擬南芥 WOX10 基因啟動子的克隆與序列分析
2017-07-03 13:21:58王占軍黃士平歐祖蘭徐忠東韓久盼余文敬
黃山學院學報 2017年3期
關鍵詞:植物
王占軍,黃士平,歐祖蘭,徐忠東,韓久盼,余文敬
(合肥師范學院 生命科學學院,安徽 合肥,230601)
擬南芥 WOX10 基因啟動子的克隆與序列分析
王占軍,黃士平,歐祖蘭,徐忠東,韓久盼,余文敬
(合肥師范學院 生命科學學院,安徽 合肥,230601)
為探索功能未知的擬南芥 WOX10 基因的表達規律,開展對擬南芥 WOX10 基因啟動子的分子克隆及測序分析,運用 PlantCARE 在線工具預測測序結果中可能含有的順式作用元件。結果表明,除 TATA-box 和 CAAT-box 外, 擬南芥 WOX10 基因啟動子含有生長素、 赤霉素和脫落酸等植物激素以及光信號,干旱、鹽分和壓力等脅迫相關的順式作用元件。該研究結果有益于解析擬南芥 WOX10 基因的轉錄調控規律及其功能。
WOX10 基因;擬南芥;啟動子;順式作用元件
WUSCHEL-RELATED HOMEOBOX (WOX)是僅存在于植物細胞內的一類編碼相關轉錄因子的基因 家 族[1]。 1996 年 ,Laux 等[2]利 用 EMS 誘 變 的 方 法 ,在擬南芥中克隆并鑒定出 WUS基因; 該基因編碼291 個氨基酸, 具有維持植物莖端分生組織中干細胞數量的功能。WUS 的同源異型結構域(Homeodomain 區) 是由 66 個氨基酸殘基所構成的特殊空間結構(Helix 1-loop-helix 2-turn-helix 3),該 結構及 其 特 殊 功 能 引 起 了 研 究 人 員 的 廣 泛 關 注[3]。Haecker 等[4]通 過 對 擬 南 芥 基 因 組 的 開 放 閱 讀 框(ORF)進行同源性比對分析,共鑒定出 14 個基因的ORF 區域與 WUS 基因的 Homeodomain 區存在高度的一致性或相似性, 這些基因被命名為 WOX1-WOX14。 目前,WOX 基因家族已在擬南芥、水稻、葡萄 、高 粱 、玉 米 等 30 多 種 植 物 中 鑒 定 出 來[5]; 根 據 功能分析的結果,可將 15 個基因家族成員分為調節植物 干細 胞活動[6]、控 制 細 胞 胚 胎發生[7]、調 節 側 生 器 官的 生 長 和 發 育[8]以 及 功 能 未 知 WOX10 基 因 共 4 種類型。
啟動子是基因表達調控系統中重要的順式作用元件,控制著基因表達的起始時間與表達程度,是基因表 達 活性的 “開關 ”[9]。 啟 動子 一 般是位 于基因編 碼區 5'端上游的一段特殊的核苷酸序列,它包含 RNA聚合酶以及一個或多個特定轉錄因子的結合位點,結 構 上 通 常 包 括 核 心 啟 動 子 元 件 (TATA-box、CAAT-box)和應答調控元件,參與轉錄的起始與調控[10]。
