利用代謝組學方法分析生防菌抑菌機理模式的初步建立
2017-05-30 17:22:15李艷波史懷
熱帶作物學報 2017年1期
李艷波 史懷
摘 要 應用9種殺菌劑來處理尖孢鐮刀菌,通過HPLC/Q-TOF方法進行代謝輪廓分析。結果表明:不同菌劑處理后的尖孢鐮刀菌代謝組顯著差異,PCA分析可將其分為6組。將6組殺菌劑作為訓練樣本集,建立殺菌劑對尖孢鐮刀菌抑菌機理的PLS-DA模型。以枯萎病生防菌短短芽胞桿菌為測試樣本,利用該模型對其活性物質進行了抑菌機理的分析判別,發現其作用方式與嘧菌酯、氰霜唑類似,即通過抑制病原菌的呼吸作用而發揮抑菌功能,說明利用該模型判別生防菌對尖孢鐮刀菌的抑菌機理是可行的。
關鍵詞 代謝組學;殺菌劑;PLS-DA模型
中圖分類號 Q946 文獻標識碼 A
Establishment of a Analysis Method for Antifungal Modes of
Biocontrol Strains on Cucumber Fusarium
Wilt by Metabonomics
LI Yanbo1, SHI Huai2 *
1 Department of Biotechnology, Fujian Vocational College of Agriculture, Fuzhou, Fujian 350119, China
2 Agricultural Bioresources Research Institute, Fujian Academy of Agricultural Sciences, Fuzhou, Fujian 350003, China
Abstract In this investigation, the antifungal modes of action of nine fungicide with different targets or modes-of-action on Fusarium oxysporum were studied. Metabolic profiles of cultures treated with nine different fungicide were acquired by LC/Q-TOF MS, significant metabolic markers were used for classification by PCA analysis, and the fungicide could be divided into six groups. Using the six groups as training sample set, PLS-DA prediction model were established. Confirmed with the model, the conclusion