Fkbp51基因敲除對小鼠肝臟轉錄組基因可變剪接的影響
2017-06-09 08:58:47周志強楊志偉雍偉東
中國比較醫學雜志 2017年5期
周志強,楊志偉,雍偉東
(中國醫學科學院醫學實驗動物研究所,北京協和醫學院比較醫學中心,衛生部人類疾病比較醫學重點實驗室,國家中醫藥管理局人類疾病動物模型三級實驗室,北京 100021)
研究報告
Fkbp51基因敲除對小鼠肝臟轉錄組基因可變剪接的影響
周志強,楊志偉*,雍偉東*
(中國醫學科學院醫學實驗動物研究所,北京協和醫學院比較醫學中心,衛生部人類疾病比較醫學重點實驗室,國家中醫藥管理局人類疾病動物模型三級實驗室,北京 100021)
目的 通過分析Fkbp51基因敲除(knock out,KO)與野生型(wild type,WT)小鼠肝臟表達譜,研究Fkbp51基因敲除對肝臟組織基因可變剪接的影響。方法 利用二代測序對Fkbp51 KO與WT小鼠肝臟進行表達譜測序,用TopHat對RNA測序結果進行可變剪接分析,篩選出KO與WT小鼠肝組織中差異的內含子保留(intron retetion,RI)和外顯子跳躍(exon skipping,SE)。通過在線工具DAVID對這些差異可變剪接體進行基因功能 (gene ontology, GO)和代謝通路(kyoto encyclopedia of genes and genomes,KEGG)富集分析,同時用NCBI 基因數據庫對這些基因進行注釋。結果 (1)Fkbp51缺失可導致小鼠肝臟mRNA可變剪接發生變化;(2)Fkbp51基因敲除造成小鼠肝臟mRNA可變剪接表達量的變化;(3)通過GO與KEGG分析,我們發現這些發生差異可變剪切的基因主要與脂肪相關衍生物的代謝、免疫、膽汁酸分泌等通路相關。(4)與差異內含子保留相關的基因主要與肌動蛋白細胞骨架調控,氨基酸及其衍生物代謝相關。結論Fkbp51基因敲除能夠改變基因組中mRNA的可變剪切,進而影響小鼠肝臟的代謝功能。
Fkbp51基因敲除小鼠;肝臟; RNA-seq;可變剪接;內含子保留;外顯子跳躍
Fkbp51是一種大分子免疫親和蛋白,可介導蛋白與蛋白之間的相互作用。同時,Fkbp51作為共伴侶蛋白通過與熱休克蛋白HSP90/HSP70形成復合物參與激素受體復合物的形成與調控[1]。研究表明Fkbp51可通過調節糖、脂代謝來發揮重要的調控作用。肝臟是進行糖、脂代謝的主要器官。在肝臟中,糖皮質激素可以促進糖異生,但它的過度分泌會造成糖代謝綜合征,如向心性肥胖、糖尿病、胰島素抵抗和脂肪病變等。糖皮質激素結合皮質激素受體后,激活后續通路,促進Fkbp51的表達,進而負反饋作用于糖皮質激素受體,降低其對糖皮質激素的敏感性,以維持機體平衡[2-4]。近年來,Fkbp51在脂肪細胞分化中所起的重要作用引起了研究者越來越多的關注,Stechschulte[5]發現脂肪生成需要Fkbp51的參與;Toneatto J[6]發現Fkbp51可抑制3T3-L1前脂肪細胞的分化。我們的研究發現,Fkbp51 KO小鼠能抵制高脂飲食誘導的肥胖,說明Fkbp51基因在能量代謝和脂肪代謝中發揮著重要的作用[7]。
