茶樹對鉛脅迫的響應及其組織鉛化學形態變化研究
2017-05-30 10:48:04葉江華吳承禎賈小麗陳曉婷李遠華王飛權盧莉鄭茂鐘胡永樂張清旭王海斌
熱帶作物學報 2017年9期
葉江華 吳承禎 賈小麗 陳曉婷 李遠華 王飛權 盧莉 鄭茂鐘 胡永樂 張清旭 王海斌
摘 要 為了分析不同茶樹對Pb(鉛)脅迫的響應及其組織中不同化學形態Pb的累積特性,本研究采用盆栽種植法,探討鐵觀音、肉桂2種茶樹在不同濃度Pb脅迫下,茶樹葉片光合生理指標,包括光合作用速率和葉綠素含量,以及茶樹組織Pb含量和不同化學形態Pb含量的變化。結果表明,隨著Pb濃度的增加,茶樹光合生理指標呈現下降趨勢,且與對照相比,鐵觀音下降的幅度高于肉桂。同一Pb脅迫濃度下,茶樹組織中Pb含量表現為根>葉。不同化學形態Pb含量分析結果表明,隨著Pb脅迫濃度的升高,2種茶樹根部可交換態Pb、碳酸鹽結合態Pb和殘留態Pb含量百分比呈現下降趨勢,而有機結合態Pb和鐵錳氧化態Pb含量呈現上升趨勢;2種茶樹葉片碳酸鹽結合態Pb、可交換態Pb、鐵錳氧化態Pb和有機結合態Pb百分含量呈現上升趨勢,而殘留態Pb百分含量呈現下降趨勢。進一步分析發現,在Pb脅迫下,不同的茶樹表現出解毒模式的差異性。
關鍵詞 茶樹;Pb脅迫;化學形態;光合生理指標
中圖分類號 S571.1 文獻標識碼 A
Biological Responses and Pb Speciation in Tissues After
Pb Exposure of Tea Plants
YE Jianghua1,4, WU Chengzhen1, JIA Xiaoli1,3, CHEN Xiaoting2,3, LI Yuanhua4, WANG Feiquan4,
LU Li4, ZHENG Maozhong1, HU Yongle1, ZHANG Qingxu2, WANG Haibin2,3 *
1 Fujian Provincial Key Laboratory of Eco-Industrial Green Technology(Wuyi University), Wuyishan, Fujian 354300, China
2 College of Life Sciences, Longyan University, Longyan, Fujian 364012, China
3 Fujian Provincial Key Laboratory of Agroecological Processing and Safety Monitoring, Fujian Agriculture and Forestry
University, Fuzhou, Fujian 350002, China
4 College of tea and food science, Wuyi University, Wuyishan 354300, China
Abstract In order to analyze the physiological responses of different tea trees to Pb stress and accumulation characteristics of Pb with different chemical forms in different tissues of tea trees, photosynthesis rate, chlorophyll content, Pb content in different tissues and Pb content with different chemical forms of Tieguanyin and Rougui tea plants were measured under Pb stess with different concentrations in pot culture. Results showed that the photosynthesis rate and chlorophyll content of tea plants descended with the increase of Pb stress concentration, but the falling range of Tieguanyin was higher than Rougui as compared with the control. At the same concentration of Pb stress, Pb contents in tea plant roots and leaves performed root> leaf. Analysis of Pb contents with different chemical forms showed that the content percentages of exchangeable Pb, carbonate-bound Pb, residual Pb from two tea tree roots had downward trends, while organic-bound Pb and Pb with Fe-Mn oxidation state had upward trends as stress concentration increased. Content percentages of exchangeable Pb, carbonate-bound Pb, organic-bound Pb and Pb with Fe-Mn oxidation state in the leaf appeared to be rising, while residual Pb declined. Further analysis indicated that detoxification patterns of different tea plant varieties exhibited variance under Pb stress.