could be drawn that the antifungal mode of action of Brevibacillus brevis, a biocontrol strain of Cucumber Fusarium Wilt, is similar to that of azoxystrobin and Cyazofamid, which act as respiratory inhibitor.
Key Words metabonomic; fungicide; PLS-DA prediction model
doi 10.3969/j.issn.1000-2561.2017.01.027
利用環境中一些對植物病原菌具有拮抗、抑制、競爭、寄生及誘導植物抗性等作用的有益微生物及其代謝產物進行植物病害防治,是現代植物病害控制的一個重要手段[1]。然而,盡管中國在植物病害生物防治方面取得了一批研究成果,但仍存在諸多問題,尤其是許多生防菌功能物質及其作用機理尚不明確[2]。這對提高生防菌抑菌活性,充分發揮其作用,以及基于作用靶標的新藥物分子生物合成設計研究造成了嚴重制約。
代謝組學(metabonomics)作為系統生物學的組成部分,顯示的是生物系統對基因以及環境變化的最終應答,代表著“組學”層次上的結束[3]。近年來,代謝組學研究取得了長足發展,在生物、醫藥等領域的研究中發揮了重要的作用[4-10]。利用代謝組學研究外源物質引起的病原菌內源性代謝物變化,能更直接地了解病原菌體內生物化學過程和狀態變化,從而更高效地探索外源物質作用靶標及作用機制。如Yu等[11]利用LC-MS比較了金果欖(Tinospora capillipes)提取物和已知作用機理的9種天然抗生素處理后金色釀膿葡萄球菌(Staphylococcus aureus)代謝輪廓變化,明確了提取物的作用機理類似于利福霉素(rifampicin)和諾氟沙星(norfloxacin)。易智彪[12]以金黃色葡萄球菌為靶標,建立已知抗生素的抗菌模式圖,進行了4種抑菌中草藥活性成分的代謝組學研究,確認桔皮黃酮提取物的抑菌機理是完全有別于現有已知機理的抗生素。這些藥物作用機理的代謝組學研究主要集中于醫藥方面,而在農藥,尤其是生物源天然活性抑菌物質的作用機理上,目前尚未見報道。
本研究采用基于HPLC/Q-TOF MS的代謝組學分析方法,以黃瓜枯萎病病原菌尖孢鐮刀菌(Fusarium oxysporum Schl.)為研究對象,通過分析已知殺菌劑對尖孢鐮刀菌代謝組的影響,建立PLS-DA預測模型并進行驗證,為進一步研究相關抑菌物質的作用機理提供參考。
1 材料與方法
1.1 材料
1.1.1 儀器與試劑 Agilent 1260/6520 Q-TOF LC/MS,配ESI電噴霧離子源;Eppendorf高速冷凍離心機;Sanyo超低溫冰箱;甲醇、乙腈、乙酸銨,均為色譜純,生產廠家為默克。供試殺菌劑原藥均購自百靈威科技有限公司。