可變剪接是mRNA前體通過不同的剪接方式產生的剪接異構體,可變剪接可以調節基因表達,同時也是產生蛋白質多樣性的原因之一。可變剪接參與了多種重要的發育過程,如果蠅性別決定[8]、人類器官和組織發育[9, 10]、肌肉組織分化[11]、CD分子的差異表達[12]、mRNA產物的降解等[13]。異常的剪接會導致疾病的發生,如地中海貧血癥[14],強直性肌營養不良、脊髓型肌萎縮癥、色素性視網膜炎、囊性纖維化、帕金森式癥、荷爾蒙缺陷等復雜疾病[15, 16]。
本文通過分析Fkbp51 KO與WT小鼠肝臟表達譜,研究Fkbp51基因敲除對其他基因可變剪接(內含子保留及外顯子跳躍)的影響,并分析差異可變剪接參與的生物學過程及代謝通路,以期進一步探索Fkbp51在肝臟糖、脂代謝中的重要作用。
1 材料和方法
1.1 實驗動物及測序
2月齡同窩SPF級Fkbp51 KO與WT雄鼠(C57BL/6),各3只,由中國醫學科學院醫學實驗動物研究所提供[SYXK(京)2014-0029,SCXK(京)2014-0004],體重為20~24 g。動物實驗方案經過中國醫學科學院醫學實驗動物研究所實驗動物使用與管理委員會(IACUC)批準。脫頸處死,取肝臟,Trizol法提取總RNA,樣品交由華大基因建庫和測序。
1.2 差異可變剪接的識別
原始序列去除低質量、污染序列及接頭后,得到干凈序列;用TopHat將干凈序列比對到參考基因組,分析比對結果,尋找外顯子間的結合位點。首先篩選出KO和WT表達量有差異的可變剪接(t檢驗,P< 0.05)。然后篩選新引入和缺失的可變剪接,所有可變剪接位點支持數都大于5。
1.3 差異可變剪接的GO功能和KEGG代謝通路分析
將發生差異可變剪接的基因通過DAVID在線工具[17, 18],以全基因組為背景進行GO功能及KEGG 代謝通路[19]富集分析。 統計方法為Fisher’s exact test,Benjamini算法校正P值,校正后p-value <0.05為差異有顯著性。
1.4 受差異可變剪接影響的通路
內含子保留和外顯子跳躍共同參與了生物學過程和代謝通路的調節。通過對差異外顯子跳躍和內含子保留基因做GO生物學過程和KEGG代謝通路注釋,來確定相關基因以SE、 RI、或兩者的組合來參與生物學過程和代謝通路。
2 結果
2 差異可變剪接基因的識別
基因發生差異外顯子跳躍事件統計如下:KO與WT共有,KO特異,WT特異的基因數分別為991、311、571個;發生內含子保留事件,KO與WT共有,KO特異,WT特異的基因數分別為1503、463、705個。以WT為例,我們定義Fkbp51KO造成WT中可變剪接消失的基因為WT特異基因、缺失的可變剪接為WT特異可變剪接;同理,我們也定義Fkbp51基因敲除造成KO中可變剪接出現的基因為KO特異基因、新增加的可變剪接為KO特異可變剪接。在KO和WT都出現相同可變剪接的基因為共有基因,可變剪接為共有可變剪接。統計分析發現在內含子保留和外顯子跳躍事件中,占主要部分的是共有可變剪接,其次是WT特異、KO特異。
