玉米抗病毒病基因工程研究進(jìn)展
2017-05-30 10:48:04燕照玲段俊枝馮麗麗陳海燕齊紅志楊翠蘋施艷任銀玲劉毓俠
南方農(nóng)業(yè)學(xué)報(bào) 2017年12期
燕照玲 段俊枝 馮麗麗 陳海燕 齊紅志 楊翠蘋 施艷 任銀玲 劉毓俠
摘要:病毒病是導(dǎo)致玉米產(chǎn)量降低和品質(zhì)下降的主要原因之一。在我國,玉米矮花葉病和粗縮病發(fā)生范圍最廣、危害最嚴(yán)重。基因工程技術(shù)可人為將抗性基因或部分片段定向?qū)酥参铽@得轉(zhuǎn)基因抗病毒植株,具有速度快、效率高等優(yōu)點(diǎn),在玉米抗病毒育種中具有重要的應(yīng)用價(jià)值。文章在總結(jié)我國主要玉米病毒病及其病原種類的基礎(chǔ)上,論述采用不同策略培育抗病毒玉米植株的研究進(jìn)展,其中,利用植物病毒基因序列的策略有病毒編碼蛋白基因介導(dǎo)的抗病性、RNA干擾(RNAi)介導(dǎo)的抗病性、人工小RNA(amiRNA)介導(dǎo)的抗病性3種,還可利用非植物病毒基因,包括寄主的抗性基因及來自其他植物、動(dòng)物和微生物的抗病毒基因如核糖體失活蛋白、核酸酶、2-5A體系的基因等;最后分析各種策略的優(yōu)缺點(diǎn)及抗病毒轉(zhuǎn)基因玉米的安全性問題,為科研工作者優(yōu)化玉米抗病毒育種工程、培育生產(chǎn)上可推廣的抗病毒品種提供參考。
關(guān)鍵詞:玉米;病毒病;甘蔗花葉病毒;水稻黑條矮縮病毒;基因工程;抗病育種
中圖分類號:S435.131 文獻(xiàn)標(biāo)志碼:A 文章編號:2095-1191(2017)12-2136-09低和品質(zhì)下降的主要原因之一,且呈發(fā)病面積逐年擴(kuò)大、病情逐年加重的趨勢,絕產(chǎn)情況時(shí)有發(fā)生。由于生產(chǎn)上缺乏行之有效的藥劑和防治措施,病毒病已成為限制玉米持續(xù)增產(chǎn)的主要障礙。培育、推廣抗病毒品種并輔以合理的栽培管理措施是目前公認(rèn)防治病毒病的最佳途徑,但玉米種質(zhì)資源對病毒病抗性的遺傳基礎(chǔ)較復(fù)雜,進(jìn)而給通過傳統(tǒng)雜交利用玉米抗性基因培育抗病品種帶來一定難度。隨著組織培養(yǎng)和基因工程技術(shù)的發(fā)展,可人為將抗性基因或部分片段定向?qū)胫参铽@得轉(zhuǎn)基因抗病毒植株,該技術(shù)具有速度快、效率高等優(yōu)點(diǎn),在玉米抗病毒育種中的應(yīng)用研究已取得了重大進(jìn)展。本文通過介紹我國主要的玉米病毒病種類及其病原,重點(diǎn)論述采用不同策略培育抗病毒玉米植株研究進(jìn)展,為科研工作者優(yōu)化玉米抗病毒育種工程提供參考。
1我國玉米病毒病主要種類及其病原
玉米在其整個(gè)生長期均可被病毒感染,產(chǎn)生葉片褪綠、枯斑、雌雄穗不育、植株矮化、壞死或直接死亡等癥狀,最終導(dǎo)致產(chǎn)量降低、品質(zhì)下降。據(jù)不完全統(tǒng)計(jì),全世界有40多種玉米病毒病,在我國發(fā)生的主要有粗縮病、矮花葉病、條紋矮縮病、紅葉病和鼠耳病等,其中矮花葉病和粗縮病發(fā)生范圍最廣、危害最嚴(yán)重(李秀坤等,2015),因此,這兩種病毒病是我國科研工作者研究的重點(diǎn)。
1.1玉米矮花葉病及其病原
矮花葉病是世界玉米產(chǎn)區(qū)普遍發(fā)生的一種病毒病,1963年首先在美國俄亥俄州被發(fā)現(xiàn)。我國于1968年首次在河南新鄉(xiāng)和安陽地區(qū)發(fā)現(xiàn)該病,隨后華北各省(市)陸續(xù)發(fā)生,目前在東北、西南及西北各省(區(qū))均有報(bào)道。矮花葉病已成為我國玉米產(chǎn)區(qū)的主要病害之一。發(fā)病植株通常葉片不均勻褪綠,形成花葉、條紋癥狀,病情嚴(yán)重時(shí)植株矮化,雌雄穗發(fā)育受抑制,最終導(dǎo)致玉米產(chǎn)量和質(zhì)量受到嚴(yán)重影響。此外,玉米矮花葉病明顯表現(xiàn)出暴發(fā)性、遷移性和間歇性三大特征,每年給玉米生產(chǎn)造成20%~80%的損失(張超等,2017)。
