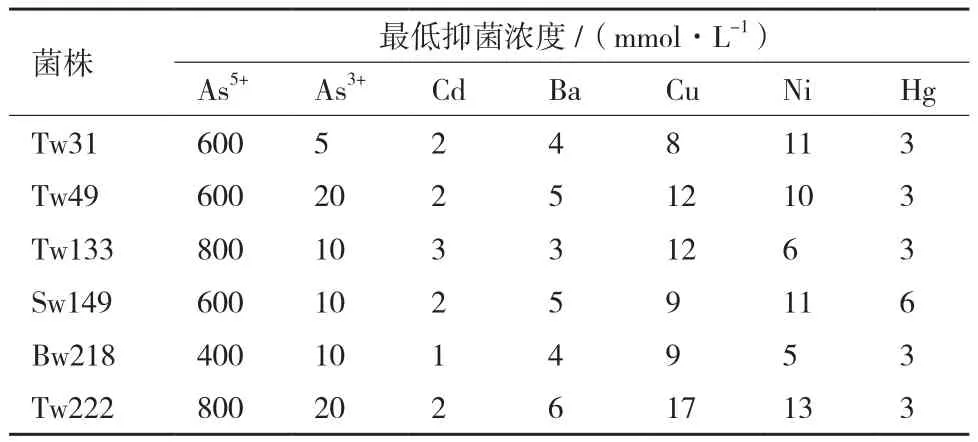
表3 菌株對砷及其他重金屬的耐受能力
2.3 菌株鑒定及相關(guān)耐砷基因檢測
根據(jù)16S rRNA基因序列構(gòu)建的系統(tǒng)發(fā)育樹結(jié)果表明這6株耐砷菌株分屬于5個不同的屬:假單胞菌屬、蒼白桿菌屬、芽孢桿菌屬、威廉氏菌屬和節(jié)細(xì)菌屬(圖2)。
利用9對引物檢測以上6株細(xì)菌的砷耐受性相關(guān)基因,結(jié)果(表4)顯示,4株耐砷菌株含有還原酶基因arsC,其中Tw133中arsC由引物對arsC4r/ arsC4f擴(kuò)增得到,而其他三株菌Tw31、Sw149和Bw218均用amlt42f/amlt376r引物得到。Tw222菌株同時含有兩個外排基因arsB和ACR3(2),菌株Bw218僅擴(kuò)增得到外排基因ACR3(2)。在這6株菌中均沒有擴(kuò)增得到氧化酶基因和甲基化酶基因,并且在菌株Tw49中沒有檢測到任何耐砷基因。
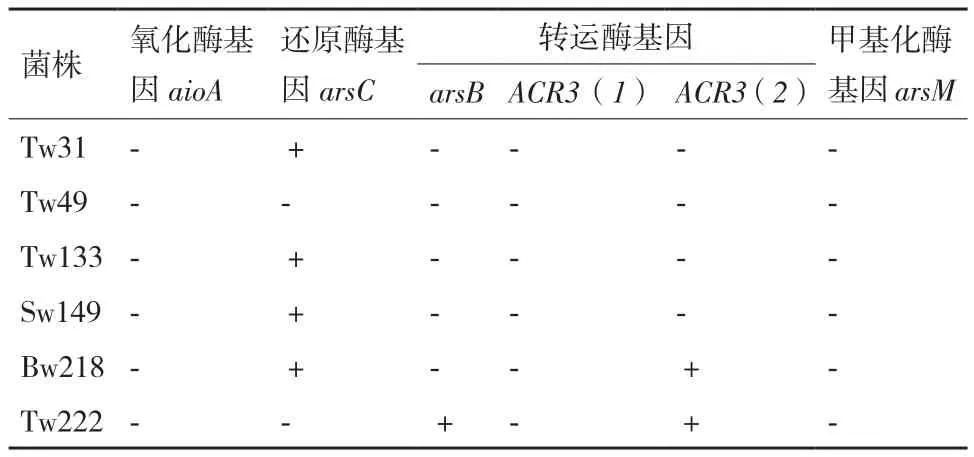
表4 菌株含有的相關(guān)砷耐受性基因
2.4 菌株氧化還原能力測定
用砷鉬藍(lán)法對6株耐高濃度砷的細(xì)菌進(jìn)行砷氧化還原能力測定。結(jié)果(圖3)顯示,這6株菌均表現(xiàn)出了不同程度的氧化還原能力,其中菌株Bacillus sp. Tw133和Pseudomonas sp. Tw222的氧化還原率明顯高于其他菌株。在72 h內(nèi),Bacillus sp. Tw133和Pseudomonas sp. Tw222均可氧化17 %左右的As3+(圖3-A),并可還原大約35 %的As5+(圖3-B)。由于Bacillus sp. Tw133菌株在72 h時的還原率表現(xiàn)出明顯的增長趨勢,因此又對其進(jìn)行了更細(xì)致地測定,結(jié)果表明Bacillus sp. Tw133在144 h還原率達(dá)到最大值48.66 %(圖3-C)。