蟲生廣布擬盤多毛孢侵染對松突圓蚧海藻糖酶活性、海藻糖、葡萄糖和蛋白質含量的影響
2017-05-30 02:17:11王莎莎何錦華羅世念彭鋒羅林黃寶靈呂成群
南方農業學報 2017年5期
王莎莎 何錦華 羅世念 彭鋒 羅林 黃寶靈 呂成群

摘要:【目的】探究蟲生廣布擬盤多毛孢對松突圓蚧營養物質代謝的影響,為深入研究廣布擬盤多毛孢致死松突圓蚧機理及開發利用廣布擬盤多毛孢防治松突圓蚧提供科學依據。【方法】以蟲生廣布擬盤多毛孢GXSTYJ03菌株制得濃度為1×107個孢子/mL的孢子懸浮液感染松突圓蚧,以噴灑無菌吐溫-80為對照,逐日測定松突圓蚧體內海藻糖、葡萄糖、蛋白質含量及海藻糖酶活性,連續測定8 d。【結果】處理組松突圓蚧海藻糖含量在各時間點均低于對照組,且隨處理時間延長呈明顯的下降趨勢,在染菌3 d后顯著低于對照組(P<0.05,下同);海藻糖酶活性在各時間點均高于對照組,在染菌后第3 d顯著高于對照組,5 d后也一直顯著高于對照組;葡萄糖含量在染菌后1~3 d迅速升高,第3 d時升至最大值(3.1479 mmol/L),隨后逐漸降低,在第4~6 d時與對照組無顯著差異(P>0.05),6 d后顯著低于對照組;蛋白質含量在各時間點均低于對照組,且在染菌2 d后一直顯著或極顯著(P<0.01)低于對照組。【結論】蟲生廣布擬盤多毛孢GXSTYJ03菌株侵染松突圓蚧過程中引起松突圓蚧體內海藻糖、葡萄糖和蛋白質含量一定程度降低,是致死松突圓蚧的原因之一。
關鍵詞: 蟲生廣布擬盤多毛孢;松突圓蚧;海藻糖;葡萄糖;海藻糖酶;蛋白質
中圖分類號: S763.306.4 文獻標志碼:A 文章編號:2095-1191(2017)05-0820-05
Influence of Entomogenous Pestalotiopsis disseminata on trehalase activity, trehalose content, glucose content and
protein content in Hemiberlesia pitysophila Takagi
WANG Sha-sha1, HE Jin-hua2, LUO Shi-nian2, PENG Feng 2, LUO Lin 3,
HUANG Bao-ling 1, LYU Cheng-qun1 *
(1 College of Forestry, Guangxi University, Nanning 530004, China; 2 Yulin Forest Pest Control Quarantine Station, Yulin, Guangxi 537000, China; 3 Yuzhou Forest Pest Control Quarantine Station, Yulin, Guangxi 537000, China)
Abstract:【Objective】Influence of Entomogenous Pestalotiopsis disseminata on nutrients metabolism of Hemiberlesia pitysophila Takagi was studied, in order to provide reference for mechanism of Entomogenous P. disseminata causing H. pitysophila Takagi dead and using Entomogenous P. disseminata to control H. pitysophila Takagi. 【Method】Entomogenous P. disseminata GXSTYJ03 strain was utilized to make the spore suspension of 1×107 spores/mL and infected H. pitysophila Takagi. Spraying sterile Tween was taken as control(CK) Contents of trehalose, glucose, protein and trehalase activity in H. pitysophila Takagi were detected for eight consecutive days. 【Result】At all measurement times, trehalose content of treatment group were lower than that of CK group, as treatment time extended, trehalose content of treatment group declined and was significantly lower than that of CK group 3 d after infection(P<0.05, the same below). At all measurement times, trehalase activity of treatment group were higher than that of CK group, and was significantly higher on day 3 of infection and 5 d after infection. Glucose content increased rapidly 1-3 d after infection and reached the peak(3.1479 mmol/L) on day 3, then declined gradually, and the difference with CK group was insignificant on 4-6 d(P>0.05), and significantly lower than that of CK group after 6 d. Protein content of treatment group was lower than that of CK group at all measurement times, and was at significant or extremely significant level(P<0.01) 2 d after infection. 【Conclusion】When Entomogenous P. disseminata GXSTYJ03 strain infecting H. pitysophila Takagi, trehalose, glucose and protein content decrease sharply, which is one of the lethal reasons.