玉米矮花葉病由一至多種病毒系統(tǒng)性侵染引起,國際上已報(bào)道的病毒有6種,均為馬鈴薯Y病毒屬(Potyvirus)成員,分別是玉米矮花葉病毒(Maize dwarf mosaic virus,MDMV)、甘蔗花葉病毒(Sugar-cane mosaic virus,SCMV)、玉米屬花葉病毒(Zea mosaic virus,ZeMV)、高梁花葉病毒(Sorghum mo-saic virus,SrMV)、約翰遜草花葉病毒(Johnson-grass mosaic virus,JGMV)和白草花葉病毒(Penni-setum mosaic virus,PenMV)。據(jù)報(bào)道,我國玉米矮花葉病的病原有SCMV(曾被認(rèn)為是MDMV-B或SC-MV-MDB)(蔣軍喜等,2003)和PenMV(曾被命名為MDMV-G)(Fan et al.,2003),目前SCMV發(fā)生普遍,而PenMV的發(fā)生較少,且多限于山西省和河北省(張超等,2017)。各病毒基因組均為單分子正義單鏈RNA(ssRNA),全長約10 kb,核酸5′端連接一個(gè)基因組連接蛋白(VPg),3′端具有一個(gè)poly(A)尾,中間是一個(gè)大的開放閱讀框,通過多聚蛋白加工和移碼翻譯策略產(chǎn)生11個(gè)具有不同功能的成熟蛋白,從N端到C端依次為第一蛋白(P1)、輔助成分—蛋白酶(HC-Pro)、第三蛋白(P3)、P3N-PIPO(位于P3閱讀框內(nèi)部)、第一個(gè)6K蛋白(6K1)、圓柱狀內(nèi)含體蛋白(CI)、第二個(gè)6K蛋白(6K2)、VPg、核內(nèi)含體蛋白a-蛋白酶(NIa-Pro)、核內(nèi)含體蛋白b(NIb)及外殼蛋白(CP)(Revers and Garcia,2015)。
1.2玉米粗縮病及其病原
粗縮病也是玉米生產(chǎn)上的主要病毒病害,具有暴發(fā)性、流行性和毀滅性的特點(diǎn)。該病于1949年首次在意大利種植的美國玉米材料中被發(fā)現(xiàn),目前遍布世界大多數(shù)玉米產(chǎn)區(qū)。在我國,粗縮病于20世紀(jì)50年代首次在新疆和甘肅被發(fā)現(xiàn),70年代和90年代一度在華北和西北等地區(qū)暴發(fā)。近年來,由于氣候變化及種植結(jié)構(gòu)調(diào)整,粗縮病在我國的發(fā)生逐漸加重,特別是在黃淮海夏玉米區(qū)套播或晚春播、早夏播玉米上危害嚴(yán)重,其中山東、江蘇、遼寧、安徽和河南等省的玉米生產(chǎn)均因粗縮病暴發(fā)而遭受重大損失。玉米幼苗期感病,生長延緩,節(jié)問變得粗短,植株嚴(yán)重矮化,同時(shí)葉背出現(xiàn)白色蠟淚狀脈突,葉片變短、僵直、表現(xiàn)深綠;生育后期植株常不能抽穗,或穗異常、結(jié)實(shí)少,嚴(yán)重降低產(chǎn)量(張曉婷等,2011)。
目前已報(bào)道能引起玉米粗縮病的病毒主要有4種,即玉米粗縮病毒(Maize rough dwarf virus,MRDV)、馬德里約祠托病毒(Mal de Rio Cuarto virus,MRCV)、水稻黑條矮縮病毒毒(Rice black-streaked dwarf virus,RBSDV)和南方水稻黑條矮縮病毒(Southern rice black-streaked dwarf virus,SRBSDV),均屬于斐濟(jì)病毒屬(Fijivirus)(張曉婷等,2011)。美國和歐洲的玉米粗縮病病原主要是MRDV;MRCV主要分布于南美洲;我國玉米粗縮病的病原主要是RBSDV和SRBSDV,其中RBSDV在全國普遍發(fā)生,尤其是北方玉米種植區(qū),其傳播介體為灰飛虱,而SRBSDV主要在南方發(fā)生,由白背飛虱傳播(章松柏等,2013)。RBSDV基因組由10條雙鏈RNA(dsRNA)組成,總長29142 bp,根據(jù)片段由大到小依次命名為S1~S10。其中,S1編碼RNA依賴RNA聚合酶,S2編碼主要核心結(jié)構(gòu)蛋白,S8和s10分別編碼核心衣殼蛋白和外層衣殼蛋白(鄧金奇和戴良英,2012)。SRB-SDV是周國輝等(2010)在我國南方地區(qū)水稻上發(fā)現(xiàn)的新病毒,后來發(fā)現(xiàn)其也侵染玉米,引發(fā)典型的粗縮病癥狀,其基因組結(jié)構(gòu)與RBSDV相似,但在序列上存在明顯差異,各片段核苷酸序列相似性均低于80%。