Key words Tea plant; Pb stress; chemical form; physiological characteristics
doi 10.3969/j.issn.1000-2561.2017.09.005
近年來,隨著工業化進程的加快,農業生產中大量使用含重金屬元素的農藥及肥料,農業灌溉用水重金屬污染等問題的日益嚴重,導致土壤重金屬污染加劇[1]。茶樹作為喜酸性植物,長期種植后土壤的酸化程度不斷增強,重金屬活躍能力提高,有效態重金屬含量在土壤中呈現上升趨勢,可能茶樹對重金屬的耐受力有所提升,同時,也極易導致茶樹組織增加重金屬積累量,進而導致茶葉產品中重金屬含量超標,嚴重損害了茶葉的經濟效益,降低了茶農收入[2-5]。
茶樹易吸收累積鉛離子,鉛(Pb)主要通過根系或葉片進入植物體,經過大量累積,會導致植物體中活性氧代謝失調,嚴重者還會影響植物生長發育[6]。眾多學者探討了重金屬對植物生長的影響,并認為高濃度重金屬會對植物生長具有一定的毒害作用[7-9]。目前,關于Pb對茶樹生長的影響體現在以下2方面,一方面土壤Pb含量對茶樹生長的影響,研究表明茶園土壤中各形態Pb的含量為硫化物殘渣態>碳酸鹽結合態>吸附態>有機結合態>交換態>水溶態,其中土壤中有效性Pb含量的高低決定了Pb對茶樹的毒害能力[10];另一方面,Pb離子進入到茶樹體內后,在茶樹體內分布的形態、特征、活性、遷移能力等是影響茶樹生長的關鍵[11-12]。
近年來,茶葉經濟價值的提升,含重金屬的農藥、化肥、有機肥等被大量施用,從而保證茶葉的產量,一方面致使茶園土壤中Pb含量增加,另一方面增加茶樹Pb元素積累量[4-5]。而前人研究主要集中在茶葉加工成品中重金屬的溶出率、重金屬在茶樹中的累積特性及重金屬對茶樹生長的影響等[13-15],關于重金屬脅迫后,其離子在茶樹體內的積累及化學形態變化未做較深入研究。鑒于此,本研究以鐵觀音、肉桂2種具有代表性的烏龍茶樹為研究對象,探討Pb脅迫下,茶樹光合生理特征變化及其組織中Pb累積特性和化學形態變化,以期為研究Pb對茶樹的毒害機理和茶樹對Pb的響應機制提供理論依據。
1 材料與方法
1.1 材料
1.1.1 植物材料 Pb脅迫處理30 d后,收集茶樹根(離根尖2 cm)和倒二葉,用于測定茶樹根、葉中Pb含量和不同化學形態Pb含量。茶樹葉片收集后,用自來水和去離子水洗滌3次,茶樹根收集后仔細清除表面附著的土壤,用去離子水洗滌10次,避免根部表面附著金屬離子。
1.1.2 茶樹種植與不同處理 將未種植過茶樹的土壤風干并研磨,過40目篩;土壤的基本理化指標為pH5.21、有機質18.43 g/kg、全氮2.05 g/kg、全磷0.43 g/kg、全鉀4.35 g/kg、速效氮124.52 mg/kg、速效磷13.17 mg/kg、速效鉀126.31 mg/kg、Pb 2.13 mg/kg。將土壤裝入盆中,每盆裝土10 kg,選擇長勢相對一致的1年生鐵觀音、肉桂茶苗,分別移栽到土壤中,每盆種植茶苗5株;茶苗移栽恢復生長1個月后,將茶苗種植土壤適當攪動使土壤松動,將配置好的醋酸鉛溶液緩慢倒入土壤中,盡量使處理液在土壤中分布均勻,每盆添加量2 L,使土壤中的Pb濃度分別達到50、150、250、350、450 mg/kg,未添加Pb溶液處理的土壤為對照(定義為0 mg/kg);處理后,常規種植茶樹30 d,每個處理種植6盆,即6個重復。