由此可見,Bacillus sp. Tw133和Pseudomonas sp. Tw222菌株在砷氧化還原方面表現(xiàn)出了較明顯的優(yōu)勢。
2.5 菌株產(chǎn)IAA能力測定
IAA是一種植物內(nèi)源激素,與植物的生長發(fā)育相關(guān),一定濃度的IAA可以促進(jìn)植物的生長。通過對優(yōu)勢菌株Bacillus sp. Tw133和Pseudomonas sp. Tw222進(jìn)行產(chǎn)IAA能力的測定。如圖4所示,B管的紅色明顯比C深,根據(jù)顯色結(jié)果初步判斷,Bacillus sp. Tw133的產(chǎn)IAA能力比Pseudomonas sp. Tw222強(qiáng)。根據(jù)IAA標(biāo)準(zhǔn)曲線計(jì)算可得,Bacillus sp.Tw133可以產(chǎn)生高達(dá)42.86 μg/mL的IAA,明顯高于Pseudomonas sp. Tw222(24.36 μg/mL)。而由此可推斷該兩株菌對植物有一定的促生作用。
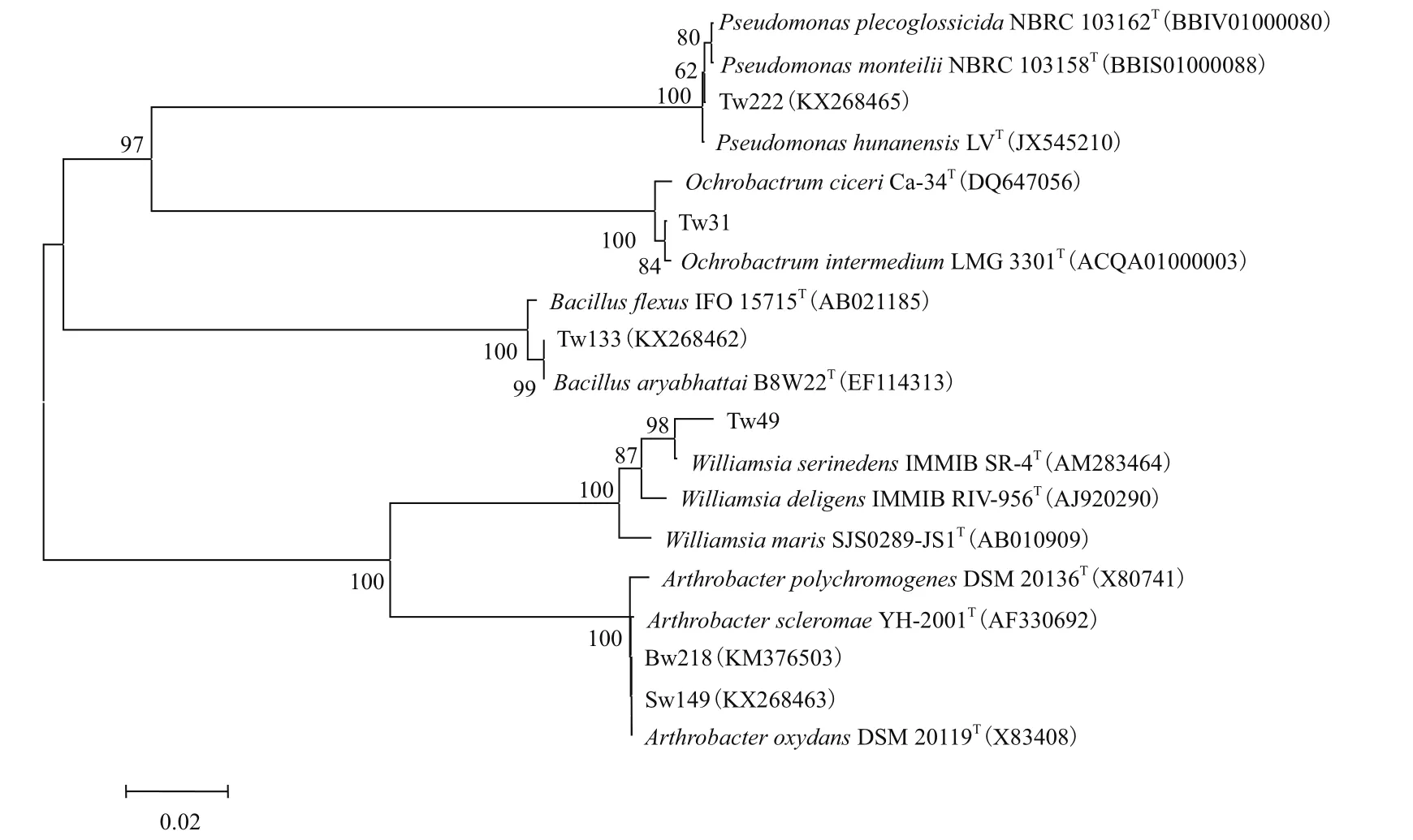
圖2 耐砷菌株與相關(guān)菌的系統(tǒng)發(fā)育樹
3 討論
隨著社會經(jīng)濟(jì)的發(fā)展,土壤砷污染問題越來越嚴(yán)重,其毒性對環(huán)境和人類健康產(chǎn)生極大的威脅。面對這種日益嚴(yán)峻的土壤砷污染問題,迫切需要一種高效、經(jīng)濟(jì)、環(huán)保的方法進(jìn)行修復(fù)。近幾年,越來越多的研究表明,自然界中的微生物在砷的遷移轉(zhuǎn)化過程中發(fā)揮了重要作用[30],因此通過細(xì)菌的代謝活動減少或去除砷污染受到了越來越多的關(guān)注。多種耐砷菌株的分離和鑒定有利于深入研究其代謝機(jī)制,從而促進(jìn)砷污染的生物修復(fù)。