2玉米抗病毒基因工程
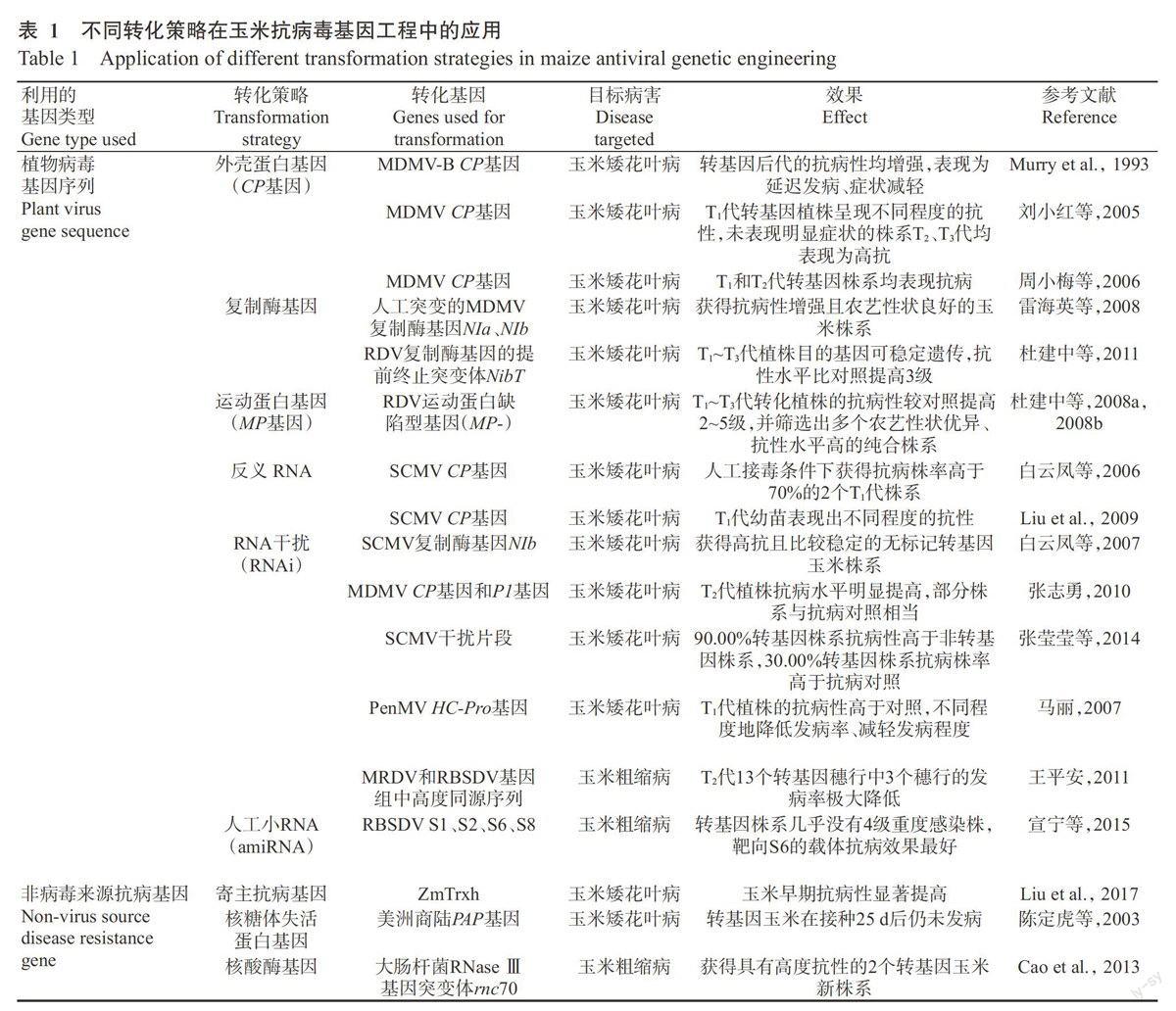
在玉米抗病毒基因工程研究中,用于轉(zhuǎn)化的基因可分為兩類:一類是植物病毒的基因序列,如CP基因、復(fù)制酶基因、運(yùn)動(dòng)蛋白(MP)基因及反義RNA等;另一類是非植物病毒基因,包括來自植物、動(dòng)物和微生物的抗病毒基因及寄主的抗性基因等。如表1所示,國內(nèi)外學(xué)者通過將兩類基因轉(zhuǎn)化玉米獲得大量陽性植株,且大多數(shù)植株的抗病性提高。
2.1利用植物病毒基因序列
自Sanford和Johnston(1985)提出來源于病原抗性的概念后,利用病毒來源的基因轉(zhuǎn)化植物逐漸成為抗病毒基因工程的重要途徑。第1代抗病毒轉(zhuǎn)基因策略主要通過表達(dá)病毒自身蛋白基因來實(shí)現(xiàn)。第2代抗病毒轉(zhuǎn)基因策略基于RNAi技術(shù),通常是將與病毒基因同源的dsRNA或發(fā)卡RNA(Hairpin RNA,hpRNA)轉(zhuǎn)入植物體內(nèi),產(chǎn)生病毒特異性的小干擾RNA(Small interfering RNA,siRNA)后,引起入侵的同源病毒基因發(fā)生沉默。第3代抗病毒轉(zhuǎn)基因策略是基于amiRNA干擾技術(shù),能有效避免脫靶效應(yīng)和重組病毒的產(chǎn)生,具有抗性水平高、遺傳穩(wěn)定性強(qiáng)、生物安全性高等優(yōu)點(diǎn)(FaNm and LarMn,2013)。
2.1.1第1代抗病毒轉(zhuǎn)基因策略 該策略主要是將植物病毒編碼蛋白的基因轉(zhuǎn)入植物細(xì)胞,其表達(dá)后可使轉(zhuǎn)基因植株獲得抗病毒能力。玉米抗病毒基因工程研究中利用的病毒基因主要包括CP基因、復(fù)制酶基因、MP基因及反義RNA,轉(zhuǎn)化植株后均取得較好的抗病效果,其中CP基因研究得最早,應(yīng)用也最多。
(1)CP是形成病毒顆粒的結(jié)構(gòu)蛋白,其主要功能是包被病毒基因組核酸及參與病毒的長距離運(yùn)輸。CP基因介導(dǎo)的抗性機(jī)制較復(fù)雜,早期研究認(rèn)為是由于CP蛋白的積累封閉病毒粒子脫殼,從而阻礙病毒侵染,后來逐漸發(fā)現(xiàn)存在著不依賴于CP蛋白本身的抗性機(jī)制,因而抗性水平也可能發(fā)生在RNA水平上(Reimann-Philipp and Beachy,1993)。Murry等(1993)首次報(bào)道,通過將MDMV-B的CP基因?qū)胗衩椎玫娇共≈仓辏斯そ臃NMDMV-A株系、MDMV-B株系鑒定,轉(zhuǎn)基因后代的抗病性均較非轉(zhuǎn)基因?qū)φ赵鰪?qiáng),表現(xiàn)為延遲發(fā)病、癥狀減輕。之后CP基因介導(dǎo)的抗玉米病毒病策略得到科學(xué)界的廣泛關(guān)注,國內(nèi)也進(jìn)行了積極探索。劉小紅等(2005)采用MD-MV CP基因轉(zhuǎn)化玉米優(yōu)良自交系18-599紅和18-599白,人工接種病毒后,對照植株全部表現(xiàn)出玉米矮花葉病的典型癥狀,而T1代轉(zhuǎn)基因植株呈現(xiàn)不同程度的抗性,其中有4個(gè)植株未表現(xiàn)明顯癥狀;對這4個(gè)株系后代進(jìn)一步接種病毒鑒定發(fā)現(xiàn),T2和T3代均表現(xiàn)為高抗玉米矮花葉病,表明轉(zhuǎn)MDMV CP基因獲得的抗病性能穩(wěn)定遺傳。周小梅等(2006)在玉米自交系綜3和綜31中轉(zhuǎn)入MDMV CP基因,也獲得了可育轉(zhuǎn)基因植株。在大田抗病性鑒定試驗(yàn)中,綜31的T1和T2代轉(zhuǎn)基因株系均表現(xiàn)出抗玉米矮花葉病,大部分植株的發(fā)病率和病情指數(shù)低于對照(T1代植株比例分別為83%和67%,T2代比例為100%和90%);溫室抗病性鑒定試驗(yàn)中,綜3的18個(gè)T1代轉(zhuǎn)基因株系有7個(gè)抗MDMV侵染,不僅發(fā)病延遲,其株高和穗形成率還明顯高于對照。接種病毒后,綜3對照的可溶性糖含量和超氧化物歧化酶(SOD)、苯丙氨酸解氨酶(PAL)、過氧化物酶(POD)活性均下降,而轉(zhuǎn)基因植株體內(nèi)的可溶性糖含量有所上升,SOD、PAL活性變化不大,POD活性高于對照,過氧化氫酶(CAT)活性變化也存在差異(韓玉杰,2009)。
