香蕉MaERF—1基因的克隆、序列分析及表達載體構建
2017-05-30 13:29:26宋順金志強徐碧玉黃東梅胡偉李凱許奕
熱帶作物學報 2017年1期
宋順 金志強 徐碧玉 黃東梅 胡偉 李凱 許奕

摘 要 從香蕉中克隆了1個乙烯響應因子(ERF)MaERF-1。序列分析表明,該基因存在1個完整的開放閱讀框(ORF)729 bp,編碼243個氨基酸。多序列比對和進化樹分析表明,MaERF-1所編碼的蛋白與其他植物中ERF編碼的蛋白具有較高的一致性。其中與馬來西亞野生香蕉同源性最高達98%,與油棕、菠蘿、海棗、葡萄、荷花、煙草的MaERF編碼的氨基酸序列的同源性分別為65%、60%、59%、54%、53%、51%。MaERF-1編碼的蛋白質分子量為26 139.03 u,理論等電點pI為7.81,其親水性氨基酸均勻分布在整個肽鏈中,多于疏水性氨基酸。通過PCR和酶切反應鑒定成功構建該基因的表達載體。
關鍵詞 乙烯響應因子;ERF;香蕉;生物信息學;表達載體
中圖分類號 S668.1 文獻標識碼 A
Gene Cloning, Sequence Analysis and Expression Vector
Construction of MaERF-1 from
Musa acuminate L. AAA
SONG Shun1, JIN Zhiqiang1, XU Biyu2, HUANG Dongmei1,
HU Wei2, LI Kai2, XU Yi1 *
1 Haikou Experimental Station, CATAS / Hainan Key Laboratory of Banana Genetic Improvement , Haikou, Hainan 570102, China
2 Institute of Tropical Bioscience and Biotechnology, CATAS / Key Laboratory of Biology and Genetic Resources of Tropical Crops,
Ministry of Agriculture, Haikou, Hainan 571101, China
Abstract We have isolated an ethylene response factor from banana, and designated as MaERF-1. The sequence analysis indicated that the complete ORF of MaERF-1 was 729 bp, and it encoded 243 amino acids. Alignment of amino acid sequences and phylogenetic analysis indicated that the protein encoded by MaERF-1 was in high similarity with ERF-encoding protein in other known plants and highly homologous with amino acid sequences in Musa acuminate by 98%, The similarity with Elaeis guineensis, Ananas comosus, Phoenix dactylifera, Nelumbo nucifera,Nicotiana tabacum was 65%, 60%, 59%, 54%, 53%, 51%, respectively. The protein molecular weight of MaERF-1 was 26 139.03 u, and the theoretical isoelectric point pI was 7.81. The hydrophilic amino acids were uniformly distributed throughout the peptide chain and more than hydrophobic amino acids. The expression vector of the gene was successfully constructed and identified by PCR and restriction enzyme digestion reactions. This experiment could be the basis for further study of MaERF-1 gene expression regulation and functional analysis.