本研究從湖南省康家灣鉛鋅礦土壤中共篩選出152株菌,其中耐高濃度砷的菌株細(xì)菌共6株。16S rRNA序列分析顯示,這6株耐砷菌株屬于5個不同的種屬,分別為假單胞菌屬、蒼白桿菌屬、芽孢桿菌屬、威廉氏菌屬和節(jié)細(xì)菌屬。其中蒼白桿菌屬[31]、芽孢桿菌屬[9]、節(jié)細(xì)菌屬[6]和假單胞菌屬[32]均有報(bào)道。目前關(guān)于威廉氏菌屬的砷耐受性尚未有文獻(xiàn)報(bào)道,因此該研究中獲得的Williamsia sp. Tw49豐富了耐砷細(xì)菌的資源。
從重金屬污染環(huán)境中分離得到的細(xì)菌普遍存在耐砷及其他重金屬的特性。有報(bào)道稱一些細(xì)菌對砷的耐受性分別可達(dá)10 mmol/L(As3+)和100 mmol/L(As5+)[33],甚至在Paul等[8]的研究中發(fā)現(xiàn) Acinetobacter,Microbacterium,Pseudomonas和Rhizobium對砷酸鹽耐受性可高達(dá)600 mmol/L。而本研究篩選得到的6株菌均表現(xiàn)出了更大的砷耐受性,其中對As5+和As3+的最大耐受性分別為800 mmol/L(Bacillus sp. Tw133和 Pseudomonas sp. Tw222) 和20 mmol/L(Williamsia sp. Tw49和 Pseudomonas sp. Tw222),如此高的砷耐受性特征為今后研究該菌的砷代謝機(jī)制提供了理論支持,也為發(fā)現(xiàn)細(xì)菌中新的砷代謝機(jī)理提供了可能性。