(2)復(fù)制酶是催化病毒合成正、負(fù)鏈RNA的RNA聚合酶。復(fù)制酶基因介導(dǎo)的抗病性機(jī)制可能是轉(zhuǎn)基因表達(dá)的復(fù)制酶反向調(diào)控病毒復(fù)制,使其復(fù)制效率降低,如果表達(dá)缺陷型復(fù)制酶,其還可能與野生型病毒復(fù)制酶產(chǎn)生競爭而干擾病毒復(fù)制(Carr et al.,1992;杜建中等,2011)。張志燕(2004)將RBSDVS1部分片段轉(zhuǎn)化玉米,獲得了可育再生植株,在T1代植株中檢測到外源基因,并且植株形態(tài)和農(nóng)藝性狀表現(xiàn)正常,為篩選抗粗縮病玉米株系奠定了基礎(chǔ)。雷海英等(2008)將人工突變的MDMV復(fù)制酶基因(NIa、NIb)轉(zhuǎn)入4種不同玉米自交系中,對轉(zhuǎn)化的植株連續(xù)2代(T1和T2)利用人工摩擦接種病毒的方法進(jìn)行田問抗病性鑒定,獲得了對玉米矮花葉病抗病性增強(qiáng)且農(nóng)藝性狀良好的玉米株系。從T2代整體的發(fā)病率來看,4種材料與其相應(yīng)的對照材料相比均表現(xiàn)出較高的抗性,其中金黃96B、478兩種轉(zhuǎn)基因材料較金黃96C、C649兩種材料的抗性強(qiáng),可能與玉米材料、轉(zhuǎn)化基因及外源基因整合位點(diǎn)等不同有關(guān)。杜建中等(2011)將水稻矮縮病毒(Rice dwarf virus,RDV)復(fù)制酶基因的提前終止突變體(NibT)導(dǎo)人玉米自交系金黃96B,對T1~T3代轉(zhuǎn)基因植株進(jìn)行鑒定,發(fā)現(xiàn)目的基因可穩(wěn)定遺傳,田問對玉米矮花葉病的抗性水平各代基本一致,比對照提高3級,同時(shí)穗長、穗粒數(shù)、百粒質(zhì)量等農(nóng)藝性狀也有所改善。而林美娟等(2005)認(rèn)為復(fù)制酶基因介導(dǎo)的抗病性比CP基因介導(dǎo)的抗病性更有效,然而由于該策略對植物病毒種類具有較高的特異性,因此其應(yīng)用范圍可能會(huì)受限。
(3)MP介導(dǎo)的病毒抗性策略一般采用缺陷型MP。抗病機(jī)理可能表現(xiàn)在蛋白質(zhì)水平上,轉(zhuǎn)基因所表達(dá)的缺陷型MP與入侵病毒的正常MP競爭寄主胞問連絲上的結(jié)合位點(diǎn),阻礙病毒在細(xì)胞間的移動(dòng)(王關(guān)林和方洪筠,2002)。Carrington等(1996)研究認(rèn)為,許多不同病毒的MP可能與相同的胞間連絲成分發(fā)生作用,因而該策略的抗病效果具有廣譜高效性。杜建中等(2008a,2008b)將RDV運(yùn)動(dòng)蛋白缺陷型基因(MP-)導(dǎo)入玉米自交系478,田間接種MDMV進(jìn)行調(diào)查,結(jié)果發(fā)現(xiàn)轉(zhuǎn)化植株的抗矮花葉病能力與對照植株相比有顯著提高,個(gè)別株系在T1代的發(fā)病率為0,T1~T3代轉(zhuǎn)化植株的抗病性逐漸提高,較對照提高2~5級。結(jié)合對優(yōu)異農(nóng)藝性狀的篩選,獲得96C0502、96C0507和96C0513等抗矮花葉病轉(zhuǎn)基因玉米純合株系,這些株系連續(xù)2年的發(fā)病率均為0,抗病性比對照提高4級。以上研究表明,利用病毒MP-基因轉(zhuǎn)化玉米是獲得抗矮花葉病玉米株系的快捷途徑。進(jìn)一步對RDV MP-轉(zhuǎn)基因玉米的生理特性進(jìn)行研究,結(jié)果發(fā)現(xiàn)轉(zhuǎn)基因改變了玉米同工酶的酶譜,第1真葉中POD、CAT和SOD的活性顯著高于非轉(zhuǎn)基因玉米,表明該基因?qū)肽軌蚣涌焓荏w基因型變異速率,從而增強(qiáng)其抗逆能力和適應(yīng)性(燕平梅等,2012)。