2.2 差異可變剪接的識別
以基因可變剪接位點是否表達以及表達量的多少為標準,我們篩選出了KO和WT差異共有(都表達,但表達量有差別)、KO特異、WT特異可變剪接。外顯子跳躍中,差異共有、KO特異、WT特異可變剪接分別為122、15、12個。內含子保留中,分別為498、104、144個。Fkbp51基因敲除對SE和RI都有影響,但對RI影響較大,共有746個差異RI可變剪接,而SE只有139個。對SE和RI來說,Fkbp51基因敲除主要影響共有可變剪接的表達量,其次造成新可變剪接的引入和原有可變剪接的缺失。
2.3 差異可變剪接的GO功能注釋
為了研究發生差異可變剪接的基因的功能,我們用DAVID做了GO功能富集分析,如表1、2。外顯子跳躍基因的GO注釋主要富集到四個過程,一是脂肪代謝衍生物相關的代謝過程,如脂肪酸,有機酸,甾體和膽固醇的代謝過程;二是免疫相關,如急性炎癥反應中血漿蛋白的激活,體液免疫應答,急性炎癥反應,激活免疫應答;三是補體的激活,包括補體激活的經典和替代途徑;四是蛋白加工和成熟,如多肽鍵裂解的蛋白成熟;其他包括輔酶的代謝過程和芳香族化合物的分解代謝過程。在細胞組分的定位中,主要位于過氧化物酶體,線粒體和細胞質核周圍。氧化物酶體的主要功能是利于脂肪、醇、氨基酸的分解,其主要的分子功能是與輔因子、維生素、脂質結合。
在表2中,差異內含子保留基因主要富集的生物學過程為mRNA的代謝過程、類固醇的生物合成過程、一碳代謝過程,肌動蛋白細胞骨架組織方式的調控,細胞氨基酸衍生物代謝過程,對未折疊蛋白的反應。主要的細胞組分位于核膜,脂質雙層膜的細胞器,最靠近能動細胞的移動方向部分,內質網。主要的分子功能包括與核苷酸的結合、RNA結合、GTP酶結合、酸性氨基酸連接酶活性、RAB鳥嘌呤核苷酸交換因子活性和腺嘌呤核苷酸結合。mRNA的代謝過程中,結合活性是非常重要的。

表1 KO與WT發生差異外顯子跳躍基因的GO注釋
注:P-值<0.05。
Note.P-value<0.05.
2.4 差異可變剪接的KEGG代謝通路分析
為了研究差異可變剪接基因參與哪些代謝通路,我們用DAVID做了KEGG代謝通路分析,如表3、4所示。差異SE基因主要富集的通路有補體和凝血級聯,初級膽汁酸合成,細胞色素P450介導的異源物代謝,PPAR信號通路。在表3中,富集在PPAR信號通路上的cyp450家族有Cyp4a14,主要參與脂肪酸的氧化;Apoc3參與脂質轉運;Scp2和Ehhadh參與脂肪酸的氧化。在細胞色素P450介導的異源物代謝過程中,Gstz1和Gsta3是谷胱甘肽S-轉移酶,參與對毒物,致癌物質和藥理活性親電子化合物的轉化解毒;Gsta3屬于谷胱甘肽S-轉移酶α家族,編碼的酶具有谷胱甘肽過氧化物酶活性;Gstz1參與氧化應激反應;Cyp2c50催化花生四烯酸分解成中鏈和ω末端羥酸;Ugt1a6a編碼葡萄糖醛酸轉移酶,轉化小的親脂性分子如固醇激素和膽紅素等。

表2 KO與WT發生差異內含子保留基因的GO注釋
注:P-值<0.05。
Note.P-value<0.05.