為了適應(yīng)砷污染的環(huán)境,細(xì)菌形成了各種砷代謝機(jī)制,目前最常見的耐砷機(jī)理包括砷的氧化、還原、甲基化等[2]。通過耐砷基因檢測,其中5株耐砷細(xì)菌中含有至少一個與砷耐受性相關(guān)的基因。6株耐砷菌株均表現(xiàn)出了不同程度的氧化還原能力,尤其是Bacillus sp. Tw133和Pseudomonas sp. Tw222的氧化率達(dá)17 %左右,但是未檢測到砷氧化酶基因和甲基化酶基因,可能是在這些菌株中存在其他類型的砷氧化酶基因[34]和甲基化酶基因或者選用的引物不合適[35]。根據(jù)As5+還原能力檢測結(jié)果可知,耐砷菌株Bacillus sp. Tw133的還原率在144 h時達(dá)到最大值48.66 %,明顯高于耐砷菌株Enterobacter coacae sp. MC204(38.1 %)[7],而且這與該菌株中檢測到砷還原酶基因(arsC)結(jié)果相符。盡管Williamsia sp. Tw49對砷表現(xiàn)出了較高的耐受性(As5+= 600 mmol/L,As3+= 10 mmol/L),但是用所選引物進(jìn)行PCR擴(kuò)增,沒有檢測到任何砷耐受性基因,推測該菌株可能存在未知的耐砷機(jī)制,有待于進(jìn)一步深入研究。
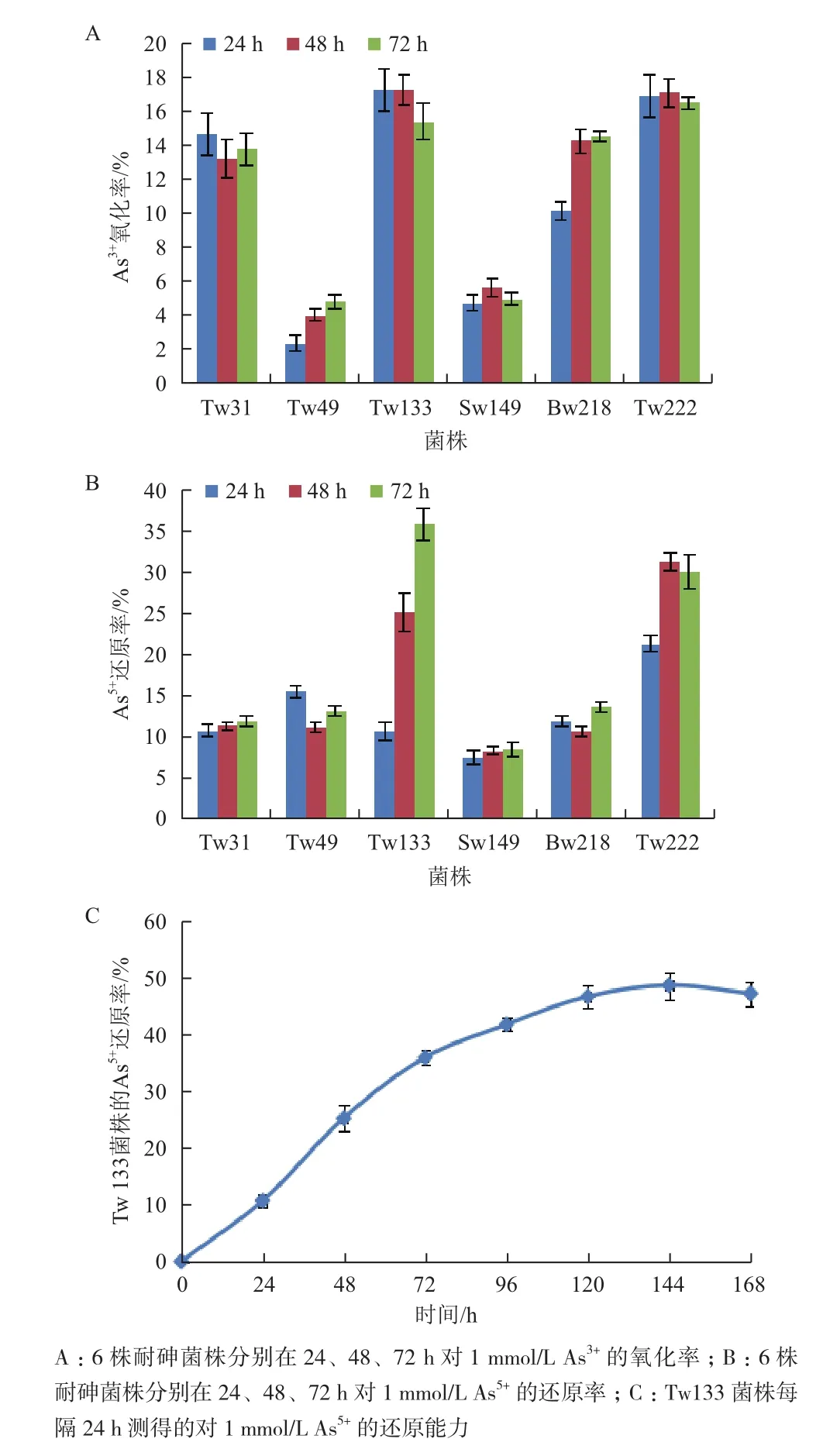
圖3 耐砷菌株分別對As的氧化還原能力

圖4 菌株產(chǎn)IAA能力顯色結(jié)果
植物修復(fù)技術(shù)是指利用植物清除土壤中的污染物質(zhì)或使污染物質(zhì)無毒化的技術(shù)[36],但是較高濃度的重金屬限制了一些耐重金屬植物的正常生長,而從污染地區(qū)分離到的一些細(xì)菌具有植物促生特性(PGP),如產(chǎn)IAA、ACC(1-氨基環(huán)丙烷羧酸)、鐵載體等[37],可以促進(jìn)植物對重金屬的吸收,因此利用微生物—植物協(xié)同作用對重金屬污染土壤進(jìn)行修復(fù)也是一種較為理想的選擇。