Key words ethylene response factor; ERF; banana; bioinformatics; expression vector
doi 10.3969/j.issn.1000-2561.2017.01.014
植物會經常遭受到許多環境的非生物脅迫變化,如高鹽、干旱或者極端的溫度。現代農業的模式會導致鹽堿地的面積增加。預測到2050年,多余50%的土地均會遭受到鹽堿化[1]。干旱的發生在許多地方都很普遍并且會不斷擴大[2]。香蕉是世界水果貿易量及消費量最大宗的鮮果,被聯合國糧農組織(FAO)定位為發展中國家僅次于水稻、小麥、玉米之后的第四大糧食作物。然而香蕉前期植株較小,根系淺生,易受旱,其葉面積大,蒸騰量大,早在其發生萎蔫之前,就已表現出缺水現狀。尤其是在高溫季節,香蕉受到干旱后,葉片失水,相對含水量下降,水分脅迫導致香蕉葉片的細胞質膜透性增大,極大地傷害香蕉正常的生理代謝活動,造成香蕉減產,品質下降,嚴重影響植株的生長發育[3]。所以,提高香蕉抗旱能力對于香蕉生產至關重要。通過研究相關的功能基因來提高香蕉抵御干旱及非生物脅迫的能力是一個重要的途徑。
在植物中,轉錄因子在響應非生物脅迫中起著很重要的作用。ERF是一個轉錄因子家族,其在植物中能夠響應許多非生物脅迫[4-8]。ERF是一個大家族,在水稻、擬南芥和黃豆基因組中分別有157、147和148個成員[9-11]。ERF可以激活或抑制,在其啟動子具有GCC盒(AGCC GCC)基因的轉錄[12]。一些ERF能夠結合CRT/DRE(A/GCCGAC)結構從而調控基因響應生物或者非生物脅迫[13-16]。ERF參與許多生物過程,包括一些代謝過程,花和種子的發育,根系的發育[17-20]。例如,小麥TaERF1通過結合GCC盒和CRT/DRE元件,能夠激活一些與非生物脅迫相關的基因,如低溫脅迫、干旱脅迫和高鹽脅迫[21]。Mishra從罌粟中分離出了PsAP2基因,其能夠與DRE和GCC元件結合,將該基因轉化煙草,轉基因植株能夠增強對脅迫以及非生物脅迫的耐受力[22]。Gao等[23]從番茄中克隆了TERF1基因,并將其轉化水稻。在轉基因水稻植株中,其脯氨酸增加,減少了水分的丟失,提高對高鹽的干旱的耐受性。同時,TERF1誘導了LIP5、 Wcor4131、OsPrx和OsABA2的表達。但是,也有一些ERF基因會降低對脅迫的耐受性,如在擬南芥中過表達AtERF4降低了對ABA的敏感度和對Nacl的耐受力[24-25]。在OsERF922轉基因植株中顯示了,該植株減低了對高鹽脅迫的耐受性,其Na+/K+比高于野生型[26]。在StERF3的轉基因煙草中,其減低了對高鹽脅迫的耐受性,然而,在StERF3 RNAi抑制的植株中能夠提高對高鹽脅迫的耐受性,同時在該植株中,防御相關基因PR1、NPR1和 WRKY1也被誘導表達[27]。在土豆中沉默ERF3的表達能夠增強其對晚疫病的抵御能力[28]。同樣,在水稻中用RNAi沉默ERF922的表達能夠增強其對青枯病的抵御能力[29]。