從表4可以看出,差異RI基因主要富集的通路與氨基酸代謝相關,包括半胱氨酸和蛋氨酸、含硒氨基酸代謝;另外一些富集與免疫相關,包括急性髓系白血病和慢性粒細胞白血病;還有一些富集到趨化因子信號轉導通路,包括補體和凝血級聯反應和黑素的合成等。已有研究發現,在腫瘤中異常可變剪接的比例高于正常細胞,而且可變剪接類型也不同。Skotheim的腫瘤研究結果表明,原本在正常細胞中占較小比例的內含子保留類型在腫瘤中上升,而外顯子跳躍的比例則下降[20]。
2.5 受差異可變剪接影響的通路
Fkbp51基因敲除對生物是一種非生物脅迫,Fkbp51KO后,機體需要通過一些基因的調節來彌補Fkbp51基因的功能,而可變剪接就是這樣一種轉錄水平的調節。在表5中,在類固醇代謝過程,急性炎癥反應中血漿蛋白的激活,補體激活,有機酸的代謝過程,羧酸代謝過程,胺的代謝過程,急性炎癥反應,多肽鍵裂解的蛋白成熟等過程,都發現了基因通過不同的可變剪接來參與生物過程和代謝通路。
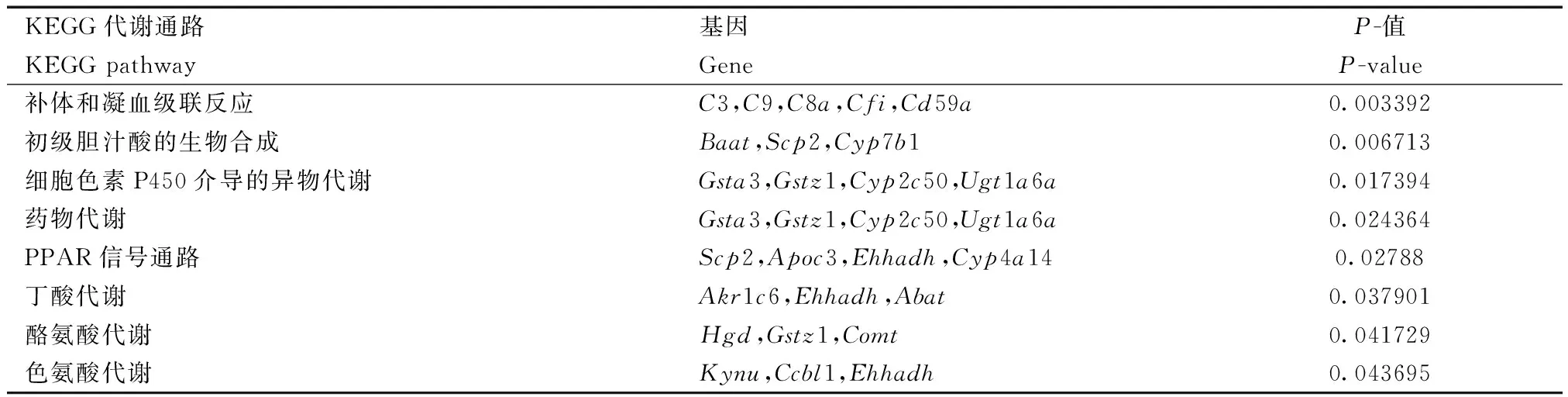
表3 KO與WT發生差異外顯子跳躍基因的KEGG pathway
注:P-值<0.05。
Note.P-value<0.05.
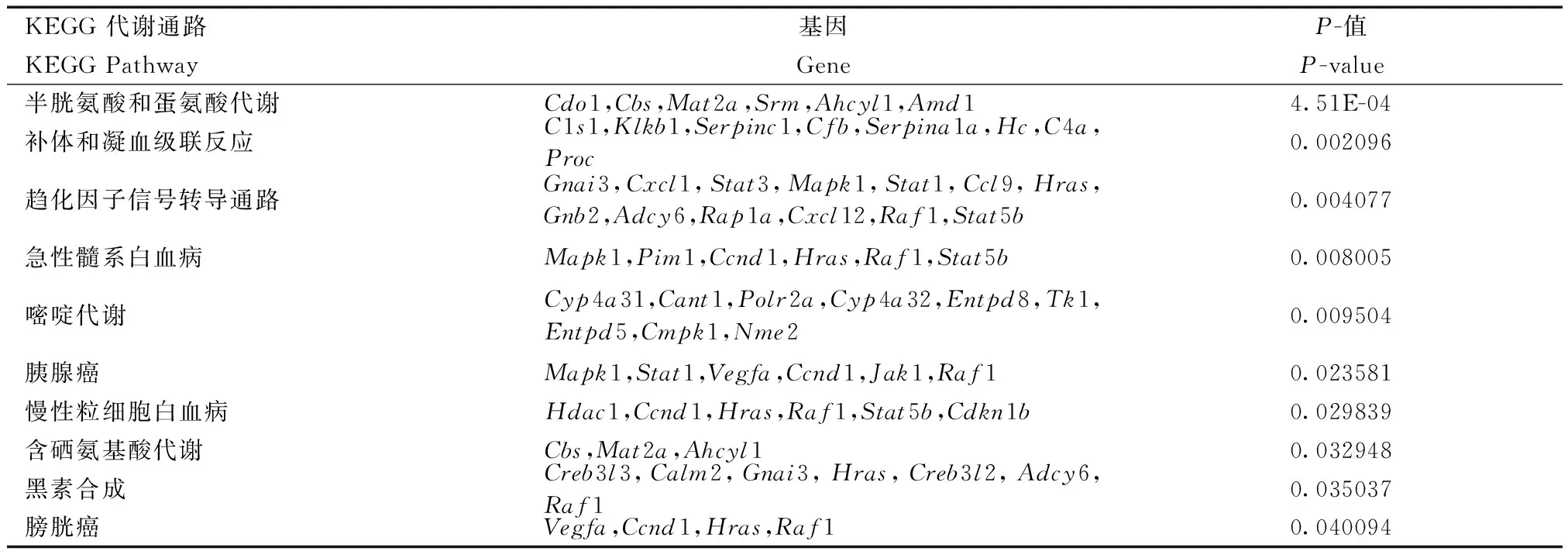
表4 KO與WT發生差異內含子保留基因的KEGG pathway
注:P-值<0.05。
Note.P-value<0.05.
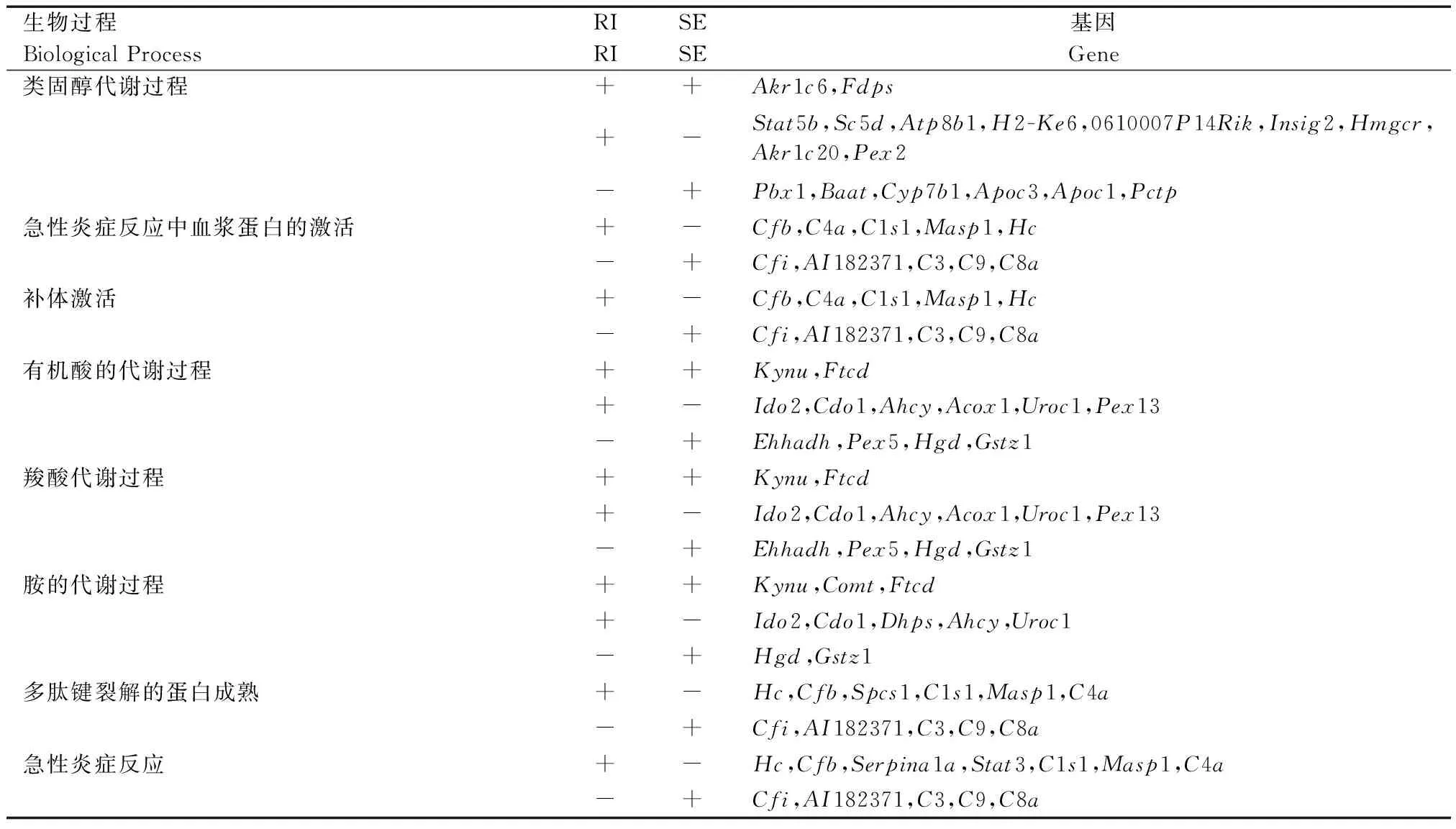
表5 幾個通路相關的基因可變剪接事件
注:‘+’代表該類型可變剪接事件出現,‘-’代表該類型可變剪接事件未出現。
Note. ‘+’ represents the occurrence of alternative splicing event, ‘-’ represents the opposite.
3 討論
Fkbp51在糖、脂代謝過程中所扮演的具體角色目前仍不清楚,其作用機制可能涉及多個方面,這是一個多基因、多途徑、多步驟、多信號通路相互作用和相互影響的過程。因此,基于RNA測序比較Fkbp51 KO與WT小鼠肝臟表達譜,鑒定差異可變剪接并分析其功能,是揭示Fkbp51基因在糖、脂代謝中作用的一個重要突破口。