文一等[38]的研究表明,鏈霉菌 Streptomyces sp. 能夠促進(jìn)砷超積累植物蜈蚣草對砷的進(jìn)一步吸收。本研究中篩選出的耐砷菌株Bacillus sp. Tw133和 Pseudomonas sp. Tw222產(chǎn) IAA的量分別可達(dá)42.86 μg/mL和24.36 μg/mL,均高于植物根際促生菌Pseudomonas sp. WP6[29]產(chǎn)IAA的量20.92 μg/mL。初步表明這兩株優(yōu)勢菌株均具有明顯的促生能力,為植物—微生物聯(lián)合修復(fù)砷污染提供一定的理論依據(jù)。實(shí)驗(yàn)后期將通過測序確定這兩株優(yōu)勢菌的耐砷基因簇,以便研究它們的耐砷機(jī)制,并用基因工程手段對基因簇進(jìn)行改造,提高其代謝砷的能力,為砷污染的生物修復(fù)奠定基礎(chǔ)。
4 結(jié)論
本實(shí)驗(yàn)從湖南康家灣鉛鋅礦區(qū)總共篩選出152株細(xì)菌菌株,其中6株耐砷菌株對As5+和As3+的耐受性分別可高達(dá)800 mmol/L和20 mmol/L,并且對其他重金屬也有不同程度的抗性。這6株砷高耐受性細(xì)菌屬于5個不同的屬,所含有的砷耐受性基因主要為還原酶基因(arsC)和外排基因(arsB、ACR3(2))。Bacillus sp. Tw133和Pseudomonas sp. Tw222對砷的氧化還原能力相對較高,尤其Bacillus sp. Tw133在144 h的As5+還原率最高可達(dá)48.66 %,并且兩株菌產(chǎn)IAA的量分別為42.86 μg/mL和24.36 μg/mL,表現(xiàn)出了較明顯的優(yōu)勢。
[1] 魏復(fù)盛, 陳靜生, 吳燕玉, 等. 中國土壤環(huán)境背景值研究[J].環(huán)境科學(xué), 1991, 12(4):12-19.