鑒于擬南芥 WOX10 基因的功能目前尚未見報道, 本研究針對 WOX10 基因的啟動子進行克隆及分析,獲得該啟動子區域基因表達調控網絡的一系列調控元件;此外,旨在基于 WOX10 基因啟動子序列的分析結果,為進一步研究 WOX10 基因的表達規律和基因功能提供理論依據。
1 實驗部分
1.1 啟動子序列的獲取
根 據 NCBI (National Center for BiotechnologyInformation,美國國立生物 技術信息中心)公布 的 擬南芥全基因組測序結果,利用 Map Viewer 確定擬南芥 WOX10 (簡寫為:AtWOX10) 基因序列的位置信息,下 載該 基因 翻 譯起始位 點 (ATG)上 游 2000 bp的啟動子序列(簡寫為:pAtWOX10)。
1.2 實驗試劑及方法
1.引物設計:依據 pAtWOX10 核苷酸序列,使用Primer Premier v5.0 軟件設計啟動子克隆引物,具體信息如下:pAtWOX10–F:AATAAACTAAAGATCCCTTTATTCATAAACAAAAG (從 5' 至 3' 端 ),pAt-WOX10-R:CTCCTCTTTCACCACTTTGCCTTCTC(從5'至 3'端)。
2. 利用 BioTeke 試劑盒提取擬南芥幼苗的基因組 DNA, 利用瓊脂糖凝膠電泳法檢測提取 DNA 的質量。
3. 使用 KOD 高保真酶進行 pAtWOX10 基因啟動子序列的 PCR 擴增,反應體系的總體積為 50μL,具 體 包 括 :10 ×PCR Buffer for KOD-Plus-Neo,5.0μL;2 mM dNTPs,5.0μL;25mM MgSO4,3.0μL;pAtWOX10 – F 引 物 ,1.5μL;pAtWOX10 – R 引 物 ,1.5μL;KOD-Plus-Neo 酶 1.0 μL; 擬 南 芥 DNA,3.0μL;ddH2O,30.0μL。
4.PCR 反應程序為:94℃預變性 5min; 循環內:94℃變 性 30sec,50℃退 火 30sec,72℃延 伸 2min,執行 30 次循環;72℃延伸 10 min。
5.PCR 產物檢測:使用 1%的瓊脂糖凝膠中檢測PCR 產物的大小及質量。
6.膠回收及連接、轉化:針對 PCR 產物大小與預期目標產物相符的條帶進行切膠,使用 Axygen 公司的 DNA 凝膠回收試劑盒進行膠回收,將膠回收產物加 Poly (A) 尾 處 理 后 , 連 接 至 TaKaRa 公 司 的pMDTM19-T 載體,轉化感受態細胞,菌液 PCR 檢測無誤后送至上海英俊生物技術有限公司(簡寫為:英俊公司)測序。
1.3 序列分析方法
運 用 PlantCARE (http://bioinformatics.psb.ugent. be/webtools/plantcare/html/) 在 線 分 析 軟 件 對 At-WOX10 基因啟動子的測序結果進行分析。
2 結果與討論
2.1 AtWOX10 基因及其啟動子的位置信息