Key words: Entomogenous Pestalotiopsis disseminata; Hemiberlesia pitysophila Takagi; trehalose; glucose; trehalase; protein
0 引言
【研究意義】松突圓蚧(Hemiberlesia pitysophila Takagi)又名松櫛盾蚧,屬同翅目(Homoptera)蚧總科(Cocoidea)盾蚧科(Diaspidae)突圓蚧屬(Hemiberlesia Cockerell)昆蟲,是嚴重危害松屬植物的害蟲(Takagi,1969;Takagi and Tang,1982)。近年來,利用蟲生廣布擬盤多毛孢(Entomogenous pestalotiopsis disseminata)制成生防菌劑防治松突圓蚧取得了理想的效果(方麗英等,2007;黃寶靈等,2009;Lü et al.,2011),但部分地區松突圓蚧危害范圍仍以一定速度繼續擴增。因此,深入研究蟲生廣布擬盤多毛孢對松突圓蚧的致病機制,能為開發具有強效、持久、安全防治松突圓蚧的廣布擬盤多毛孢菌株提供科學依據,促使該項生物防治技術的推廣應用。【前人研究進展】方麗英等(2007)從自然死亡的松突圓蚧死尸上分離得到蟲生廣布擬盤多毛孢并將其制成生防菌防治松突圓蚧,發現以生防菌防治松突圓蚧的效果顯著,其致死率遠高于松突圓蚧自然死亡率。但在生防菌制備過程中無法避免蟲生真菌退化現象,菌株的退化會嚴重影響防治效果,因此郭麗娟等(2014)對蟲生廣布擬盤多毛孢GXSTYJ03菌株的退化原因進行研究,明確了影響該菌株退化的主要因素及菌株復壯的最佳方法,為大批量生產菌粉劑提供了技術保障。黃蓉等(2014)、呂成群等(2014)探討蟲生廣布擬盤多毛孢GXSTYJ03侵染對松突圓蚧胞外酶作用及對松突圓蚧酚氧化酶原系統的影響,掌握了該菌株的生物學特性,分析了影響蟲生廣布擬盤多毛孢對松突圓蚧致病的毒力因子,明確菌株孢子懸浮液1×107個孢子/mL是酶制劑生產和病蟲害防治的最佳使用量,以及松突圓蚧對蟲生廣布擬盤多毛孢侵染的免疫反應。然而昆蟲的免疫反應是一個復雜體系,會調度多種抵抗因子來防御病原真菌入侵,這個過程會消耗大量營養物質,主要表現為海藻糖、葡萄糖和蛋白質含量發生變化。薛皎亮等(2006)對油松毛蟲感染白僵菌后體內蛋白質變化情況進行研究,發現感染白僵菌1~8 d后蟲體內的蛋白質含量持續下降。熊琦(2013)研究發現,桃小幼蟲在受到球孢白僵菌TST05菌株侵染后,其體內的海藻糖和蛋白質含量明顯下降,葡萄糖含量呈先升高后下降趨勢。昆蟲體內的營養物質主要以海藻糖形式存儲,海藻糖存在于昆蟲幾乎所有組織和器官中,利用唯一能將海藻糖分解為葡萄糖的海藻糖酶來供給能量(于彩虹等,2008)。病原真菌菌絲一旦進入昆蟲血腔,便利用昆蟲體內的物質作為營養以保證菌絲的生長和大量繁殖,逐漸感染昆蟲內部組織和器官,最終使昆蟲感染死亡。