[2] 蔡林, 王革嬌. 抗砷性微生物及其抗砷分子機(jī)制研究進(jìn)展[J].微生物學(xué)通報(bào), 2009, 36(8):1253-1259.
[3] Krumova K, Nikolovska M, Groudeva V. Isolation and identification of arsenic-transforming bacteria from arsenic-contaminated sites in Bulgaria[J]. Biotechnol Biotechnol Equip, 2008, 22(2):721-728.
[4] Zargar K, Hoeft S, Oremland R, et al. Identification of a novel arsenite oxidase gene, arxA, in the Haloalkaliphilic, arseniteoxidizing bacterium Alkalilimnicola ehrlichii strain MLHE-1[J]. Journal of Bacteriology, 2010, 192(14):3755-3762.
[5] 張瀠月, 班允赫, 史榮久, 等. 具有砷(Ⅴ)還原能力的硫酸鹽還原菌篩選及生長特性研究[J]. 河南科學(xué), 2015, 33(4):553-558.
[6] Cai L, Liu GH, Rensing C, et al. Genes involved in arsenic transformation and resistance associated with different levels of arsenic-contaminated soils[J]. BMC Microbiol, 2009, 9(4):1-11.
[7] Jareonmit P, Mehta M, Sadowsky MJ, et al. Phylogenetic and phenotypic analyses of arsenic-reducing bacteria isolated from an old tin-mine area in Thailand[J]. World J Microbiol Biotechnol, 2012, 28(5):2287-2292.
[8] Paul D, Poddar S, Sar P. Characterization of arsenite-oxidizing bacteria isolated from arsenic-contaminated groundwater of West Bengal[J]. J Environ Sci Health Part A-Toxic/Hazardous Substances Environ Eng, 2014, 49(13):1481-1492.
[9] Sanyal SK, Mou TJ, Chakrabarty RP, et al. Diversity of arsenite oxidase gene and arsenotrophic bacteria in arsenic-affected Bangladesh soils[J]. AMB Express, 2016, 6(1):1-11.
[10] 韓永和, 王珊珊. 微生物耐砷機(jī)理及其在砷地球化學(xué)循環(huán)中的作用[J]. 微生物學(xué)報(bào), 2016, 56(6):901-910.
[11] Cavalca L, Corsini A, Zaccheo P, et al. Microbial transformations of arsenic:perspectives for biological removal of arsenic from water[J]. Future Microbiol, 2013, 8(6):753-768.
[12] Desoeuvre A, Casiot C, Héry M. Diversity and distribution of arsenic-related genes along a pollution gradient in a river affected by acid mine drainage[J]. Microb Ecol, 2016, 71:672-685.
[13] Yang HC, Rosen BP. New mechanisms of bacterial arsenic resistance[J]. Biomed J, 2016, 39(1):5-13.
[14] Qin J, Rosen BP, Zhang Y, et al. Arsenic detoxification and evolution of trimethylarsine gas by a microbial arsenite S-adenosylmethionine methyltransferase[J]. Proc Natl Acad Sci USA, 2006, 103(7):2075-2080.
[15] 廖曉勇, 陳同斌, 謝華, 等. 磷肥對砷污染土壤的植物修復(fù)效率的影響:田間實(shí)例研究[J]. 環(huán)境科學(xué)學(xué)報(bào), 2004, 24(3):455-462.
[16] 宋書巧, 周永章, 周興, 等. 土壤砷污染特點(diǎn)與植物修復(fù)探討[J]. 熱帶地理, 2004, 24(1):6-9.
[17] 紀(jì)冬麗, 孟凡生, 薛浩, 等. 國內(nèi)外土壤砷污染及其修復(fù)技術(shù)現(xiàn)狀與展望[J]. 環(huán)境工程技術(shù)學(xué)報(bào), 2016, 6(1):90-99.
[18] Weeger W, Lievremont D, Perret M, et al. Oxidation of arsenite to arsenate by a bacterium isolated from an aquatic environment[J]. Biometals, 1999, 12(2):141-149.