本研究通過克隆香蕉水通道蛋白基因MaERF-1,利用生物信息學對該基因的功能進行進一步的分析,推測其生物學功能,同時成功構建了該基因的植物表達載體,為進一步研究ERF基因的表達調控以及功能分析奠定了基礎。
1 材料與方法
1.1 材料
1.1.1 植物材料 2016年1月初采集巴西香蕉(Musa acuminata L. AAA group‘Brazilian)的根、莖、葉、花、果,用無菌水清洗后立即放置于液氮中速凍,于 -70 ℃冰箱中保存備用。實驗所用香蕉均采自中國熱帶農業科學院熱帶生物技術研究所。
1.1.2 載體與菌株 克隆載體pMD18-T載體購自TaKaRa公司產品,大腸桿菌感受態DH5α購自天根公司。植物表達載體Pcambia 1304為實驗室保存。
1.1.3 酶與化學試劑 DNA Marker、限制性內切酶、LA Taq酶、LA PCR in vitro Cloning Kit購自TaKaRa公司。RevertAidTM First Strand cDNA Synthesis Kit購自Fermentas公司。質粒提取試劑盒、DNA回收試劑盒和RNA提取試劑盒購自Omega Bio-Tek公司,其他化學藥品為分析純。
1.2 方法
1.2.1 總RNA提取及cDNA獲得 取巴西蕉根、莖、葉、花、果組織,采用改良的CTAB法 / 植物RNA試劑盒(Omega)結合法提取和純化總RNA。以RevertAidTM First Strand cDNA Synthesis Kit 將RNA反轉錄成cDNA并置于-80 ℃冰箱中保存備用。
1.2.2 目的基因的獲得 從香蕉A基因組測序數據庫中得到一個ERF家族基因MaERF-1,根據其序列設計一對引物F1,R1(F1:5′-CATGCCATGG
CGATGGATTTTGAGGATTCCTCCTCCA-3′,R1:5′-GGACTAGTTCGGGTGGACGAACACCACATGG-3′,斜體堿基為酶切位點序列),擴增MaERF-1全長序列。PCR擴增程序為:95 ℃預變性3 min;94 ℃變性40 s,55 ℃退火40 s,72 ℃延伸50 s,共 35個循環。按照分子克隆實驗指南進行PCR 擴增產物回收、連接、轉化和鑒定。對已鑒定的陽性克隆進行測序分析。
1.2.3 生物信息學分析 將基因MaERF-1的cDNA序列和開放閱讀框(open reading frame,ORF)編碼的氨基酸序列在NCBI數據庫中的BLASTx進行同源性搜索和比對;利用Clustal X1.81和MEGA 3.1 軟件分析MaERF-1蛋白與其他植物的ERF蛋白的進化關系,構建分子進化樹;利用expasy數 據 庫(http://web.expasy.org/compute_pi/)預測蛋白質pI和分子質量;用Protscale(http://www.expasy.ch/tools/protscale.html/)工具對MaERF-1編碼的蛋白質產物疏水性和親水性進行分析。