我們觀察到,Fkbp51KO小鼠,高脂飼喂時,體重明顯減輕,肝臟脂滴減少。在基礎和高脂誘導條件下,氧氣消耗量,二氧化碳產生量,呼吸交換速率及產熱量均高于WT[7]。已有研究報道可變剪接因子和其他一些基因的敲除能夠影響可變剪接[21]。因此我們想了解Fkbp51基因敲除是否能影響其他基因的可變剪接,進而影響Fkbp51表型相關的生物過程和代謝通路。
首先,通過分析數據共篩選出139個差異外顯子跳躍。通過進一步的基因功能(GO)和KEGG代謝通路分析,我們發現這些差異基因主要參與脂肪衍生物相關的代謝過程,如脂肪酸,有機酸,類固醇和膽固醇的代謝過程。其中,PCTP是磷脂轉移蛋白,具有脂質結合和磷脂酰膽堿轉運活性;APOC1是載脂蛋白,在結合磷脂酰膽堿和抑制磷脂酶中發揮重要作用;BAAT是膽汁酸輔酶A,具有受體結合和棕櫚酰輔酶A水解酶的活性,在膽汁分泌中扮演重要角色;Akr1c6編碼酮類還原酶,利用NADH/NADPH催化醛與酮之間的轉換;Apoc3是載脂蛋白,與脂質結合和膽固醇結合相關。Fdps編碼法尼基焦磷酸合酶,可以催化合成香葉基焦磷酸酯和法尼基焦磷酸酯,而法尼基焦磷酸酯是膽固醇和甾醇生物合成過程中重要的中間物質。Pex5是過氧化物酶體因子5,與酶和蛋白的N端結合相關。Hgd是尿黑酸1,2雙加氧酶,能催化尿黑酸的加氧。Ehhadh編碼的蛋白是一個雙功能酶,是過氧化物酶體β-氧化途徑4種酶的一種。Gstz1是谷胱甘肽S-轉移酶,具有谷胱甘肽過氧化物酶的活性。因此深入研究Fkbp51與這些基因的關系,對于探究Fkbp51在脂代謝中的作用具有重要意義。
其次,我們篩選出746個差異內含子保留,這些差異基因主要參與肌動蛋白細胞骨架組織方式的調控,氨基酸及其衍生物的代謝,與脂代謝和糖代謝相關性不大,但與類固醇的代謝相關。這說明和內含子保留相比,外顯子跳躍在脂代謝和糖代謝中的作用更重要。內含子保留可能更多地參與翻譯前轉錄過程的調控,如通過無義介導的mRNA降解,而不是通過翻譯成蛋白質來發揮作用。
本文主要討論了Fkbp51基因敲除對可變剪接的影響,對研究其在脂肪代謝中的作用提供一個新的思路。為了驗證Fkbp51基因敲除造成的可變剪切的生物學意義,下一步我們將重點研究Ehhadh、Gsta3、Comt、Glo1、Acox1、Ahcy、Rpl17和Hc這些可變剪接體的變化,通過3‘或5’ RACE方法,對相應剪切體克隆,通過體外和體內實驗進一步研究這些可變剪切體的生物學功能,以期有新的發現。