感染導致寄主昆蟲死亡的因素是多方面的,其中,病菌在蟲體內大量繁殖,耗盡蟲體的營養物質是導致昆蟲死亡的一個重要因素(樊金華等,2013)。【本研究切入點】關于蟲生廣布擬盤多毛孢GXSTYJ03菌株的生產及防治松突圓蚧等方面的研究已較完善,但有關蟲生廣布擬盤多毛孢的致病機制僅見其侵染可抑制松突圓蚧的酚氧化酶原系統功能,從而克服松突圓蚧免疫反應的報道。目前,關于蟲生廣布擬盤多毛孢對松突圓蚧體內營養物質代謝影響的研究尚無文獻報道。【擬解決的關鍵問題】逐日測定感染蟲生廣布擬盤多毛孢GXSTYJ03菌株的松突圓蚧體內海藻糖、葡萄糖、蛋白質含量及海藻糖酶活性,了解蟲生廣布擬盤多毛孢對松突圓蚧營養物質代謝的影響,為廣布擬盤多毛孢致死松突圓蚧機理研究及開發利用廣布擬盤多毛孢防治松突圓蚧提供科學依據。
1 材料與方法
1. 1 試驗材料
供試蟲生廣布擬盤多毛孢GXSTYJ03菌株由廣西大學林學院黃寶靈研究員提供;供試松突圓蚧采自玉林市玉州區林場馬尾松林。
1. 2 試驗方法
1. 2. 1 松突圓蚧染菌 參照呂成群等(2014)的方法染菌。先制備GXSTYJ03菌株的1×107個孢子/mL孢子懸浮液,然后用裝有孢子懸浮液的手動噴霧器將菌液噴于帶有松突圓蚧的松針針葉,保證針葉基部葉鞘部分被菌液噴濕。以噴灑無菌吐溫-80且帶有松突圓蚧的松針為對照組。在20 ℃、濕度95%、14 h/10 h光暗交替條件下室內培養。
1. 2. 2 松突圓蚧海藻糖、葡萄糖、蛋白質含量及海藻糖酶活性測定 處理后1~8 d逐日在對照組和處理組中隨機取樣,每組取100只松突圓蚧作為一個樣品,每組3個樣品作為重復。用預冷的1.5 mL 0.2 mol/L的磷酸緩沖液(pH 7.0)冰浴碾磨蟲體,并將勻漿液最終定容至2.0 mL,4800 r/min離心10~15 min,取上清液待測。
1. 2. 2. 1 海藻糖含量測定 采用蒽酮法(馮慧,1989)測定海藻糖含量。
標準曲線制作:先配制海藻糖標準溶液100 mL,濃度為40 mmol/L[稱取海藻糖(Sigma)1.5132 g,以蒸餾水定容],-70 ℃儲存。用蒸餾水稀釋得7個濃度梯度,分別為0.05、0.10、0.20、0.40、0.60、0.80和1.60 mmol/L。取上述溶液100 μL移入10 mL離心管中,加入100 μL 1%硫酸溶液,在90 ℃條件下加熱10 min,冰浴冷卻,再加入100 μL 30%氫氧化鉀溶液,并再次加熱10 min,冰浴冷卻后加入2.5 mL顯色劑[250 mL 80%的硫酸溶液+0.5 g蒽酮(Sigma)],加熱10 min后冰浴冷卻,立即于630 nm處讀取OD。
海藻糖含量測定:以待測提取液100 μL代替標準溶液,按上述方法進行測定。
1. 2. 2. 2 海藻糖酶活性測定 采用3,5-二硝基水楊酸法(雷芳等,2006)測定海藻糖酶活性,并略有改進。