1.2.4 MaERF-1表達載體的構建 選用Pcambia 1304植物表達載體構建MaERF-1表達基因融合載體,以NcoI和SpeI酶切pMD18-T/MaERF-1重組質粒和Pcambia 1304空載體,分別回收啟動子片段和線性載體,連接,轉化DH5α后進行菌液PCR及酶切檢測,重組質粒命名為MaERF-1::1304。
2 結果與分析
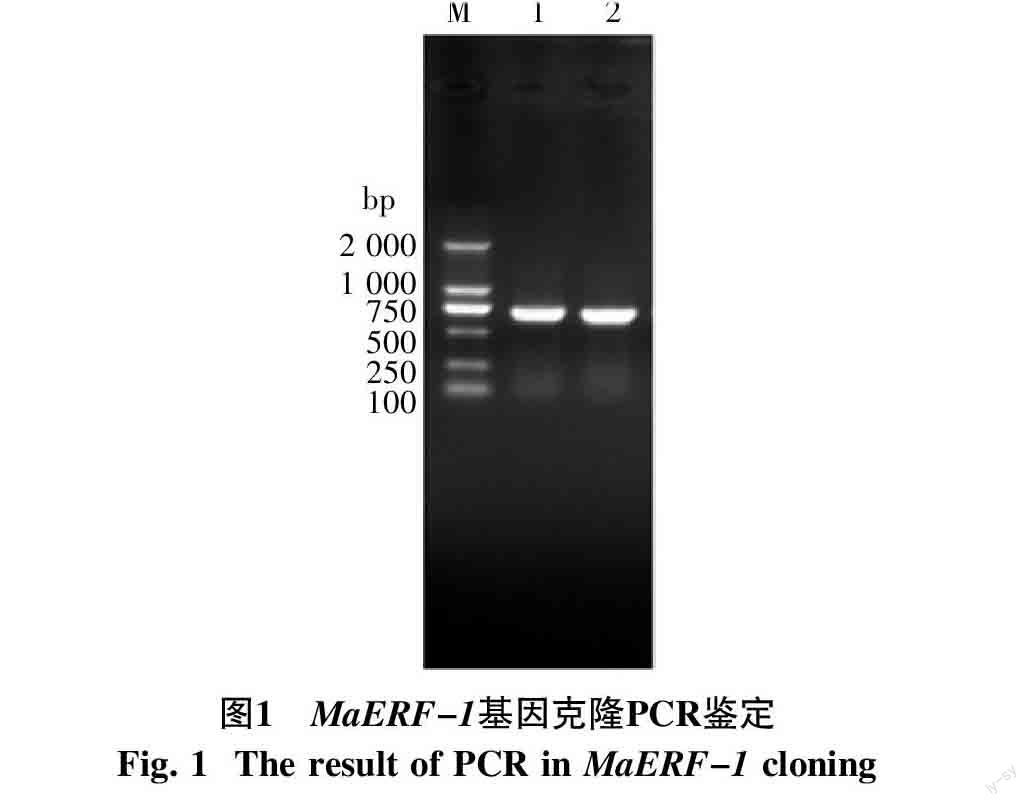
2.1 MaERF-1基因的克隆與鑒定
以巴西蕉各器官組織混合cDNA為模版,F1和R1為引物,從香蕉栽培品種巴西蕉中克隆到一個ERF家族基因MaERF-1,其ORF為729 bp,PCR結果如圖1所示。
2.2 生物信息學分析
2.2.1 MaERF-1基因蛋白質序列差異的比對 利用DNAman將MaERF-1 cDNA推導的氨基酸序列與NCBI中已登錄的其他高等植物的ERF氨基酸序列進行同源關系的比較,結果(圖2)顯示,各種植物ERF編碼的氨基酸序列存在較高的同源性,多數都達到55%以上。BLASTX分析表明,MaERF-1編碼的氨基酸序列與馬來西亞野生蕉MaERF-1-like(XP_009379780.1)、菠蘿AcERF-2(OAY63143.160)、海棗PcERF-2(XP_008807522.1)、荷花NnERF-2(XP_010243826.1)、葡萄VvERF-2(XP_002267961.1)、煙草NtERF-2(XP_016448076.1)、油棕EgERF-2(XP_010940400.1)編碼的氨基酸序列具有較高的一致性,分別為98%、60%、59%、53%、54%、51%、65%。
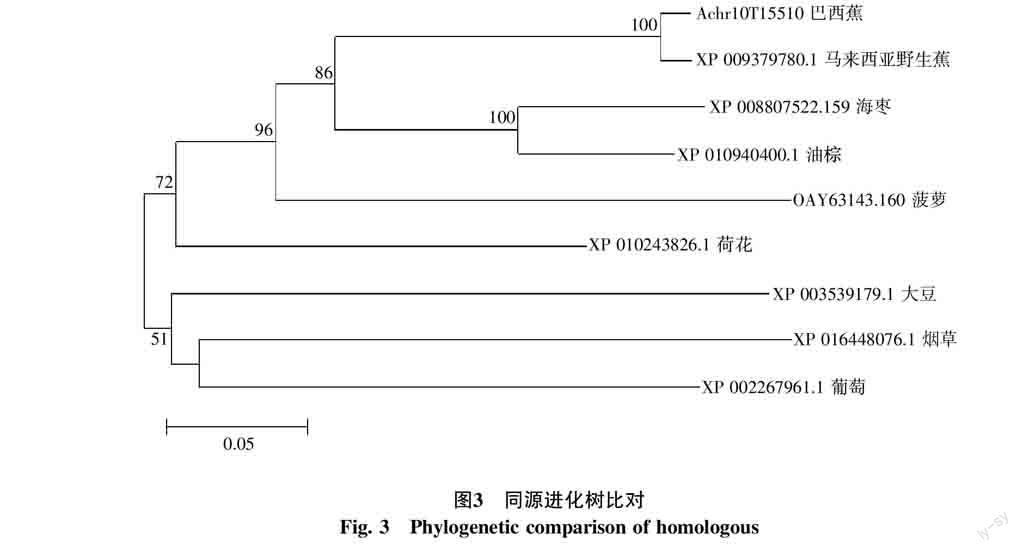
2.2.2 MaERF-1基因進化樹分析 利用Clustal X1.81和MEGA 3.1軟件,將MaERF-1 cDNA推導的氨基酸序列與NCBI中已登錄的其他植物的SIP氨基酸序列進行系統進化樹的比對分析。結果表明本研究得到的巴西蕉MaERF-1基因所編碼的氨基酸序列與馬來西亞野生蕉MaERF-1-like具有較近的親緣關系(圖3)。
2.2.3 MaERF-1編碼蛋白質理化性質分析 利用Expasy分析MaERF-1編碼蛋白質的理化性質,如圖4顯示,該蛋白具有243個氨基酸,分子量為26 139.03 u,理論等電點pI為7.81。
2.2.4 MaERF-1編碼蛋白質產物的親水性、疏水性分析 對MaERF-1編碼蛋白質產物親水性、疏水性分析結果表明(圖5),MaERF-1多肽鏈中精氨酸Arg具有最低的分值-2.800,異亮氨酸Ile具有最高分值1.600。依據氨基酸分值越低親水性越強和分值越高疏水性越強的規律,可見,精氨酸Arg親水性最強,而異亮氨酸Ile疏水性最強,而就整體來看,親水性氨基酸均勻分布在整個肽鏈中,多于疏水性氨基酸。