1.1.2 供試菌株 尖孢鐮刀菌FJAT-9728菌株,由福建省農業科學院農業生物資源研究所提供。
1.2 方法
1.2.1 供試殺菌劑對尖孢鐮刀菌的室內毒力測定
菌株活化后接種于察氏培養基(蔗糖3 g/L,NaNO3 0.3 g/L,K2HPO4 0.1 g/L,KCl 0.05 g/L,MgSO4·7H2O 0.05 g/L,FeSO4 0.001 g/L,瓊脂2 g)上,25 ℃培養3 d,用直徑5 mm的打孔器切取菌落外緣制備成菌餅。將9種殺菌劑分別制成含0.1、1、10、100、200 μg/mL的察氏平板,以不加殺菌劑的平板為對照,每個濃度重復3次。將菌餅置于含藥培養基中央,25 ℃培養3 d后測定菌落直徑,以平均數代表菌落大小,計算各殺菌劑的相對抑制率、毒力回歸方程、EC50值。
1.2.2 菌株培養 菌株于察氏培養基25 ℃培養7 d,用0.03%吐溫80沖洗菌落,孢子懸液經滅菌的脫脂棉過濾后計數并稀釋至1×106 spores/mL,取200 μL均勻涂布在分別內含各供試殺菌劑以及枯萎病生防菌短短芽胞桿菌(Brevibacillus brevis)活性代謝產物并鋪有玻璃紙的察氏培養基上,每個處理3次重復,另設空白對照,25 ℃培養7 d。殺菌劑的使用濃度根據測得的EC50值設定,短短芽胞桿菌活性代謝產物的提取參照黃素芳等[13]的方法。
1.2.3 樣品制備 參照羅飛飛等[14]的方法并稍加改進:菌株培養7 d后將菌落刮入50 mL離心管,加入20 mL 0.03%吐溫80渦旋3 min,經20 μm無菌濾布過濾后蒸餾水沖洗,菌絲裝入5 mL離心管,液氮冷凍淬滅,凍干48 h后將菌絲粉碎。各處理分別稱取10 mg凍干的菌絲,加入2 mL 80% 甲醇渦旋30 s,超聲提取1 h后4 ℃靜置12 h。提取物5 600 ×g離心10 min,取1.5 mL上清,0.22 μm濾膜過濾后進行HPLC-MS分析。
1.2.4 液相色譜與質譜條件 色譜條件:Agilent ZORBAX Extend-C18色譜柱(2.1×50 mm,1.8-Micron);進樣體積10 μL;柱溫35 ℃;流動相A=10 mmol/L乙酸銨溶液,流動相B=乙腈,梯度洗脫程序:0~8 min 10% A,8~15 min 10%~50% A,15~30 min 50% A;30~40 min 50%~10% A,40~55 min 10% A。
質譜條件:ESI源;正離子模式;毛細管電壓20 V;噴霧器壓力30 psi;干燥氣流量10 L/min,干燥氣溫度300 ℃;碰撞電壓230 V;椎體孔電壓65 V;質量范圍100~1 000 m/z;標準質量121.050 873、922.009 798[15]。