(4)反義RNA是一類與mRNA互補(bǔ)的ssRNA,其通過與靶向mRNA配對產(chǎn)生dsRNA,阻止mRNA翻譯成蛋白質(zhì)或?qū)е耺RNA降解,從而阻斷病毒侵染(牙庫甫江·阿西木等,2015)。Liu等(2009)將SCMVCP基因以反義方向?qū)擞衩鬃越幌?8-599紅,對T1代種子長出的幼苗接種SCMV-MDB,發(fā)現(xiàn)其對病毒表現(xiàn)出不同程度的抗性。白云鳳等(2006)將SCMV反義CP基因?qū)胗衩鬃越幌稻C3,人工接毒條件下獲得抗病株率高于70%的2個(gè)T1代株系,與轉(zhuǎn)MD-MV或SCMV CP基因獲得的結(jié)果相比,轉(zhuǎn)反義CP基因后代的抗病株率高、抗病性強(qiáng),說明轉(zhuǎn)反義CP基因也是玉米獲得矮花葉病抗性的一條有效途徑。反義RNA不會(huì)翻譯成蛋白質(zhì)與入侵病毒進(jìn)行異源包裝,也不會(huì)與入侵病毒發(fā)生遺傳物質(zhì)重組,因而該策略與正義RNA相比具有更高的生物安全性。
2.1.2第2代抗病毒轉(zhuǎn)基因策略 RNAi是植物體內(nèi)天然存在的抗病毒機(jī)制。基于RNAi的抗病毒轉(zhuǎn)基因策略是人為強(qiáng)化植物的這種天然機(jī)制,選擇病毒基因組中的某段序列設(shè)計(jì)成雙鏈結(jié)構(gòu)轉(zhuǎn)入植物進(jìn)行表達(dá),其會(huì)被植物體內(nèi)的Dicer(一種RNA酶)降解成21~25 nt的siRNA,然后siRNA與植物的AGO蛋白結(jié)合形成RNA誘導(dǎo)的沉默復(fù)合體(RNA-induced si-lencing complex,RISC),其在siRNA指導(dǎo)下將互補(bǔ)的病毒RNA降解,從而阻止病毒入侵(牙庫甫江·阿西木等,2015)。RNAi介導(dǎo)的抗病性策略特異性強(qiáng),較易獲得抗性高的植株,且抗病性能穩(wěn)定遺傳,同時(shí)避免病毒蛋白翻譯、病毒RNA重組及異源包裝等潛在風(fēng)險(xiǎn),與傳統(tǒng)抗病毒基因工程相比具有更高的安全性(白云鳳等,2007)。因此,RNAi技術(shù)為培育抗病毒植株提供了一條高效途徑,成為玉米抗病毒基因工程研究的重要策略。
白云鳳等(2007)構(gòu)建SCMV復(fù)制酶基因(NIb)的反向重復(fù)序列表達(dá)載體并轉(zhuǎn)化玉米自交系,對T1和T2代植株接種病毒進(jìn)行鑒定,獲得對SCMV高抗且較穩(wěn)定的無標(biāo)記轉(zhuǎn)基因玉米株系,同時(shí)發(fā)現(xiàn)其抗病株率和抗病程度均高于傳統(tǒng)的轉(zhuǎn)正義基因和反義基因。張志勇(2010)分別選取MDMV CP基因和P1基因的保守序列構(gòu)建RNAi表達(dá)載體,之后轉(zhuǎn)化玉米自交系18-599,對T2代轉(zhuǎn)基因植株接種病毒發(fā)現(xiàn),其抗病水平明顯提高,部分株系與抗病對照H9-21相當(dāng)。河南農(nóng)業(yè)大學(xué)玉米種質(zhì)資源創(chuàng)新與高產(chǎn)機(jī)制研究課題組將人工合成的SCMV干擾片段構(gòu)建成RNAi載體,以HiⅡ、HiⅡA、HiⅡB、H99的幼胚為受體材料進(jìn)行轉(zhuǎn)化獲得了轉(zhuǎn)基因株系,對T1和T2代接種SCMV進(jìn)行鑒定發(fā)現(xiàn),90.00%轉(zhuǎn)基因株系抗病性高于非轉(zhuǎn)基因株系,30.00%轉(zhuǎn)基因株系抗病株率高于抗病對照黃早4,轉(zhuǎn)基因株系的抗病性明顯提高,說明利用RNAi技術(shù)培育抗矮花葉病的轉(zhuǎn)基因玉米株系具有可行性(張瑩瑩等,2014)。對于PenMV引起的玉米矮花葉病,馬麗(2007)也嘗試采用RNAi策略進(jìn)行抗病基因工程研究,其根據(jù)PenMV HC-Pro基因保守序列設(shè)計(jì)引物構(gòu)建了反向重復(fù)序列表達(dá)載體并轉(zhuǎn)化玉米,經(jīng)人工接種PenMV鑒定,轉(zhuǎn)基因T1代植株的抗病性高于對照,表現(xiàn)為發(fā)病率降低、發(fā)病程度減輕。