2.3 MaERF-1表達載體的構建
在上游引物F1和下游引物R1分別加入NcoI和SpeI內切酶,將pMD18-T/MaERF-1和Pcambia 1304載體同時用NcoI和SpeI內切酶酶切后,回收目的片段后連接轉化,PCR以及雙酶切鑒定如圖6,結果顯示,PCR產物和酶切條帶均為729 bp。
3 討論
香蕉是一年生草本植物,喜高溫多濕,缺水時葉片變薄、果指變短,生長延緩,導致減產,因此對香蕉抗逆品種的選育是十分必要的。近年來,通過挖掘香蕉本身所具有的抗逆基因資源,研究該基因的功能來進一步提高香蕉的抗逆性就成為一個重要的手段。
本實驗克隆到1個香蕉乙烯響應因子MaERF-1,通過在NCBI上BLAST分析各高等植物中ERF氨基酸序列的同源關系,結果顯示MaERF-1編碼的氨基酸與各種植物中ERF編碼的氨基酸序列存在較高的同源性,達到55%以上,其中與馬來西亞野生蕉同源性最高為98%。對各種植物ERF氨基酸序列比對和系統進化樹分析發現,不同植物的ERF序列相似性較高,以ERF氨基酸序列構建的進化樹能準確反映不同植物間的親緣關系,也可作為評價香蕉等植物種植資源的重要依據。通過對ERF編碼蛋白質的親水性和疏水性進行分析,表明其親水性氨基酸多于疏水性氨基酸。將MaERF-1成功構建到植物表達載體中,后續實驗可以通過將表達載體轉化到如擬南芥、煙草等模式植物中,對該基因進行進一步的功能驗證和分析。
當植物遭受到外界的非生物脅迫時,植物自身有復雜的系統去保護其去抵御不同的脅迫。ERF在植物中是屬于(AP2/ERF)轉錄因子家族中的亞族,對ERF的功能研究主要集中在對其在抗逆性方面的研究[30-32],如抗旱、抗鹽、抗寒等。例如,水稻ERF基因OsBIERF1、OsBIERF3 和OsBIERF4不僅能被稻瘟病菌誘導表達,還能受到高鹽、冷害、干旱、傷害脅迫而上調表達[33]。在煙草中過表達ERF11能夠提高植物抵御青枯病的能力。在擬南芥中,AtERF71能夠顯著被高鹽、甘露醇和ABA誘導表達,在高鹽脅迫下,植株中的活性氧(ROS)累積顯著減少[34]。轉化GmERF7的煙草能夠提高對高鹽脅迫的耐受性,該轉基因植株中葉綠素和糖含量顯著增加[35]。因此,本實驗通過對該基因的功能進行生物信息學分析,并且將其連接到植物表達載體中,該實驗結果為進一步研究ERF基因的表達調控以及后續轉基因分析基因功能奠定了基礎。
參考文獻
[1] Dean R, Van Kan J A, Pretorius Z A, et al. The Top 10 fungal pathogens in molecular plant pathology[J]. Mol Plant Pathol, 2012, 13(4): 414-430.