葡萄糖標準曲線制作:先配制葡萄糖標準溶液200 mL,濃度為20 mmol/L(稱取葡萄糖0.79268 g,以蒸餾水定容),-70 ℃儲存。用蒸餾水稀釋,得濃度為0.2、0.4、0.6、0.8、1.0、1.2和1.4 mmol/L的葡萄糖溶液。分別取上述溶液300 μL移入10 mL離心管中,加入300 μL顯色劑[0.5 g 3,5-二硝基水楊酸(Sigma)+0.1 g苯酚+
0.5 g氫氧化鈉+0.025 g無水亞硫酸鈉,蒸餾水定容至50 mL]于90 ℃條件下加熱5 min,冰浴冷卻,再加入100 μL 40%四水合酒石酸堿鈉水溶液,立即于550 nm處讀取OD。
海藻糖酶活性測定:用移液槍吸取待測提取液100 μL于10 mL離心管中,加入200 μL海藻糖標準溶液(40 mmol/L),于37 ℃條件下水浴30 min,然后沸水浴2~3 min終止反應,冰浴冷卻后加入上述顯色劑,并按照上述方法測定葡萄糖含量。該葡萄糖為海藻糖酶催化分解海藻糖所形成的葡萄糖。海藻糖酶活性以mmol/L(葡萄糖)·mg(蛋白)·min表示。
1. 2. 2. 3 葡萄糖含量測定 按試劑盒(南京建成生物工程研究所)操作方法進行。
1. 2. 2. 4 蛋白質含量測定 以牛血清蛋白(BSA,Sigma)為標準蛋白,采用考馬斯亮藍G-250染色法(Bradford,1976)進行測定。松突圓蚧蛋白質含量單位為mg/100只。
2 結果與分析
2. 1 不同處理對松突圓蚧海藻糖含量的影響
處理后逐日取樣測定松突圓蚧體內海藻糖含量,結果如圖1。由圖1可知,對照組松突圓蚧體內的海藻糖含量無較大波動,在0.6924~0.7286 mmol/L小幅度變化;而處理組各時間點所測海藻糖含量均小于對照組,且隨處理時間延長呈明顯的下降趨勢,在染菌后1~3 d海藻糖含量急速下降,第4 d略有升高但相較第3 d無顯著差異(P>0.05,下同),隨后緩慢下降,并在第7 d達最低點,為0.1702 mmol/L,與對照組相比降低了76.53%,兩者間差異達極顯著水平(P<0.01,下同)。
2. 2 不同處理對松突圓蚧海藻糖酶活性的影響
處理后逐日取樣測定松突圓蚧體內海藻糖酶活性,結果如圖2。由圖2可知,松突圓蚧體內的海藻糖酶活性變化趨勢與海藻糖含量變化趨勢相反,染菌后海藻糖酶活性逐漸升高,在染菌后3 d海藻糖酶活性(1.6570 mmol/L·min·mg)顯著高于對照組(P<0.05,下同);染菌后4 d有所降低,但與對照組差異不顯著;隨后又開始上升,并在第7 d時達最高點(1.7173 mmol/L·min·mg),顯著高于對照組,而在第6和8 d時與對照組相比達極顯著差異水平。
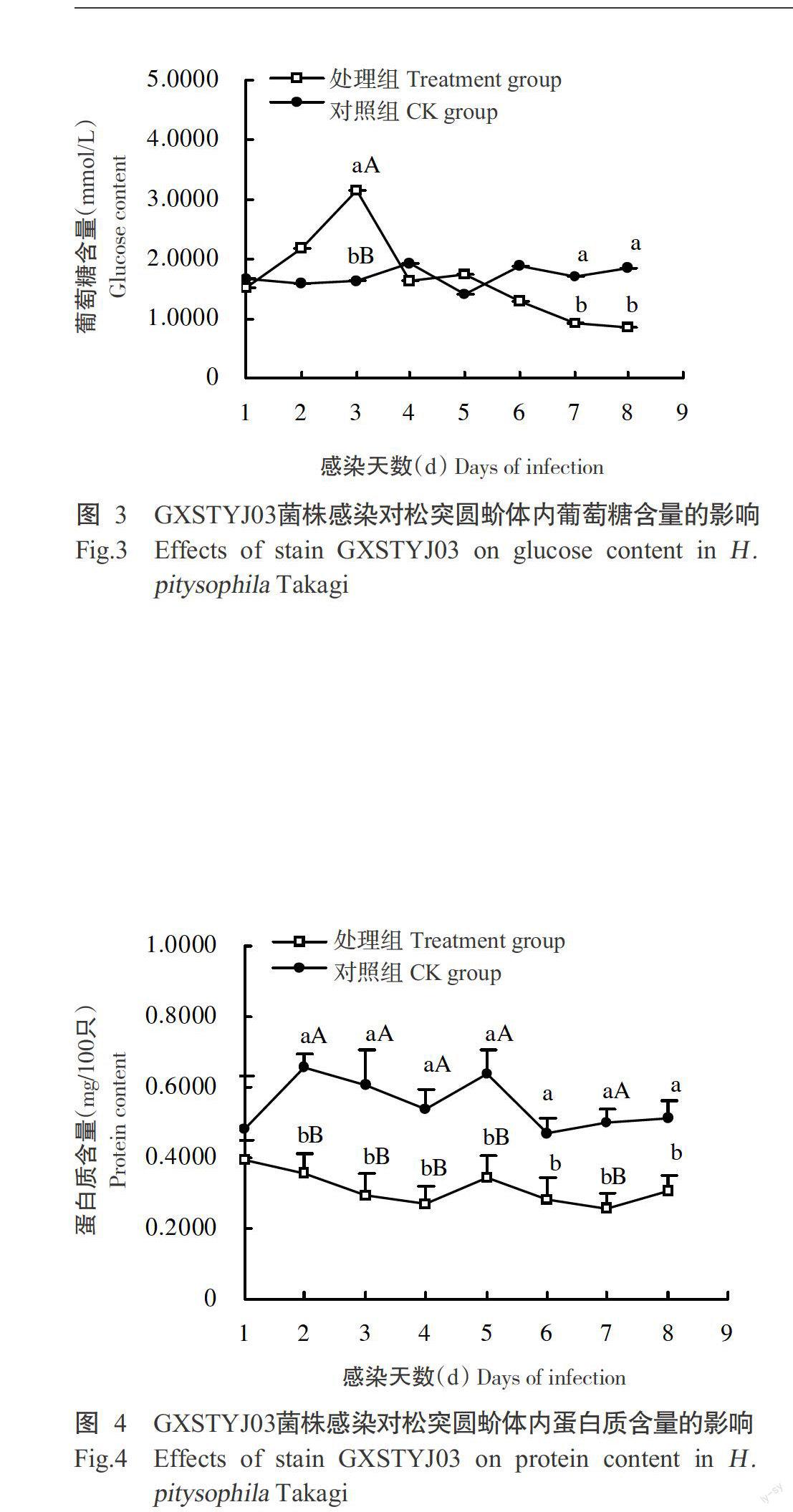
2. 3 不同處理對松突圓蚧葡萄糖含量的影響