1.2 方法
1.2.1 茶苗光合生理指標測定 Pb脅迫處理30 d后,選取不同處理下的茶苗倒二葉(功能葉),采用LI-6400便攜式光合儀測定茶苗功能葉光合作用速率;采用SPAD-502葉綠素儀測定茶苗倒二葉的SPAD值,即葉綠素含量,每個樣品重復測定6次。
1.2.2 茶苗組織Pb含量測定 參照趙先明等[16]研究的方法,并略作修改。將不同濃度Pb處理下,茶苗的根、葉于80 ℃干燥至恒重,并分別粉碎過40目篩,待測。分別稱取上述樣品0.5 g置于150 mL聚四氟乙烯燒杯中,加入25 mL濃硝酸和高氯酸混合液(體積比為5 ∶ 1),蓋上表面皿,浸泡過夜。次日,160 ℃加熱消解,至消化液呈淡黃色或無色,體積約為1~2 mL為止。冷卻后,用少量蒸餾水多次洗滌燒杯,合并洗滌液并轉入50 mL的容量瓶中,定容,混勻過濾,待測,且每個樣品3個重復。同時,以試劑空白為對照。通過原子吸收分光光度法進行數據測定。
1.2.3 茶苗組織中不同化學形態Pb含量測定
不同濃度Pb處理下,采用化學試劑逐步提取法提取茶苗組織中不同化學形態的Pb[17-18],所需提取劑及提取順序依次為:80%乙醇,提取可交換態Pb;去離子水,提取碳酸鹽結合態Pb;1 mol/L的氯化鈉溶液,提取有機結合態Pb;2%醋酸,提取鐵錳氧化態Pb;0.6 mol/L的鹽酸,提取殘留態Pb。具體提取方法為:稱取2 g茶苗組織新鮮樣品置于燒杯中,加入25 mL提取劑,浸沒樣品,置于30 ℃恒溫箱中靜置18 h,回收提取液。繼續添加等體積的同一提取劑浸提2 h,再次回收提取液,重復3次,即24 h內完成4次提取。回收的提取液轉移至燒杯。依照上述方法對各樣品進行不同提取劑的提取,同時每個樣品設置3個重復。
將不同提取劑獲得的提取液蒸發至近干,去離子水定容至10 mL,以相應提取劑為空白對照,原子吸收分光光度法測定樣品中Pb含量。
1.3 數據統計分析
數據使用Excel、DPS軟件進行方差分析、顯著性分析以及相關性分析。
2 結果與分析
2.1 光合作用速率、葉綠素含量分析
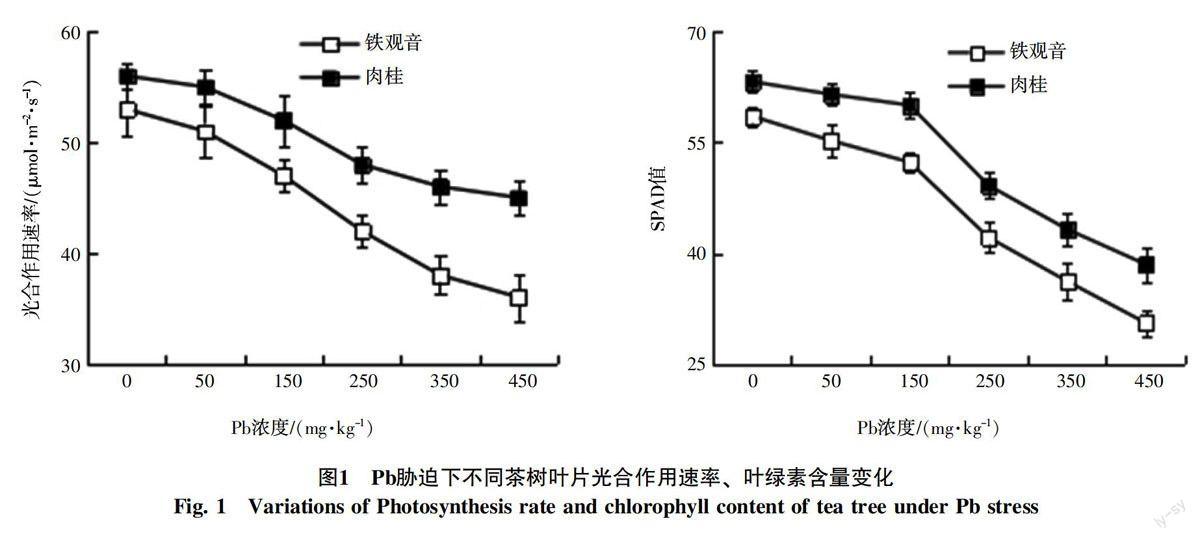
Pb脅迫下,隨著Pb濃度的增加,茶樹葉片光合生理相關指標呈現下降趨勢(圖1)。與對照相比,隨著Pb濃度的升高(50~450 mg/kg),鐵觀音茶樹葉片的光合作用速率、葉綠素含量下降率分別由3.77%上升至32.08%、5.64%上升至47.78%,肉桂茶樹葉片的光合作用速率、葉綠素含量下降率分別由1.79%上升至19.64%、2.72%上升至39.10%;可見,2種茶樹葉片的光合作用速率、葉綠素含量響應Pb脅迫的敏感性存在一定差異。