[2] Liu W, Liu J, Triplett L, et al. Novel insights into rice innate immunity against bacterial and fungal pathogens[J]. Annu Rev Phytopathol, 2014, 52: 213-241.
[3] Boyd L A, Ridout C, OSullivan D M, et al. Plant-pathogen interactions: disease resistance in modern agriculture[J]. Trends Genet, 2013, 29(4): 233-240.
[4] Chisholm S T, Coaker G, Day B, et al. Host-microbe interactions: shaping the evolution of the plant immune response[J]. Cell, 2006, 124(4): 803-814.
[5] Rojo E, Solano R, Sánchez-Serrano J J. Interactions between signaling compounds involved in plant defense[J]. J Plant Growth Regul, 2003, 22(1): 82-98.
[6] Glazebrook J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens[J]. Annu Rev Phytopathol, 2005, 43: 205-227.
[7] De Vos M, Van Oosten V R, Van Poecke R M, et al. Signal signature and transcriptome changes of Arabidopsis during pathogen and insect attack[J]. Mol Plant-Microbe Interact, 2005, 18(9): 923-937.
[8] Jung J, Won S Y, Suh S C, et al. The barley ERF-type transcription factor HvRAF confers enhanced pathogen resistance and salt tolerance in Arabidopsis[J]. Planta, 2007, 225(3): 575-588.
[9] Vogel M O, Moore M, Konig K, et al. Fast retrograde signaling in response to high light involves metabolite export, MITOGEN-ACTIVATED PROTEIN KINASE6, and AP2/ERF transcription factors in Arabidopsis[J]. Plant Cell, 2014, 26(3): 1 151-1 165.
[10] Muller M, Munne-Bosch S. Ethylene response factors: a key regulatory hub in hormone and stress signaling[J]. Plant Physiol, 2015, 169(1): 32-41.
[11] Cao Y, Song F, Goodman R M, et al. Molecular characterization of four rice genes encoding ethylene-responsive transcriptional factors and their expressions in response to biotic and abiotic stress[J]. J Plant Physiol, 2006, 163(11): 1 167-1 178.
[12] Lai Y, Dang F, Lin J, et al. Overexpression of a Chinese cabbage BrERF11 transcription factor enhances disease resistance to Ralstonia solanacearum in tobacco[J]. Plant Physiol Biochem, 2013, 62: 70-78.
[13] Sherif S, El-Sharkawy I, Paliyath G, et al. PpERF3b, a transcriptional repressor from peach, contributes to disease susceptibility and side branching in EAR-dependent and -independent fashions[J]. Plant Cell Rep, 2013, 32(7): 1 111-1 124.
[14] Tian Z, He Q, Wang H, et al. The potato ERF transcription factor StERF3 negatively regulates resistance to Phytophthora infestans and salt tolerance in potato[J]. Plant Cell Physiol, 2015, 56(5): 992-1 005.
[15] Liu D F, Chen X J, Liu J Q, et al. The rice ERF transcription factor OsERF922 negatively regulates resistance to Magnaporthe oryzae and salt tolerance[J]. J Exp Bot, 2012, 63(10): 3 899-3 912.
[16] Jia Y. Marker assisted selection for the control of rice blast disease[J]. Pestic Outlook, 2003, 14(4): 150-152.
[17] Gutterson N, Reuber T L. Regulation of disease resistance pathways by AP2/ERF transcription factors[J]. Curr Opin Plant Biol, 2004, 7: 465-471.
[18] Nakano T, Suzuki K, Fujimura T, et al. Genome-wide analysis of the ERF gene family in Arabidopsis and rice[J]. Plant Physiol, 2006, 140: 411-432.
[19] Fujimoto S Y, Ohta M, Usui A, et al. Arabidopsis ethylene-responsive element binding factors act as transcriptional activators or repressors of GCC box-mediated gene expression[J]. Plant Cell, 2000, 12: 393-404.