[1] Zannas AS, Wiechmann T, Gassen NC,etal. Gene-Stress-Epigenetic Regulation of FKBP5: Clinical and Translational Implications[J]. Neuropsychopharmacology,2016, 41(1): 261-274.
[2] Hubler TR, Scammell JG. Intronic hormone response elements mediate regulation of FKBP5 by progestins and glucocorticoids[J]. Cell Stress Chaperones,2004, 9(3): 243-252.
[3] U M, Shen L, Oshida T,etal. Identification of novel direct transcriptional targets of glucocorticoid receptor[J]. Leukemia,2004, 18(11): 1850-1856.
[4] Paakinaho V, Makkonen H, Jaaskelainen T,etal. Glucocorticoid receptor activates poised FKBP51 locus through long-distance interactions[J]. Mol Endocrinol,2010, 24(3): 511-525.
[5] Stechschulte LA, Hinds TJ, Khuder SS,etal. FKBP51 controls cellular adipogenesis through p38 kinase-mediated phosphorylation of GRalpha and PPARgamma[J]. Mol Endocrinol,2014, 28(8): 1265-1275.
[6] Toneatto J, Guber S, Charo NL,etal. Dynamic mitochondrial-nuclear redistribution of the immunophilin FKBP51 is regulated by the PKA signaling pathway to control gene expression during adipocyte differentiation[J]. J Cell Sci,2013, 126(Pt 23): 5357-5368.