2.2 Pb含量分析
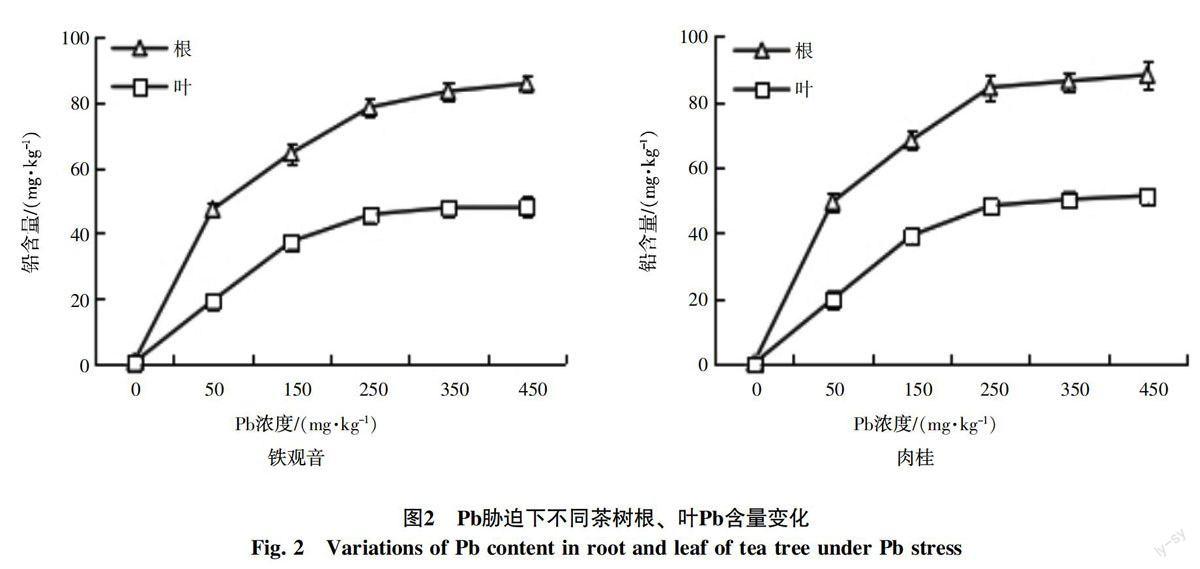
不同Pb濃度脅迫下,隨著土壤Pb濃度的增加,茶樹根、葉的Pb含量均呈現上升趨勢,且根部Pb含量明顯高于葉部(圖2)。進一步分析發現,隨著Pb濃度的升高(0~450 mg/kg),鐵觀音茶樹根部Pb含量由0.53 mg/kg上升至85.87 mg/kg,肉桂則由0.56 mg/kg上升至88.12 mg/kg;葉部Pb含量,鐵觀音由0.27 mg/kg上升至48.14 mg/kg,肉桂由0.29 mg/kg上升至51.38 mg/kg。可見,不同濃度Pb脅迫下,不同茶樹根、葉中Pb含量差異較小。
2.3 不同化學形態Pb含量分析
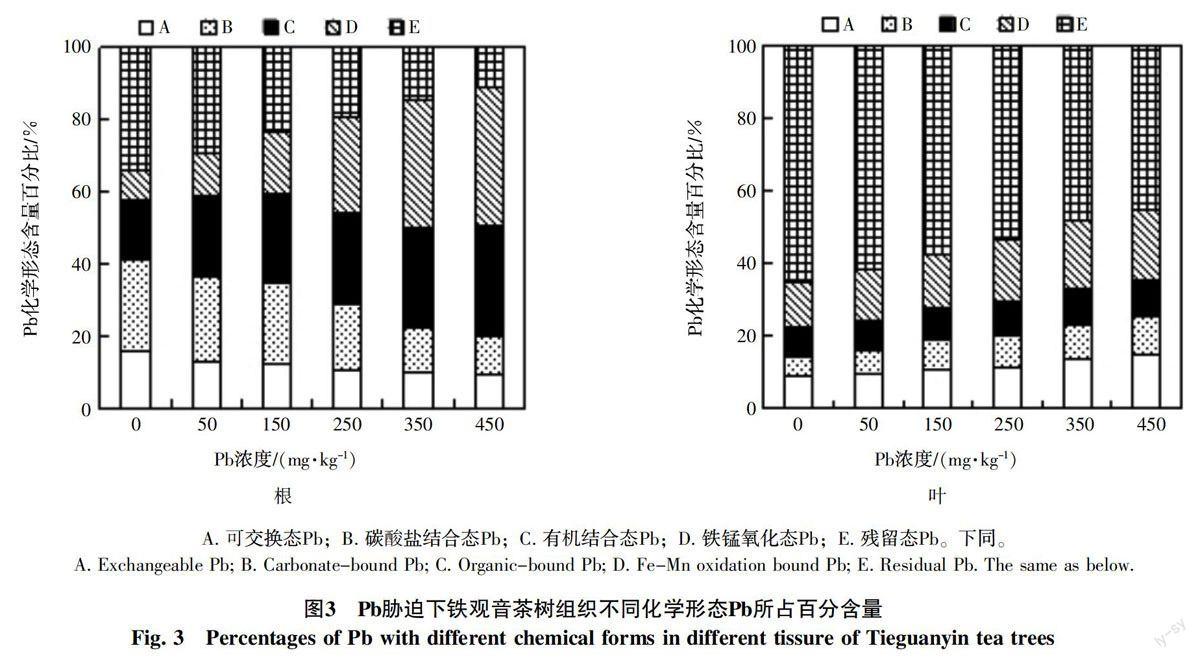
由圖3和圖4可知,隨著Pb濃度的增加(0~450 mg/L),2種茶樹根部可交換態Pb、碳酸鹽結合態Pb和殘留態Pb百分含量呈現下降趨勢,鐵觀音分別為9.82%~18.41%、10.21%~23.24%、12.13%~31.45%,肉桂分別為9.35%~16.17%、10.79%~25.21%、11.06%~34.26%;而有機結合態Pb和鐵錳氧化態Pb百分含量在2種茶樹中均表現為上升趨勢,即鐵觀音分別為19.52%~42.53%和7.38%~25.31%,肉桂分別為16.32%~30.53%和8.04%~38.27%。可見,Pb脅迫下,隨著土壤Pb脅迫濃度的增加,茶樹根部有機結合態Pb和鐵錳氧化態Pb百分含量不斷上升,而可交換態Pb、碳酸鹽結合態Pb和殘留態Pb則相反;鐵觀音與肉桂2種茶樹在可交換態Pb、碳酸鹽結合態Pb和殘留態Pb百分含量差異較小,而有機結合態Pb百分含量上,鐵觀音明顯高于肉桂;鐵錳氧化態Pb百分含量則肉桂明顯高于鐵觀音。
由圖3和圖4可知,隨著Pb濃度的增加,鐵觀音和肉桂葉部可交換態Pb、碳酸鹽結合態Pb、有機結合態Pb和鐵錳氧化態Pb百分含量呈現上升趨勢,鐵觀音分別為9.31%~21.23%、5.13%~9.58%、6.87%~15.27%和10.31%~22.65%,肉桂分別為8.83%~15.14%、5.48%~10.23%、7.96%~10.12%和12.46%~19.27%;而殘留態Pb百分含量呈現下降趨勢,鐵觀音為31.27%~68.38%,肉桂則為45.24%~65.27%。分析表明,Pb脅迫下,鐵觀音葉片的可交換態Pb、碳酸鹽結合態Pb、有機結合態Pb百分含量明顯高于肉桂,鐵錳氧化態Pb百分含量則低于肉桂。
2.4 生理指標及其組織Pb含量的相關性分析