[20] Büttner M, Singh K B. Arabidopsis thaliana ethylene-responsive element binding protein (AtEBP), an ethylene-inducible, GCC box DNA-binding protein interacts with an ocs element binding protein[J]. Proc Natl Acad Sci, 1997, 94: 5 961-5 966.
[21] Zhou J, Tang X, Martin G B. The Pto kinase conferring resistance to tomato bacterial speck disease interacts with proteins that bind a cis-element of pathogenesis-related genes[J]. EMBO J, 1997, 16: 3 207-3 218.
[22] Solano R, Stepanova A, Chao Q, et al. Nuclear events in ethylene signaling: a transcriptional cascade mediated by ETHYLENE-INSENSITIVE3 and ETHYLENE-RESPONSE-FACTOR1[J]. Genes Dev, 1998, 12: 3 703-3 714.
[23] Berrocal-Lobo M, Molina A, Solano R. Constitutive expression of ETHYLENE-RESPONSE-FACTOR1 in Arabidopsis confers resistance to several necrotrophic fungi[J]. Plant J, 2002, 29: 23-32.
[24] McGrath K C, Dombrecht B, Manners J M, et al. Repressor- and activator-type ethylene response factors functioning in jasmonate signaling and disease resistance identified via a genome-wide screen of Arabidopsis transcription factor gene expression[J]. Plant Physiol, 2005, 139: 949-959.
[25] Onate-Sánchez L, Anderson J P, Young J, et al. AtERF14, a member of the ERF family of transcription factors, plays a nonredundant role in plant defense[J]. Plant Physiol, 2007, 143: 400-409.
[26] Pré M, Atallah M, Champion A, et al. The AP2/ERF domain transcription factor ORA59 integrates jasmonic acid and ethylene signals in plant defense[J]. Plant Physiol, 2008, 147: 1 347-1 357.
[27] Moffat C S, Ingle R A, Wathugala D L, et al. ERF5 and ERF6 play redundant roles as positive regulators of JA/Et-mediated defense against Botrytis cinerea in Arabidopsis[J]. PLos One, 2012, 7(4): e35995.
[28] Huang P Y, Catinot J, Zimmerli L. Ethylene response factors in Arabidopsis immunity[J]. J Exp Bot, 2016, 67(5): 1 231-1 241.
[29] Sakuma Y, Liu Q, Dubouzet J G, et al. DNA-binding specificity of the ERF/AP2 domain of Arabidopsis DREBs, transcription factors involved in dehydration- and cold-inducible gene expression[J]. Biochem Biophys Res Commun, 2002, 290: 998-1 009.
[30] Pieterse C M J, Leon-Reyes A, Van der Ent S, et al. Networking by small-molecule hormones in plant immunity[J]. Nat Chem Biol, 2009, 5: 308-316.
[31] Windram O, Madhou P, McHattie S, et al. Arabidopsis defense against Botrytis cinerea: chronology and regulation deciphered by high-resolution temporal transcriptomic analysis[J]. Plant Cell, 2012, 24: 3 530-3 557.
[32] Lewis L A, Polanski K, Torres-Zabala M de, et al. Transcriptional dynamics driving MAMP-triggered immunity and pathogen effector-mediated immunosuppression in Arabidopsis leaves following infection with Pseudomonas syringae pv. tomato DC3000[J]. Plant Cell, 2015, 27: 3 038-3 064.
[33] Buscaill P, Rivas S. Transcriptional control of plant defence responses[J]. Curr Opin Plant Biol, 2014, 20: 35-46.
[34] Lorenzo O, Piqueras R, Sánchez-Serrano J J, et al. ETHYLENE RESPONSE FACTOR1 integrates signals from ethylene and jasmonate pathways in plant defense[J]. Plant Cell, 2003, 15: 165-178.
[35] Brown R L, Kazan K, McGrath K, et al. A role for the GCC-box in jasmonate-mediated activation of the PDF1.2 gene of Arabidopsis[J]. Plant Physiol, 2003, 132: 1 020-1 032.