抗玉米粗縮病研究中,郭靖(2011)分別選擇MRDV S10和RBSDVS10、S8保守區(qū)域中的303、452和291 bp片段作為RNAi靶序列,構(gòu)建成具有反向重復(fù)結(jié)構(gòu)的植物表達(dá)載體后轉(zhuǎn)化玉米自交系18-599,獲得了陽性轉(zhuǎn)化植株。王平安(2011)選取MRDV和RBSDV基因組中高度同源的一段558 bp序列作為干擾序列,轉(zhuǎn)化玉米HiⅡ幼胚后,對T2代采取人工接種飼毒灰飛虱方法進(jìn)行田間抗性鑒定,結(jié)果發(fā)現(xiàn)非轉(zhuǎn)基因穗行發(fā)病率為57.1%,13個(gè)轉(zhuǎn)基因穗行中3個(gè)穗行的發(fā)病率相對較低(0~15.8%),被初步鑒定為轉(zhuǎn)基因抗粗縮病株系。
為了獲得兼抗矮花葉病和粗縮病的轉(zhuǎn)基因玉米植株,可構(gòu)建同時(shí)含有2種病毒干擾片段的復(fù)合表達(dá)載體。王迅等(2010)分別將RBSDV S8、S10部分片段與SCMV CP基因的一段融合,通過正反向插入植物表達(dá)載體構(gòu)建2種病毒來源基因的RNAi載體,將其導(dǎo)人玉米自交系A(chǔ)188中,部分陽性轉(zhuǎn)基因植株對玉米矮花葉病具有明顯的抗性。甘德芳(2011)分別擴(kuò)增SCMV和MRDV的CP基因(MRDV為S10)特異性干擾片段,構(gòu)建成反向重復(fù)序列并串聯(lián)在一起得到RNAi復(fù)合表達(dá)載體,通過轉(zhuǎn)化玉米獲得了整合有2種病毒CP基因RNAi片段的轉(zhuǎn)化株。對T2代4葉期玉米接種SCMV后,不同轉(zhuǎn)化株系的抗病性存在差異,但均高于對照。至今,有關(guān)二價(jià)轉(zhuǎn)基因玉米植株對粗縮病的抗性尚無報(bào)道,下一步需要對獲得的轉(zhuǎn)基因材料進(jìn)行粗縮病抗性檢測,以驗(yàn)證采用RNAi策略抗多種病毒的效果及可行性。
2.1.3第3代抗病毒轉(zhuǎn)基因策略 amiRNA抗病毒策略是以植物內(nèi)源前體miRNA(Pre-miRNA)為骨架,將其莖環(huán)結(jié)構(gòu)中的miRNA序列替換為與病毒基因序列互補(bǔ)的amiRNA序列形成pre-amiRNA,其轉(zhuǎn)入植物后在內(nèi)源miRNA合成機(jī)制作用下生成靶向病毒mRNA的amiRNA,指導(dǎo)RISC降解病毒mRNA(Tiwari et al.,2014)。利用amiRNA策略培育抗病毒植株已在許多植物和病毒組合中被證明有效,其在玉米抗病育種方面的潛力也引起了廣泛的重視。宣寧等(2015)根據(jù)玉米zea-miR159a的前體序列和RBSDV基因組序列信息設(shè)計(jì)引物,構(gòu)建了用于沉默RBSDV S1、S2、S6、S8的amiRNA載體,之后轉(zhuǎn)化玉米自交系綜31,選擇miRNA表達(dá)量高的純合體株系進(jìn)行自然發(fā)病試驗(yàn),結(jié)果表明,轉(zhuǎn)基因株系的抗病表現(xiàn)優(yōu)于野生型玉米,幾乎沒有4級重度感染株,而野生型玉米全部發(fā)病,且4級重度感染株比例占37.5%,4個(gè)amiRNA載體中以靶向基因沉默抑制子的S6-miR159轉(zhuǎn)基因玉米抗病表現(xiàn)最好,健株(0級)和輕微感病株(1級)比例達(dá)41.5%,說明利用amiRNA技術(shù)培育抗粗縮病玉米新品種也可行。
2.2利用非病毒來源的抗病毒基因
盡管研究者采用不同策略轉(zhuǎn)化病毒基因序列均獲得了抗甚至高抗病毒病的玉米材料,但將病毒基因序列轉(zhuǎn)入植物,不確定因素較多,生物安全性一直以來備受人們質(zhì)疑,因此,研究人員多年來致力于發(fā)掘不同來源的抗病毒基因。