Pb脅迫下,茶樹根部指標相關性分析表明(表1),土壤Pb濃度與鐵觀音茶樹根部Pb、根部可交換態Pb、有機結合態Pb、鐵錳氧化態Pb含量呈顯著或極顯著正相關;而土壤Pb濃度與肉桂茶樹根部Pb含量、根部有機結合態Pb含量、鐵錳氧化態Pb含量呈顯著或極顯著正相關;此外,2種茶樹的根部Pb含量均與茶樹根部可交換態Pb、有機結合態Pb、碳酸鹽結合態Pb、鐵錳氧化態Pb、殘留態Pb含量呈顯著或極顯著正相關。可見,隨著土壤中Pb濃度的增加,茶樹根部Pb含量呈顯著上升趨勢,而根部組織中Pb含量變化會顯著性影響根部組織中不同化學形態Pb含量的變化。
茶樹葉片指標的相關性分析結果表明(表2),Pb脅迫下,土壤Pb濃度與兩種茶樹葉片Pb、葉片可交換態Pb、碳酸鹽結合態Pb、有機結合態Pb、鐵錳氧化態Pb、殘留態Pb含量呈顯著或極顯著正相關,而與葉片光合作用速率、葉綠素含量呈極顯著負相關;兩種茶樹葉片Pb含量與葉片可交換態Pb、碳酸鹽結合態Pb、有機結合態Pb、鐵錳氧化態Pb、殘留態Pb含量呈顯著或極顯著正相關,而與葉片光合作用速率、葉綠素含量呈顯著或極顯著負相關。可見,隨著Pb濃度的增加,茶樹葉片Pb含量及其不同化學形態Pb含量均顯著上升,而葉片生理指標-光合作用速率、葉綠素含量則顯著下降。
3 討論
Pb是植物的非必需元素,過量的Pb進入植株體內,會引起植物的一系列生理響應,如植物吸收過量的Pb離子,易導致植物光合系統受損,最終影響植物產量和品質[19-20]。本研究中,鐵觀音和肉桂茶樹葉片光合作用速率、葉綠素含量對Pb脅迫水平的敏感性有所不同,隨著Pb脅迫水平的升高,2種茶樹葉片的光合作用速率、葉綠素含量均不斷下降(圖1,表2),與前人有關研究[21-23]結果一致;與對照相比,同一濃度Pb作用下,鐵觀音葉片光合生理相關指標含量下降率明顯高于肉桂(圖1)。可見,Pb脅迫可能導致茶樹生理特性變化,而鐵觀音表現出對Pb脅迫的敏感性優于肉桂,肉桂具有更強的重金屬耐受性和適應性。
Pb離子通過根系吸收,在植物體內轉運,并在不同的部位積累[24]。據報道,茶樹體內Pb含量可以明顯高于周圍環境,且體內器官組織中Pb的分布率差異明顯[25]。本研究表明,隨著Pb濃度的升高,茶樹不同組織中Pb含量均呈現上升趨勢,且Pb含量趨勢為根>葉,鐵觀音和肉桂同一組織的Pb累積量無明顯差異(圖2)。可見,不同的茶樹在Pb吸收和累積等方面存在著一定的相似性。
植物對金屬離子的耐受性體現為:一方面由于自身對金屬離子濃度的拮抗作用,減少攝入;另一方面是不同植物對吸收后的金屬離子的轉化及解毒機制存在差異[26-27]。比如,金屬離子進入植物體后,被轉化為不同化學形態,即植物響應金屬離子脅迫的策略表現為不同化學形態金屬離子的含量及其所占比例的不同[28]。據報道,茶樹具有吸收并累積鉛的生理特征,那么茶樹可能存在鉛的特殊解毒機制[29]。本研究發現,Pb脅迫下,隨著Pb脅迫濃度的增加,茶樹根部有機結合態Pb和鐵錳氧化態Pb含量百分比不斷上升,而可交換態Pb、碳酸鹽結合態Pb和殘留態Pb則相反;2種茶樹葉部可交換態Pb、有機結合態Pb、碳酸鹽結合態Pb和鐵錳氧化態Pb百分含量呈現上升的趨勢,殘留態Pb含量則相反(圖3、圖4)。前人研究表明,可交換態的金屬是毒害植物、影響其物質代謝的關鍵形態,碳酸鹽態金屬離子結合較弱,容易釋放,容易遷移,以上2種形態金屬的比率可作為金屬離子植物造成危害的能力的評價因子[18]。