對松突圓蚧體內葡萄糖含量的測定結果(圖3)顯示,對照組蟲體內的葡萄糖含量在試驗期內無明顯變化,維持在1.6741~1.8606 mmol/L;而處理組松突圓蚧體內的葡萄糖含量變化明顯,在感染初期葡萄糖含量上升,在染菌后3 d迅速升高至3.1479 mmol/L,是對照組的1.92倍,且差異達極顯著水平,隨后開始下降,在4~6 d時與對照組無顯著差異,7 d后處理組葡萄糖含量顯著低于對照組。
2. 4 不同處理對松突圓蚧蛋白質含量的影響
由圖4可看出,處理組松突圓蚧體內的蛋白質含量在試驗期內均低于對照組,在第7 d時降到最低值0.2581 mg/100只,只有對照組的51.41%,且處理后2~5 d和第7 d時蛋白質含量與對照組相比差異達極顯著水平。
3 討論
昆蟲細胞內海藻糖含量較高時能保護細胞活性,增強昆蟲生存和防御傷害的能力,在抵御沖擊負荷等方面有重要作用(雷芳等,2006;陸玉建等,2015)。病原真菌在侵染寄主過程中,菌絲生長所需能量主要來源于寄主。本研究結果顯示,松突圓蚧被真菌侵染3 d后,其體內海藻酶活性迅速升高,且一直高于對照組,反之海藻糖含量迅速降低且一直低于對照組;隨著海藻糖被海藻糖酶降解,在第1~3 d時葡萄糖含量顯著升高,為真菌菌絲生長和繁殖提供了能量,在染菌3 d后葡萄糖含量開始下降,且在5 d后明顯低于對照組,說明病菌突破了昆蟲的抵御防線,使得昆蟲疾病加重逐漸進入死亡階段。菌絲除了直接消耗寄主體內的葡萄糖外,也有可能利用自身的海藻糖酶分解寄主體內的海藻糖,從而促使寄主提高海藻糖酶活性來分解海藻糖獲取能量,因此導致處理組的海藻糖酶活性始終高于對照組,海藻糖含量則低于對照組,該變化對松突圓蚧的取食行為和免疫能力有極大影響,與樊金華等(2013)研究感染布氏白僵菌后油松毛蟲血淋巴中海藻糖酶活性、海藻糖和葡萄糖含量的變化結果相似。
隨著真菌菌絲在寄主體內生長和大量繁殖,寄主體內海藻糖和葡萄糖被大量消耗,蟲體為了維持自身的生理活動,極有可能分解蛋白質。菌絲在侵染蟲體時也會釋放蛋白酶(黃蓉等,2014)分解寄主蛋白質從而貫穿于蟲體各組織,因此本研究處理組幼蟲的蛋白質含量在染菌2 d后即顯著低于對照組。總之,正是由于病原真菌侵染蟲體導致蟲體內海藻糖酶活性增強將海藻糖轉化為葡萄糖,這一過程既為真菌提供了營養而有利于病原真菌生長和提高感染力,同時又破壞蟲體內血糖平衡,從而加速昆蟲死亡。
據報道,某些真菌具有海藻糖酶,其分為酸性海藻糖酶和中性海藻糖酶,本研究中蟲生廣布擬盤多毛孢GXSTYJ03菌株在侵染松突圓蚧時是否利用海藻糖酶及是何種海藻糖酶在起作用有待進一步探究;同時,蟲生廣布擬盤多毛孢對松突圓蚧體內保護酶和解毒酶等相關防御酶系的影響亦有待后續研究。
4 結論
本研究結果表明,蟲生廣布擬盤多毛孢侵染松突圓蚧過程中引起蟲體內海藻糖、葡萄糖和蛋白質含量一定程度降低,導致其無法供給維持自身生命活動所需的能量,是致死松突圓蚧的原因之一。
參考文獻:
樊金華,謝映平,薛皎亮,張海濤. 2013. 感染布氏白僵菌后油松毛蟲血淋巴中海藻糖酶活性、海藻糖和葡萄糖含量的變化[J]. 昆蟲學報,56(8):864-869. [Fan J H,Xie Y P,Xue J L,Zhang H T. 2013. Changes of trehalase activity and contents of trehalose and glucose in the hemolymph of Dendrolimus tabulaeformis(Lepidoptera:Lasiocampidae) larvae infected with Beauveria brongniartii[J]. Acta Entomologica Sinica,56(8):864-869.]
方麗英,黃寶靈,王緝健,李貴玉,李瑞龍,呂成群. 2007. 松突圓蚧生防菌的毒力試驗[J].中國森林病蟲,26(6): 25-27. [Fang L Y,Huang B L,Wang J J,Li G Y,Li R L,Lü C Q. 2007. Toxicity experiment of biocontrol fungus isolated from emiberlesia pitysophila[J]. Forest Pest and Disease,26(6):25-27.]
馮慧. 1989. 昆蟲生物化學分析法[M]. 北京:農業出版社:9-10. [Feng H. 1989. Biochemical Analysis Method for Insect[M]. Beijing:Agriculture Press:9-10.]
郭麗娟,黃寶靈,呂成群,黃蓉,李瑞龍,何錦華,何鈺,盧強,黃艷紅,任涵. 2014. 不同環境條件對蟲生廣布擬盤多毛孢退化的影響[J]. 廣西林業科學,43(2):147-150. [Guo L J,Huang B L,Lü C Q,Huang R,Li R L,He J H,He Y,Lu Q,Huang Y H,Ren H. 2014. Effect of different environmental factors on degeneration of Entomogenous Pestalotiopsis disseminate[J]. Guangxi Forestry Science,43(2):147-150.]
黃寶靈,丁波,方麗英,宋力,陳炳雄,李健榮,王清. 2009. 松突圓蚧生防菌粉劑的林間擴散能力[J]. 中國生物防治,25(S1):24-28. [Huang B L, Ding B, Fang L Y,Song L,Chen B X,Li J R,Wang Q. 2009. The diffusion ability of biocontrol fungi pulvis of Hemiberlesia pitysophila Takagi in forest[J]. Chinese Journal of Biological Control,25(S1):24-28.]