1.3 數據分析
參照史懷等[6]的方法,原始LC-MS數據利用Agilent MassHunter工作站軟件(含Qualitative Anlysis和Mass Profiler Professional模塊)進行處理,包括分子特征提取(MFE)、背景扣除、數據篩選。9個殺菌劑及空白共10個處理3次重復30組數據經處理后,以檢測到的代謝物及其相對豐度值為指標,以各處理為樣本,構建數據矩陣進行主成分(PCA)分析,選取對代謝輪廓變化影響最大的2個PC值作得分散點圖;以不同殺菌劑處理樣品為訓練樣本集,通過PLS-DA算法建立預測模型,并將生防菌活性代謝物處理樣品作為未知樣品導入模型預測其殺菌機理。
2 結果與分析
2.1 供試殺菌劑對尖孢鐮刀菌的室內毒力測定
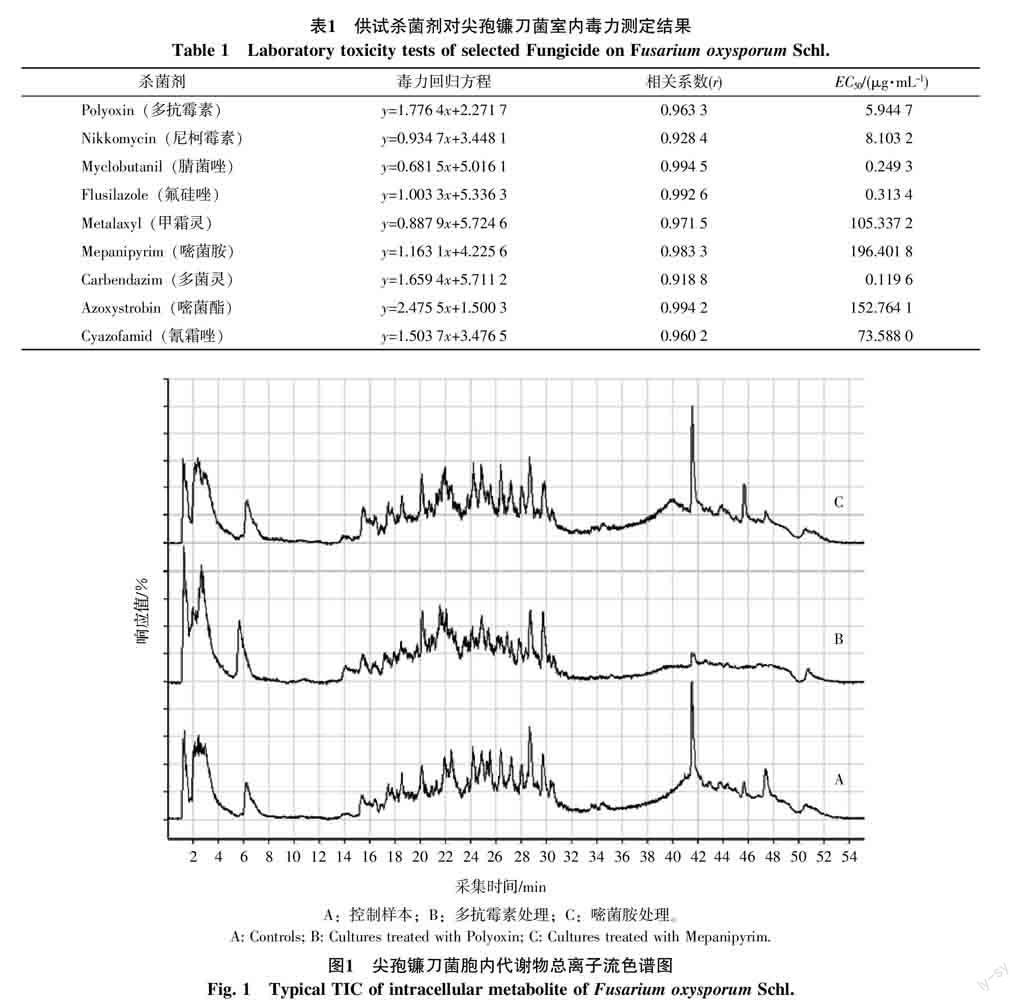
室內毒力測定的結果表明,供試的9種殺菌劑對尖孢鐮刀菌均有抑制作用,其毒力回歸方程及EC50值見表1。
根據測定的結果,試驗殺菌劑的使用濃度設置為:多抗霉素6 μg/mL、尼柯霉素8 μg/mL、腈菌唑0.25 μg/mL、氟硅唑0.3 μg/mL、甲霜靈100 μg/mL、嘧菌胺200 μg/mL、多菌靈0.12 μg/mL、嘧菌酯150 μg/mL、氰霜唑75 μg/mL。
2.2 不同殺菌劑處理后的尖孢鐮刀菌代謝物
圖1為部分尖孢鐮刀菌樣品的總離子流色譜圖,其中A、B、C分別為控制樣本、多抗霉素處理以及嘧菌胺處理。由圖1可知,控制樣本與殺菌劑處理后得到的代謝物譜圖輪廓相似,其代謝物的出峰時間主要集中在0~8、14~32、38~52 min 3個時間段。每個處理的代謝物集中出現區域波譜形態與強度并不完全一致,尤其在38~52 min區間有明顯差異。
2.3 不同致病性尖孢鐮刀菌代謝組學PCA分析
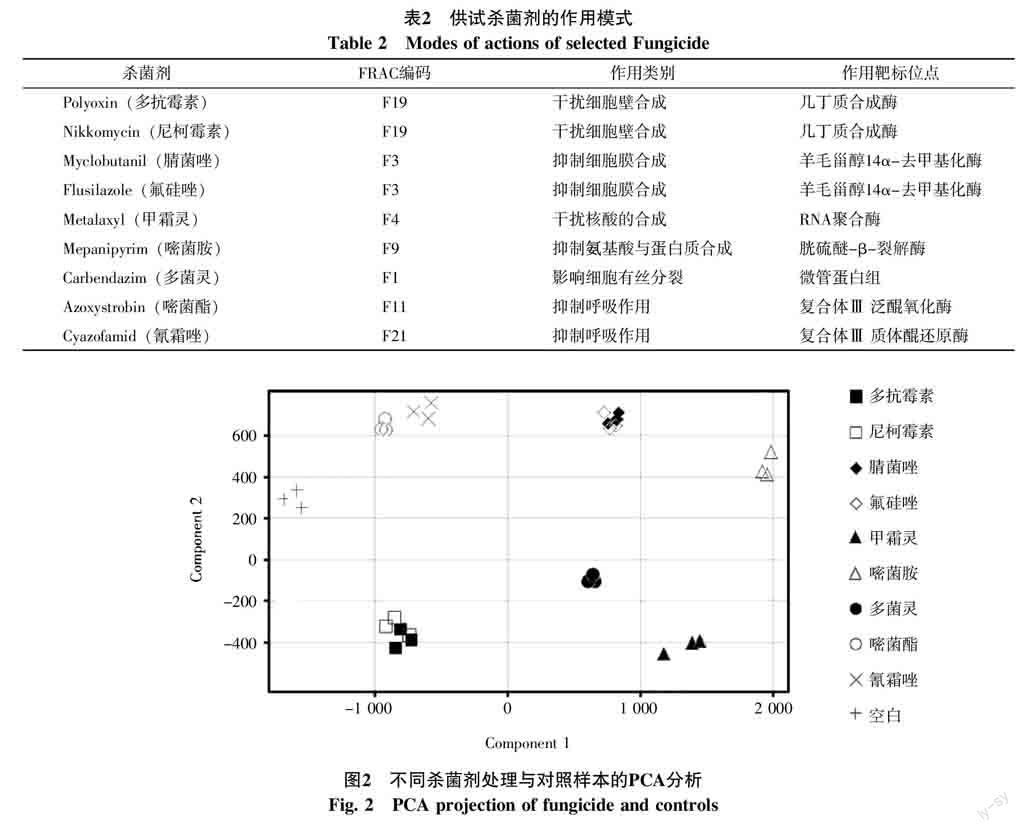
由圖2可知,各殺菌劑處理后的樣本均能與對照樣本清晰的區分開來,而具有相同作用機理的殺菌劑其投影聚集在一起(殺菌劑的作用模式見表2[16])。多菌靈(●)的作用靶標是微管蛋白組,影響真菌的有絲分裂,其集群明顯與其他作用方式的殺菌劑分離開來,甲霜靈(▲)和嘧菌胺(△)也是如此。而多抗霉素(■)和尼柯霉素(□)都是作用于幾丁質合成酶,干擾病原菌的細胞壁生成,被聚為一組,同樣,具有相同作用靶標的腈菌唑(◆)與氟硅唑(◇)也被聚集在同一組。相對于其他殺菌劑,嘧菌酯(○)和氰霜唑(×)對抑制呼吸作用位點不同,但其作用機理相似,所以腈菌唑(◆)與氟硅唑(◇)在圖2中十分接近。說明尖胞鐮刀菌受到殺菌劑影響后,在統計學意義上,相同或相似作用位點的殺菌劑具有相同的代謝信息,而不同作用模式的殺菌劑的代謝信息完全不同。
2.4 殺菌劑作用機理模型建立與生防菌作用機理分析
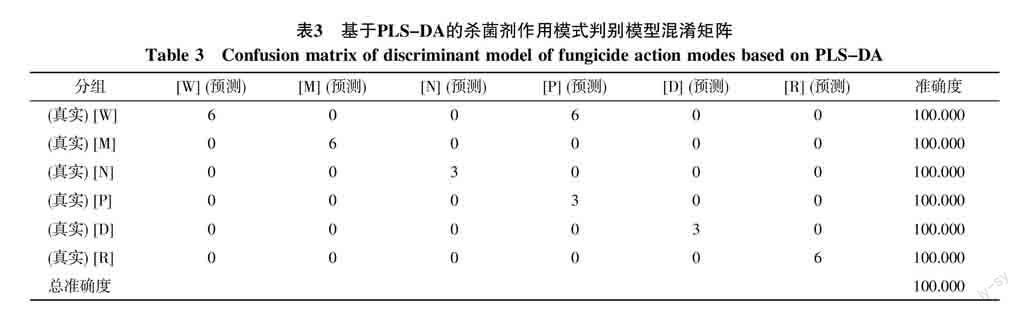