而有機結合態主要是與蛋白質結合,極易影響植物代謝過程中的酶活性及其功能的表達,從而影響植物的生長[30]。鐵錳氧化態和殘留態可將金屬離子固定形成難溶的化合物,也是植物抵御毒害特有的機制[31-32]。本研究中,Pb脅迫下,與肉桂相比,鐵觀音葉片光合生理指標下降幅度更大,對Pb脅迫更敏感,可能是由于二者解毒模式的差異,即葉片中不同化學形態Pb含量存在差異,鐵觀音葉片的可交換態Pb、碳酸鹽結合態Pb、有機結合態Pb百分含量明顯高于肉桂,鐵錳氧化態Pb百分含量則低于肉桂。
綜上所述,本研究以鐵觀音、肉桂為材料,探討不同茶樹品種對Pb脅迫的響應及其組織中不同化學形態Pb的累積特性,結果表明,Pb脅迫可降低2種茶樹的光合作用速率及葉片葉綠素含量,其中鐵觀音的下降幅度高于肉桂;其次,不同茶樹組織中Pb含量表現為根>葉,且不同的茶樹品種對Pb脅迫表現出的解毒模式存在差異,鐵觀音茶樹根、葉部主要以提高有機結合態Pb降低毒害,而肉桂以提高鐵錳氧化態Pb降低毒害。關于不同化學形態Pb在植物組織中形成后,如何影響細胞變化,最終影響植物生長發育,還有待進一步研究。
參考文獻
[1] 段德超, 于明革, 徐 辰, 等. 外源胡敏酸對土壤有機質組分變化的影響及其與茶樹鉛有效性的關系[J]. 土壤學報, 2013, 50(5): 940-947.
[2] Zhang Z C, He X L, Li T X. Status and evalution of the soil nutrients in tea plantation[J]. Procedia Environmental Sciences, 2012, 12: 45-51.
[3] Li W, Zheng Z C, Li T X, et al. Effect of tea plantation age on the distribution of soil organic carbon fraction within water-stable aggregates in the hilly region of western Sichuang, China[J]. Catena, 2015, 133: 198-205.
[4] Alcolea A, Vázquez M, Caparrós A, et al. Heavy metal removal of intermittent acid mine drainage with an open limestone channel[J]. Minerals Engineering, 2012, 26: 86-98.
[5] Davies H, Weber P, Lindsay P, et al. Characterisation of acid mine drainage in a high rainfall mountain environment, New Zealand[J]. Science of the Total Environment, 2011, 409: 2 971-2 980.