[19] 景麗杰, 馬甲. 火焰原子吸收分光光度法測定污染土壤中 5種重金屬[J]. 中國土壤與肥料, 2009, 1:74-77.
[20] Yanagi M, Yamasato K. Phylogenetic analysis of the family Rhizobiaceae and related bacteria by sequencing of 16S rRNA gene using PCR and DNA sequencer[J]. FEMS Microbiol Lett, 1993, 107(1):115-20.
[21] Kim OS, Cho YJ, Lee K, et al. Introducing EzTaxon-e:a prokaryotic 16S rRNA gene sequence database with phylotypes that represent uncultured species[J]. Int J Sys Evol Microbiol, 2012, 62(3):716-721.
[22] Tamura K, Peterson D, Peterson N, et al. MEGA5:Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods[J]. Mol Biol Evol, 2011, 28(10):2731-2739.
[23] Saitou N, Nei M. The neighbor-joining method:A new method for reconstructing phylogenetic trees[J]. Mol Biol Evol, 1987, 4(4):406-425.
[24] Inskeep WP, Macur RE, Hamamura N, et al. Detection, diversityand expression of aerobic bacterial arsenite oxidase genes[J]. Environ Microbiol, 2007, 9(4):934-943.
[25] Sun YM, Polishchuk EA, Radoja U, et al. Identification and quantification of arsC genes in environmental samples by using real-time PCR[J]. J Microbial Methods, 2004, 58(3):335-349.
[26] Escudero LV, Casamayor EO, Chong G, et al. Distribution of microbial arsenic reduction, oxidation and extrusion genes along a wide range of environmental arsenic concentrations[J]. PLoS ONE, 2013, 8(10):e78890.
[27] Achour AR, Bauda P, Billard P. Diversity of arsenite transporter genes from arsenic-resistant soil bacteria[J]. Res Microbiol, 2007, 158(2):128-137.
[28] Jia Y, Huang H, Zhong M, et al. Microbial arsenic methylation in soil and rice rhizosphere[J]. Environ Sci Technol, 2013, 47(7):3141-3148.
[29] 康貽軍, 程杰, 梅麗娟, 等. 植物根際促生菌的篩選及鑒定[J].微生物學(xué)報(bào), 2010, 50(7):853-861.
[30] 楊婧, 朱永官. 微生物砷代謝機(jī)制的研究進(jìn)展[J]. 生態(tài)毒理學(xué)報(bào), 2009, 4(6):761-769.
[31] Branco R, Chung AP, Morais PV. Sequencing and expression of two arsenic resistance operons with different functions in the highly arsenic-resistant strain Ochrobactrum tritici SCII24T[J]. BMC Microbiol, 2008, 8(1):1-12.
[32] 管思琪, 羅蕾, 邢輝, 等. 湖南石門磺廠礦區(qū)尾礦庫抗砷菌株的分離、鑒定及性質(zhì)研究[J]. 江蘇農(nóng)業(yè)科學(xué), 2014, 42(4):300-303.
[33] Jackson CR, Dugas SL. Phylogenetic analysis of bacterial and archaeal arsC gene sequences suggests an ancient, common origin for arsenate reductase[J]. BMC Evol Biol, 2003, 3(1):1-10.
[34] Sultana M, Vogler S, Zargar K, et al. New clusters of arsenite oxidase and unusual bacterial groups in enrichments from arseniccontaminated soil[J]. Arch Microbiol, 2012, 194(7):623-635.
[35] Cordi A, Pagnout C, Devin S, et al. Determination of physiological, taxonomic, and molecular characteristics of a cultivable arsenicresistant bacterial community[J]. Environ Sci Poll Res, 2015, 22(18):13753-13763.
[36] 洪雪花, 張婉靜, 張愛, 等. 環(huán)境砷的存在狀態(tài)、生物轉(zhuǎn)化和修復(fù)研究進(jìn)展[J]. 云南農(nóng)業(yè)大學(xué)學(xué)報(bào), 2011, 26(4):567-571.