通過PCA分析得知不同作用模式的殺菌劑處理后尖胞鐮刀菌的代謝行為不同,為了提高代謝組學方法鑒定未知物質抑菌機理的能力,本研究進一步利用供試殺菌劑處理后的尖胞鐮刀菌代謝物構建統計學預測模型,利用該模型對未知物質進行殺菌機理的預測。將不同處理的樣本按殺菌機理分為影響細胞壁合成(W)、影響細胞膜合成(M)、影響核酸合成(N)、影響氨基酸與蛋白合成(P)、干擾有絲分裂(D)以及抑制呼吸作用(R)6個組,作為訓練樣本集,建立基于通過偏最小二乘判別分析(PLS-DA)算法的預測模型。從表3的混淆矩陣中可知,訓練樣本集的各樣品預測準確度均達到100%,表明該方法具有極高的預測能力。
以枯萎病生防菌短短芽胞桿菌(Brevibacillus brevis)的活性代謝產物作為未知樣本,導入模型中進行預測。3個重復樣品的預測結果均被歸類于R組,即抑制呼吸作用,其置信測度分別為0.903、0.896和0.915(表4)。
3 討論
用9種已知抑菌機理的殺菌劑分別處理尖孢鐮刀菌,采用HPLC/Q-TOF方法對尖孢鐮刀菌代謝輪廓進行分析結果表明,殺菌劑處理以及空白對照組的尖孢鐮刀菌代謝輪廓有顯著差異,在PCA得分圖中可明顯區分。把不同殺菌劑處理的樣品分為6種抑菌模式作為訓練樣本集,建立殺菌劑作用方式的PLS-DA模型,并利用該模型對枯萎病生防菌短短芽胞桿菌的活性物質進行了抑菌機理的預判結果表明,其作用方式與嘧菌酯、氰霜唑類似,即通過抑制病原菌的呼吸作用而發揮抑菌功能。
短短芽胞桿菌FJAT-0809-GXL菌株是福建省農業科學院農業微生物創新團隊篩選到的一株高效生防菌,該菌株對作物枯萎病的病原菌尖孢鐮刀菌具有很好的抑制作用[17-18]。車建美等[19-20]對其抑菌功能成分進行了分析,認為活性成分是多種物質的混合物,但對尖孢鐮刀菌起生防作用的主要成分為羥苯乙酯。羥苯乙酯屬于尼泊金酯類防腐劑,能夠破壞生物氧化過程中電子傳遞鏈的完整性,抑制病原菌的呼吸作用,從而抑制病原菌生長[21]。這與本研究通過建立殺菌劑作用機理判別模型預測的結果相符,證明該模型預測具有可行性。
通常,生防菌活性物質的發現、確定以及其作用機理的研究需要進行大量的試驗,采用代謝組學的方法可以快速的對活性物質作用方式進行預判,避免了繁瑣的實驗步驟,為生防菌的活性物質發現及其作用機理的研究提供了一條新途徑。當然,本研究選取的殺菌劑并不能完全代表所有的已知殺菌作用方式,因此建立的模型尚不完善,有待后續實驗繼續補充。
參考文獻
[1] Cook R J. Making greater use of introduced microorganisms for biological control of palt pathogens[J]. Annual Review of Phytopathology, 1993, 31: 53-80.