[6] 申 璐, 肖 斌, 周 旋, 等. 外源亞精胺對鉛脅迫下茶樹生長的影響[J]. 茶葉科學, 2014, 34(1): 87-94.
[7] Navarro M, Wood R J. Plasma changes in micronutrients following a multivitamin and mineral supplement in healthy adults[J]. Journal of the American College of Nutrition, 2003, 22: 124-129.
[8] Sarma L S, Kumar J R, Reddy K J, et al. Development of an extractive spectro-photometric method for the determination of copper(II) in leaf vegetable and pharmaceutical samples using pyridoxal-4-phenyl-3-thiosemicarbazone (PPT)[J]. Journal of Agricultural and Food Chemistry, 2005, 53: 5 492-5 498.
[9] Wan Ngah W S, Hanafiah M A K M. Removal of heavy metal ion from wastewater by chemically modified plant wastes as adsorbrnts: a review[J]. Bioresource Technology, 2008, 99: 3 935-3 948.
[10] 周國華, 孫彬彬, 賀 靈, 等. 安溪土壤-茶葉鉛含量關系與土壤鉛臨界值研究[J]. 物探與化探, 2016, 40(1): 148-153.
[11] Wang X, Liu Y G, Zeng G M, et al. Sub cellular distribution and chemical forms of cadmium in Bechmeria nivea (L.) Gaud[J]. Environmental and Experimental Botany, 2008, 62: 389-395.
[12] Wu F B, Dong J, Qian Q Q, et al. Sub cellular distribution and chemical forms of Cd and Cd-Zn interaction in different barley genotypes[J]. Chemosphere, 2005, 60: 1 437-1 446.