[7] 張曼,邱彬,曹勇,等. 共伴侶蛋白FKBP51在高脂誘導肥胖中的作用[J]. 中國比較醫學雜志,2015(07): 53-58.
[8] Schutt C, Nothiger R. Structure, function and evolution of sex-determining systems in Dipteran insects[J]. Development,2000, 127(4): 667-677.
[9] Black DL. Mechanisms of alternative pre-messenger RNA splicing[J]. Annu Rev Biochem,2003, 72: 291-336.
[10] Maniatis T, Tasic B. Alternative pre-mRNA splicing and proteome expansion in metazoans[J]. Nature,2002, 418(6894): 236-243.
[11] Carstens RP, Wagner EJ, Garcia-Blanco MA. An intronic splicing silencer causes skipping of the IIIb exon of fibroblast growth factor receptor 2 through involvement of polypyrimidine tract binding protein[J]. Mol Cell Biol,2000, 20(19): 7388-7400.
[12] Wang HY, Xu X, Ding JH,etal. SC35 plays a role in T cell development and alternative splicing of CD45[J]. Mol Cell,2001, 7(2): 331-342.
[13] Maquat LE. Nonsense-mediated mRNA decay: splicing, translation and mRNP dynamics[J]. Nat Rev Mol Cell Biol,2004, 5(2): 89-99.