[37] Shagol CC, Krishnamoorthy R, Kim K, et al. Arsenic-tolerant plantgrowth-promoting bacteria isolated from arsenic-polluted soils in South Korea[J]. Environ Sci Pollut Res, 2014, 21(15):9356-9365.
[38] 文一, 廖曉勇, 閻秀蘭. 鏈霉菌的抗砷特性及其對蜈蚣草富集砷的作用[J]. 生態(tài)毒理學(xué)報(bào), 2013, 8(2):186-193.
(責(zé)任編輯 狄艷紅)
Isolation,Identification and Characterization of Arsenic-tolerant Bacteria from Lead-zinc Mine Tailing
WU Dan ZHANG Zhi-peng MA Yu-chao
(College of Biological Sciences and Biotechnology,Beijing Forestry University,Beijing 100083)
The aim of this study is to isolate highly arsenic-tolerant bacterial strains from heavy metal contaminated soils from the Kangjiawan lead-zinc mine,Hunan Province. Serial dilution and plating method was used to isolate arsenic-tolerant bacteria,and the strains were identified by 16S rRNA sequence analysis. Then the detection of arsenic tolerance-related genes was carried out. The arsenic molybdenum blue experiment was conducted to evaluate arsenic oxidation-reduction ability of arsenic-tolerant bacteria. Furthermore,the indole acetic acid(IAA)produced by the dominant bacterial strains was quantitatively determined. A total of 152 arsenic-tolerant bacterial strains were isolated from soils samples,and six strains were resistant to As5+and As3+up to 800 mmol/L and 20 mmol/L,respectively. The six arsenictolerant bacterial strains belonged to five different genera,Pseudomonas,Ochrobactrum,Bacillus,Williamsia,and Arthrobacter. In arsenic tolerance-related gene detection experiments,arsenate reductase gene arsC existed in strains Tw31,Tw133,Sw149,and Tw222,and strains Bw218 and Tw222 contained arsenic ion efflux permease gene arsB/ACR3(2). Within 72 h,the As3+oxidation rate and As5+reduction rate of strains Tw133 and Tw222 were higher than those of other strains,which were about 17 % and 35 %,respectively. In particular,strain Tw133 had an As5+reduction rate of 48.66 % at 144 h. Moreover,the synthetic quantities of IAA in the two strains were 42.86 μg/mL and 24.36 μg/mL,respectively. The isolated strains Tw133 and Tw222 showed obvious advantages in terms of arsenic tolerance,arsenic oxidationreduction capacity and IAA production,providing experimental materials for the further study of arsenic-tolerant mechanism in bacteria.
arsenic-tolerant bacteria;arsenic-tolerant gene;arsenic oxidation-reduction;IAA
10.13560/j.cnki.biotech.bull.1985.2016-0983
2016-10-27
國家十二五科技支撐計(jì)劃(2012BAC09B03),國家自然科學(xué)基金項(xiàng)目(J1310005),北京市科技新星項(xiàng)目(2011033),中央高校基本科研基金項(xiàng)目(2016JX03)
吳丹,女,碩士研究生,研究方向:重金屬污染土壤中耐砷微生物;E-mail:wuzhdan@163.com
馬玉超,女,博士,副教授,研究方向:微生物資源開發(fā)與利用;E-mail:mayuchao@bjfu.edu.cn
猜你喜歡
發(fā)明與創(chuàng)新(2022年30期)2022-10-03 08:40:56
小學(xué)科學(xué)(學(xué)生版)(2021年7期)2021-07-28 06:44:32
動漫星空(興趣百科)(2020年12期)2020-12-12 05:31:40
小學(xué)科學(xué)(學(xué)生版)(2020年6期)2020-08-13 07:47:12
當(dāng)代陜西(2019年7期)2019-04-25 00:22:18
領(lǐng)導(dǎo)決策信息(2018年26期)2018-10-12 02:18:26
人大建設(shè)(2018年6期)2018-08-16 07:23:10
文理導(dǎo)航·科普童話(2017年5期)2018-02-10 19:42:14
無人機(jī)(2017年10期)2017-07-06 03:04:36
小星星·閱讀100分(低年級)(2015年10期)2015-10-22 08:30:04