黃蓉,呂成群,黃寶靈,郭麗娟,姚姜銘,李瑞龍,何錦華,何鈺,盧強. 2014. 蟲生廣布擬盤多毛孢蛋白酶、幾丁質酶、脂肪酶產生水平及其毒力關系研究[J]. 南方農業學報,45(7):1172-1177. [Huang R,Lü C Q,Huang B L,Guo L J,Yao J M,Li R L,He J H,He Y,Lu Q. 2014. Relationships between virulence and activities of protease, chitinase and lipase produced by entomogenous Pestalotiopsis disseminate[J]. Journal of Southern Agriculture,45(7):1172-1177.]
雷芳,張桂芬,萬方浩,馬俊. 2006. 寄主轉換對B型煙粉虱和溫室粉虱海藻糖含量和海藻糖酶活性的影響[J]. 中國農業科學,39(7):1387-1394. [Lei F,Zhang G F,Wan F H,Ma J. 2006. Effects of plant species switching on contents and dynamics of trehalose and trehalase activity of Bemisia tabaci B-biotype and Trialeurodes vaporariorum[J]. Scientia Agricultrua Scinca,39(7):1387-1394.]
陸玉建,張韓杰,劉南南. 2015. 大腸桿菌otsA基因的克隆和轉化本生煙草[J]. 江蘇農業學報,31(1):32-38. [Lu Y J,Zhang H J,Liu N N. 2015. Cloning of otsA gene in Escherichia coli and its transformation into Nicotiana benthamiana[J]. Jiangsu Journal of Agricultural Sciences,31(1):32-38.]
呂成群,黃寶靈,任涵,李文祿,黃蓉,郭麗娟. 2014. 蟲生廣布擬盤多毛孢GH10菌株對松突圓蚧的致病力測定[J]. 南方農業學報,45(6):980-983. [Lü C Q,Huang B L,Ren H,Li W L,Huang R,Guo L J. 2014. Virulence of Pestalotiopsis disseminata GH10 strain to Hemiberlesia pitysophila[J]. Journal of Southern Agricultrue,45(6):980-983.]
熊琦. 2013. 桃小食心蟲病原真菌——球孢白僵菌TST05菌株的研究[D]. 太原:山西大學. [Xiong Q. 2013. Study on Beauveria bassiana strain TST05-An entomopathogenic fungus for Carposina sasakii[D]. Taiyuan: Shanxi University.]
薛皎亮,牛宇,謝映平. 2006. 油松毛蟲感染白僵菌后體內蛋白質、酯酶和多酚氧化酶的變化[J]. 應用與環境生物學報,12(6):814-818. [Xue J L,Niu Y,Xie Y P. 2006. Changes of protein,esterase and PPO in Dentrolinus tabulaefomis infected by Beauveria bassiana[J]. Chinese Journal of Applied & Environmental Biology,12(6):814-818.]
于彩虹,盧丹,林榮華,王曉軍,姜輝. 2008. 海藻糖——昆蟲的血糖[J]. 昆蟲知識,45(5):832-836. [Yu C H, Lu D, Lin R H, Wang X J, Jiang H. 2008. Trehalose—The blood sugar in insects[J]. Chinese Bulletin of Entomology,45(5):832-836.]
Bradford M M. 1976. A rapid and sensitive method for the quantification of microgram quantities of protein using the principle of protein-dye binding[J]. Analytical Biochemistry, 72: 248-254.
Lü C Q,Huang B L,Qiao M J,Wei J G,Ding B. 2011. Entomopathogenic fungi on Hemiberlesia pitysophila[J]. PLoS one,6(8):1-6.
Takagi S. 1969. A new species of Chionaspis from Florida(Homoptera:Coccoidea)[J]. Kontyu,37(3):269-271.
Takagi S,Tang F T. 1982. A new scale insect of the Quernaspis group(Homoptera, Coccoidea,Diaspididae) from China[J]. Kontyu,50(1):100-103.
(責任編輯 麻小燕)