2.2.1寄主抗病基因 植物在與病原體長期相互作用過程中形成了抗病基因(Resistance gene,R基因)系統(tǒng),能夠?qū)Σ≡w的侵染產(chǎn)生防御反應(yīng)。至今在植物體內(nèi)克隆獲得具病毒抗性的R基因有40多種,多數(shù)基因的抗性具有特異性,人們利用植物R基因控制病毒已取得了很好的效果(牙庫甫江·阿西木等,2015)。關(guān)于玉米中矮花葉病和粗縮病抗性的數(shù)量性狀基因座(Quantitative trait locus,QTL)定位研究已有大量報(bào)道,但由于玉米對病毒抗性的遺傳基礎(chǔ)較復(fù)雜,從玉米中分離R基因的工作進(jìn)展緩慢。Shi等(2013)研究表明,ZmeIF4E參與調(diào)控防衛(wèi)基因表達(dá)并誘導(dǎo)寄主的局部和系統(tǒng)抗性,認(rèn)為其可能是玉米粗縮病抗性候選基因。還有研究發(fā)現(xiàn),玉米對SC-MV的抗性涉及Scmvl和Scmv2兩個(gè)主效QTLs,其分別位于第6和第3染色體上,在抗病早期和后期發(fā)揮作用(Liu et al.,2017)。瞿會(huì)(2016)利用元分析方法整合玉米Scmv2信息,在“一致性”QTL區(qū)段內(nèi)確定與抗病相關(guān)的候選基因,并克隆獲得完整的基因編碼區(qū)。Liu等(2017)針對Scmvl開展精細(xì)定位,克隆獲得玉米的主效抗病基因ZmTrxh,其編碼非典型的H型硫氧還蛋白,將ZmTrxh基因轉(zhuǎn)入HiⅡ中發(fā)現(xiàn),玉米早期對SCMV的抗性顯著提高,外源ZmTrxh的表達(dá)水平高低與植株的抗性呈高度正相關(guān)。
2.2.2核糖體失活蛋白基因 核糖體失活蛋白(Ribosome-inactivating proteins,RIPs)屬于N糖苷酶,能夠特異性地將28S rRNA位于4324A處的腺嘌呤糖苷鍵降解,阻斷EF2/GTP復(fù)合物與核糖體60S大亞基結(jié)合,使蛋白質(zhì)合成受抑,該蛋白能夠在病毒侵染細(xì)胞后使核糖體失活,導(dǎo)致病毒無法進(jìn)行復(fù)制(牙庫甫江·阿西木等,2015)。目前,玉米抗病毒基因工程研究中使用的RIPs主要是美洲商陸抗病毒蛋白(Pokeweed antiviral protein,PAP),其為從美洲商陸植株體內(nèi)提取的一種堿性毒蛋白,屬于Ⅰ型RIPs,具有對危害動(dòng)物、植物及人體的病原真菌、細(xì)菌和病毒的廣譜抗性。陳定虎等(2003)在轉(zhuǎn)PAP基因玉米幼苗葉片上接種SCMV,至25 d植株沒有發(fā)病,而對照植株在接種后7 d表現(xiàn)出花葉癥狀,表明PAP基因?qū)CMV侵染具有顯著抑制作用。由于PAP對植物病原具有廣譜抗性,其在植物保護(hù)領(lǐng)域具有廣闊的應(yīng)用前景。
2.2.3核酸酶基因 RNaseⅢ家族是一類依賴于dsRNA的特異性內(nèi)切核酸酶,在真核生物和原核生物中均有存在,植物病毒基因組多為ssRNA,其復(fù)制時(shí)形成dsRNA中間體,正好是RNaseⅢ家族的靶向底物(Zhang et al.,2001)。為了降低野生型RNaseⅢ對植物正常生長產(chǎn)生的危害,可通過突變使其只能結(jié)合而不能切割dsRNA。Cao等(2013)將來源于大腸桿菌的RNaseⅢ基因突變體rnc70轉(zhuǎn)化玉米綜31,然后將得到的轉(zhuǎn)基因植株種植于粗縮病重病田,經(jīng)多代篩選和自交,獲得對RBSDV具有高度抗性的2個(gè)轉(zhuǎn)基因玉米新株系。田間鑒定發(fā)現(xiàn),中等發(fā)病條件下,轉(zhuǎn)化植株發(fā)病率和發(fā)病程度極低,出現(xiàn)免疫植株;嚴(yán)重發(fā)生年份,部分植株(其中一個(gè)株系中有6/7的植株)也顯示出良好的抗性,而非轉(zhuǎn)基因植株高度感病。人工接種條件下,轉(zhuǎn)基因植株不僅發(fā)病率明顯降低,病株中病毒的含量也明顯下降。由于rnc70基因介導(dǎo)的抗性對能形成dsRNA結(jié)構(gòu)的病毒均有效,因此,轉(zhuǎn)rnc70基因的玉米材料可能對粗縮病、矮花葉病等多種玉米病毒病都表現(xiàn)出抗性,但需進(jìn)一步驗(yàn)證,以充分發(fā)揮其應(yīng)用潛力。