[13] Wang L, Jiang L, Zhao Z Y, et al. Lithium content of some teas and their infusions consumed in China[J]. Food Science and Biotechnolog, 2014, 23(1): 323-325.
[14] Mehra A, Barker C L. Leaching and bioavailability of aluminium, copper and manganese from tea (Camellia sinensis)[J]. Food Chemistry, 2007, 100: 1 456-1 463.
[15] Seenivasan S, Manikandan N, Muraleedharan N N, et al. Heavy metal content of black teas from South India[J]. Food Control, 2008, 19: 746-749.
[16] 趙先明, 汪艷霞, 杜 曉, 等. 水培茶樹對鉛的吸收與累積特性研究[J]. 茶葉科學, 2011, 31(3): 237-246.
[17] Wu F B, Dong J, Qian Q Q, et al. Subcellular distribution and chemical form of Cd and Cd-Zn interaction in different barley genotypes[J]. Chemosphere, 2005, 60(10): 1 437-1 446.
[18] 趙曉東, 謝英荷, 李廷亮, 等. 植物對污灌區土壤鋅形態的影響[J]. 應用與環境生物學學報, 2015, 21(3): 477-482.
[19] Qiao X Q, Shi G X, Jia R, et al. Physiological and biochemical responses induced by lead stress in Spirodela polyrhiza[J]. Plant Growth Regulation, 2012, 67(3): 217-225.
[20] Zeng L S, Liao M, Chen C L, et al. Effects of lead contamination on soil enzymatic activities, microbial biomass, and rice physiological indices in soil-lead-rice (Oryza sativa L.) system[J]. Ecotoxicology and Environmental Safety, 2007, 67(1): 67-74.
[21] 王瓊瑤. 鎘、 銅、 鉛脅迫下蒲兒根生理特性和富集特征研究[D]. 雅安: 四川農業大學, 2011.
[22] 衣艷君, 李芳柏, 劉家堯. 尖葉走燈蘚 (Plagiomnium cuspidatum)葉綠素熒光對復合重金屬脅迫的響應[J]. 生態學報, 2008, 28(11): 5 437-5 444.
[23] 孫天國, 沙 偉, 劉 巖. 復合重金屬脅迫對兩種蘚類植物生理特性的影響[J]. 生態學報, 2010, 30(9): 2 332-2 339.
[24] Mohammad N, Samar M, Alireza I. Levels of Cu, Zn, Pb, and Cd in the leaves of the tea plant (Camellia sinensis) and in the soil of Gilan and Mazandaran farms of Iran[J]. Food Measure, 2014, 8: 277-282.
[25] Han W Y, Zhao F J, Shi Y Z, et al. Scale and causes of lead contamination in Chinese tea[J]. Environmental Pollution, 2006, 139: 125-132.
[26] Suthee J, Sheila M P, Pimluck K, et al. Copper, lead and zinc removal from metal-contaminated wastewater by adsorption onto agricultural wastes[J]. Environmental Technology, 2015, 36(24): 3 071-3 083.
[27] Mahendran V, Shanmugam S. One-pot synthesis of hydrazono-sulfonamide adducts using Cu(BTU) MOF catalyst and their remarkable AIEE properties: unprecedented copper(II)-catalyzed generation of ketenimine[J]. RSC Advances, 2015, 5: 20 003-20 010.
[28] 江行玉, 趙可夫. 植物重金屬傷害及其抗性機理[J]. 應用與環境生物學學報, 2001, 7(1): 92-99.
[29] 郭 巍. 銅、鉛在土壤-植物系統中環境化學行為的研究[J]. 江蘇環境科技, 2006, 19(2): 10-12.
[30] 彭 勃, 黃小平, 張大文. 泰來藻、 海昌蒲體內銅的化學形態與累積規律[J]. 生態學雜志, 2010, 29(10): 1 993-1 997.
[31] 謝立祥, 常明慶, 王 平, 等. 微波萃取土壤中重金屬Cr的形態分析研究[J]. 環境科學與技術, 2010, 33(12): 90-93.
[32] 陳牧霞, 地里拜爾·蘇力坦, 楊 蕭, 等. 新疆污灌區重金屬含量及形態研究[J]. 干旱區資源與環境, 2007, 21(1): 150-154.