[2] 邱德文. 我國植物病害生物防治的現狀及發展策略[J]. 植物保護, 2010(4): 15-18, 35.
[3] Vaclavik L, Lacina O, Hajslova J, et al. The use of high performance liquid chromatography-quadrupole time-of-flight mass spectrometry coupled to advanced data mining and chemometric tools for discrimination and classification of red wines according to their variety[J]. Analytica Chimica Acta, 2011, 685: 45-51.
[4] German J B, Roberts M A, Watldns S M. Personal metabolomics as a next generation nutritional assessment[J]. Journal of Nutrition, 2003, 133(12): 4 260-4 266.
[5] Widodo, Patterson J H, Newbigin E, et al. Metabolic responses to salt stress of barley (Hordeum vulgare L.) cultivars, Sahara and Clipper, which differ in salinity tolerance[J]. Journal of Experimental Botany, 2009, 60: 4 089-4 103.
[6] 史 懷, 劉 波, 陳 崢, 等. 基于LC/Q-TOF MS的芽孢桿菌代謝組學分析方法[J]. 福建農業學報, 2012, 27(10): 1 112-1 119.
[7] Buchholz A, Hurlebaus J, Wandery C, et al. Metabolomics: quantification of intracellular metabolite dynamics[J]. Biomol Eng, 2002, 19(1): 5-15
[8] Boersma M G, Solyanikova I P, VanBerkel W J H, et al. 19 NMR metabolomics for the elucidation of microbial degradation pathways of fluorophenols[J]. J Ind Microbiol Biotechnol, 2001, 26(1/2): 22-34.
[9] Brindle J T, Antti H, Holmes E, et al. Rapid and noninvasive diagnosis of the presence and severity of coronary heart disease using 1H NMR-based metabonomics[J]. Nature Medicine, 2002, 8(12): 1 439-1 442.
[10] Lindon J C, Nicholson J K, Holmes E, et al. Contemporary issues in toxicology the role of metabonomics in toxicology and its evaluation by the COMET project[J]. Toxieol Appl Pharmacol, 2003, 187(3): 137-146.
[11] Yu Y, Yi Z B, Liang Y Z, et al. Main antimicrobial components of Tinospora capiilipes, and their mode of action against Staphylococcus aureus[J]. FEBS Letters, 2007, 581: 4 179-4 183.
[12] 易智彪. 四種抑菌中草藥活性成分的代謝組學研究[D]. 長沙: 中南大學, 2007.
[13] 黃素芳, 車建美, 劉 波, 等. 短短芽胞桿菌FJAT-0809-GXL代謝產物活性物質提取方法的優化[J]. 福建農業學報, 2011, 26(4): 528-532.
[14] 羅飛飛, 李淑林, 陳龍云, 等. 代謝組學方法鑒定球孢白僵菌孢子萌發和殺蟲毒力相關的標記物[J]. 微生物學報, 2014, 54(1): 33-41.
[15] 史 懷, 劉 波, 陳 崢, 等. 青枯雷爾氏菌致病性的代謝組學異質性研究[J]. 福建農業學報, 2013, 28(4): 343-352.
[16] FRAC Code List 2015: Fungicides sorted by mode of action (including FRAC Code numbering)[EB/OL]. [2016-06-25]. http: //www.frac.info/publications/downloads.
[17] 葛慈斌, 劉 波, 藍江林, 等. 生肪菌JK-2對尖孢鐮刀菌抑制特性的研究[J]. 福建農業學報, 2009, 24(1): 29-34.
[18] 黃素芳, 肖榮風, 楊述省, 等. 短短芽孢桿菌JK-2胞外物質抗香蕉枯菌病菌的穩定性[J]. 中國農學通報, 2010, 26(18): 284-288.
[19] 車建美. 短短芽胞桿菌 (Brevibacillus brevis) 對龍眼保鮮機理的研究[D]. 福州: 福建農林大學, 2011.
[20] Che J M, Liu B, Chen Z, et al. Identification of ethylparaben as the antimicrobial substance produced by Brevibacillus brevis FJAT-0809-GLX[J]. Microbiological Research, 2015, 172: 48-56.
[21] 袁海濤, 芮漢明, 寧正祥. 尼泊金酯類和富馬酸酯類抑菌效果的研究[J]. 食品科技, 2001(5): 43-44.