2.2.4其他抗病毒基因 dsRNA依賴性蛋白激酶R(PKR)具有多重功能,在寄主防御病毒侵染中發(fā)揮重要作用。PKR被細(xì)胞內(nèi)的dsRNA或病毒復(fù)制中間體激活后,能使真核生物翻譯起始因子elF-2的α亞單位磷酸化,導(dǎo)致病毒蛋白不能合成,從而抑制病毒繁殖(Sadler and Williams,2007)。PKR基因?qū)NA病毒具有廣譜抗性,其在玉米抗病毒基因工程中的功效值得探索。尹祥佳(2011)應(yīng)用In-Fusion克隆技術(shù)將人源PKR基因和抗草甘NEPSPS基因連接到載體上得到了雙價(jià)植物表達(dá)載體,轉(zhuǎn)化玉米自交系掖478和昌7-2后均獲得陽性轉(zhuǎn)化植株,為獲得兼具病毒病抗性和除草劑抗性的轉(zhuǎn)基因玉米材料打下了基礎(chǔ)。
2-5A體系主要存在于高等脊椎動(dòng)物體內(nèi),是一個(gè)由干擾素(1FN)誘導(dǎo)的降解單鏈RNA的抗病毒系統(tǒng),之后的研究發(fā)現(xiàn)其對dsRNA動(dòng)物病毒及植物病毒也表現(xiàn)出有效抗性,推測2-5A體系是具有應(yīng)用前景的廣譜抗病毒系統(tǒng)(Mitra et al.,1996)。張志燕(2004)將動(dòng)物2-5A體系的RNase L基因和2-5A基因同時(shí)轉(zhuǎn)入玉米自交系Z3和z31,均獲得雙陽性玉米再生植株,為培育具有廣譜病毒病抗性的玉米品系打下了基礎(chǔ)。
3展望
植物抗病毒基因工程研究所用的基因序列分為病毒來源的基因序列和非病毒來源的基因序列。由于病毒基因組小、序列易測定、基因易克隆和操作,利用病毒來源基因序列的抗病毒基因工程研究取得長足進(jìn)展,采用的相關(guān)策略可分為病毒編碼蛋白基因介導(dǎo)的抗病性、RNAi介導(dǎo)的抗病性和amiR-NA介導(dǎo)的抗病性3種。在玉米抗病毒基因工程研究中,利用前兩種策略均得到了不同抗病水平的轉(zhuǎn)基因植株,第3種策略的應(yīng)用剛起步,與前2種策略相比,具有精確、高效、可控且操作簡便的優(yōu)點(diǎn),在培育抗病毒植物中具有廣闊的應(yīng)用前景,因而有必要加強(qiáng)其在玉米抗病毒基因工程中的應(yīng)用。利用病毒基因序列通常只抗1種或與其相關(guān)的幾種病毒,目前提高矮花葉病抗性水平的研究相對較多,也獲得了大量抗性種質(zhì)材料,而抗粗縮病和兼抗2種病毒病的研究比較滯后,需要加強(qiáng)探索,以便盡快解決我國玉米生產(chǎn)上病毒病防治的難題。相對于病毒基因序列,將非病毒來源的抗性基因?qū)胗衩咨锇踩詴?huì)更高,但從玉米中分離抗病基因的工作進(jìn)展緩慢,還需要加快研究。此外,來自其他物種的核糖體失活蛋白、核酸酶、2-5A體系等的基因理論上均能促使受體植物獲得廣譜抗性,目前通過轉(zhuǎn)化這些基因也得到了較多的玉米材料,但其廣譜抗病毒效果有待進(jìn)一步驗(yàn)證。
玉米抗病毒基因工程的研究主要集中在采用不同策略獲得抗病毒植株及不同因素對抗病毒效果的影響方面,隨著技術(shù)的發(fā)展和抗病性的提高,人們開始關(guān)注其安全性問題。目前已有研究表明,轉(zhuǎn)基因抗矮花葉病玉米具有與常規(guī)玉米相同的生物學(xué)營養(yǎng)等價(jià)性,長期飼喂對大鼠的生長發(fā)育、生殖功能均無明顯影響,其對大鼠也不存在亞慢性毒性和遺傳毒性,但對大鼠的學(xué)習(xí)與記憶能力有明顯降低作用,推測轉(zhuǎn)基因玉米可能影響了神經(jīng)細(xì)胞的正常活性(周文麗等,2014)。由于相關(guān)研究較少,關(guān)于轉(zhuǎn)基因抗病毒玉米的食用安全性、生物安全性和生態(tài)安全性,還需進(jìn)行全面評價(jià)。轉(zhuǎn)基因玉米中關(guān)于抗蟲、抗除草劑玉米的研究較成熟并已進(jìn)入商品化生產(chǎn)階段,而抗病毒玉米的研究與發(fā)展遠(yuǎn)遠(yuǎn)落后,只有經(jīng)過抗病性和安全性的雙重驗(yàn)證,轉(zhuǎn)基因抗病毒玉米才能投入大田生產(chǎn),真正發(fā)揮其抗病作用。