[14] Caceres JF, Kornblihtt AR. Alternative splicing: multiple control mechanisms and involvement in human disease[J]. Trends Genet,2002, 18(4): 186-193.
[15] Wang ET, Sandberg R, Luo S,etal. Alternative isoform regulation in human tissue transcriptomes[J]. Nature,2008, 456(7221): 470-476.
[16] Pan Q, Shai O, Lee LJ,etal. Deep surveying of alternative splicing complexity in the human transcriptome by high-throughput sequencing[J]. Nat Genet,2008, 40(12): 1413-1415.
[17] Huang DW, Sherman BT, Lempicki RA. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources[J]. Nat Protoc,2009, 4(1): 44-57.
[18] Huang DW, Sherman BT, Lempicki RA. Bioinformatics enrichment tools: paths toward the comprehensive functional analysis of large gene lists[J]. Nucleic Acids Res,2009, 37(1): 1-13.
[19] Ogata H, Goto S, Sato K,etal. KEGG: Kyoto Encyclopedia of Genes and Genomes[J]. Nucleic Acids Res,1999, 27(1): 29-34.
[20] Skotheim RI, Nees M. Alternative splicing in cancer: noise, functional, or systematic?[J]. Int J Biochem Cell Biol,2007, 39(7-8): 1432-1449.
[21] Moroy T, Heyd F. The impact of alternative splicing in vivo: mouse models show the way[J]. RNA,2007, 13(8): 1155-1171.
Effects ofFkbp51 gene knockout on the alternative splicing of liver transcriptome in mice
ZHOU Zhi-qiang, YANG Zhi-wei*, YONG Wei-dong*
(Institute of Laboratory Animal Science,Chinese Academy of Medical Sciences (CAMS) & Comparative Medical Center,Peking Union Medical College (PUMC); Key Laboratory of Human Disease Comparative Medicine, Ministry of Health; Key Laboratory of Human Disease Animal Models, State Administration of Traditional Chinese Medicine, Beijing 100021, China)
Objective The purpose of this study is to understand the influence ofFkbp51 gene knockout on alternative splicing of mRNA in liver tissues. Methods mRNAs of liver from bothFkbp51 knockout(KO) and wild type(WT) mice were isolated. mRNA expression profiling was performed using RNA-seq reads. The mRNA reads produced from RNA-seq was analysed by TopHat for alternative splicing. Exon skipping and intron retetion were identified according to alignment analyses. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment were analyzed using online tools DAVID. All genes which had differently expressed alternative splicing between WT and KO were annotated using NCBI database. Results (1)Fkbp51 KO can introduce new mRNA alternative splicing; (2)Fkbp51 KO also cause mRNA alternative splicing expression change; (3) According to GO and KEGG analysis, it was found that the mRNA alternative splicing changes mainly enriched in fat metabolism, immune, bile acid secretion, and PPAR signaling pathway etc. (4) The genes which have different intron retetion events are mainly involved in regulation of actin cytoskeleton organization and amino acids metabolism. ConclusionsFkbp51 gene knockout can cause the change of mRNA alternative splicing, and thereby affect the metabolism of liver in mice.
FKBP51 knockout mice;Liver;RNA-seq;Alternative splicing;Intron retention;Exon skipping
艾滋病和病毒性肝炎傳染病重大專項(2014ZX10004002);國家自然科學基金(81272273)。
周志強(1986-),男,碩士生,研究方向:基因與發育生物學。E-mail: 522845911@qq.com
雍偉東(1968-),男,研究方向:生殖與發育生物學。Email: wyong@cnilas.org; 楊志偉(1969-),男,研究方向:高血壓。Email: zhw_yang@hotmail.com
R-33
A
1671-7856(2017) 05-0031-06
10.3969.j.issn.1671-7856. 2017.05.009
2016-11-15
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
音樂探索(2022年2期)2022-05-30 21:01:37
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
山東女子學院學報(2014年6期)2014-03-01 02:24:55