依據 NCBI 數據庫中獲得的 AtWOX10 基因及其啟動子的相關信息,揭示 AtWOX10 基因位于擬南芥的 1 號 染色體上(如圖 1A),利 用 GSDS(http://gsds.cbi.pku.edu.cn/)基因結構分析工具解析 AtWOX10基因序列后,發現 AtWOX10 基因全長 783bp,其中編 碼 區 長 度 為 594bp, 含 有 3 段 外 顯 子 區 域(7184485-7184642;7184731-7185129;7185221-7185267) 和兩段內含子區域 (7184643-7184730;7185130-7185220)(見圖 1B 和表 1)。
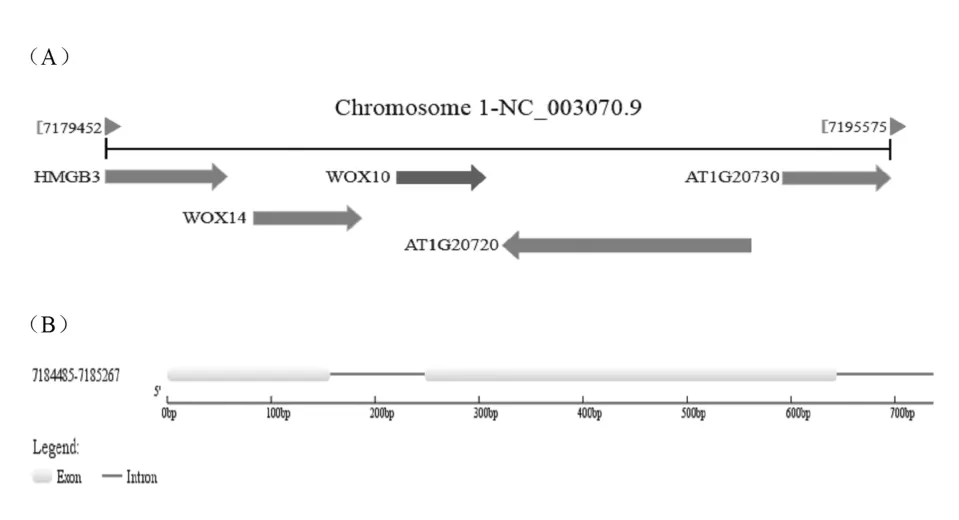
圖1 AtWOX10 基因在染色體上位置(A)及基因結構(B)示意圖
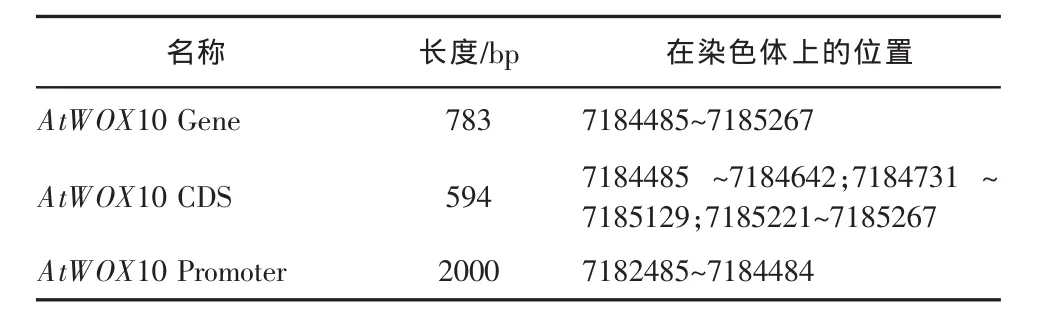
表1 AtWOX10 基因及其啟動子信息
2.2 AtWOX10 基因啟動子的克隆

圖2 AtWOX10 基因啟動子序列的PCR 產物(A)和膠回收產物(B)
利用具有高保真性的 KOD 聚合酶和特異性引物克隆 AtWOX10 基因的啟動子序列, 由于 PCR 反應體系的退火溫度較低,造成產物出現了雜帶,針對PCR 產物進行膠回收純化后, 凝膠電泳檢測的條帶單一。
2.3 AtWOX10 基因啟動子的序列分析
英俊公司測序結果顯示, 原長為 2000bp 的 At-WOX10 基因啟動子, 實際克隆產物的長度為 1999bp, 在 NCBI 公 布 的 AtWOX10 基 因 啟 動 子 -1922 bp 的位置缺失 1 個 A 堿基。 運用 PlantCARE 在線工具分析 AtWOX10 基因啟動子的測序結果,詳細信息如表 2 所示。 在 ATG 上游 500bp 長度內可能存在 4個 TATA-box(-100;-240;-280;-473)、7 個 CAAT-box(-163;-199;-221;-233;-249;-363;-391),具有真核生物核心啟動子元件典型的特征,二者共同介導的 RNA 聚合酶參與的 WOX10 基因轉錄過程的起始與調控,并且在該序列的多個位點重復出現,這對于提升基因對外界影響因素變化應答速度及反應效率是必不可少的。此外,該序列還存在多種特異性的順式調控元件,包括光誘導應答反應、激素誘導應答反應、脅迫誘導應答反應以及特異性表達產物調控等多方面功能;其中光應答元件共 7 種,包括 AE-box、E-box、G-box、GT-1、PIF3、SORLIP1AT 和-10 promoter element。 -10 promoter element是對藍光、白光和紫外光應答的元件,在植物的花器官發育調控中起著重要調控作用。 激素應答調控元件共 8 種, 包括 ARF、ASF1、AtMYB2、DPBF、GARE、MYBATRD22、TGA-element和 W-BOX,其中生長素應答元件 ARF 與赤霉素應答元件 GARE 常在植物的根部轉錄調控中發揮作用。脅迫誘導應答元件共 9 種,包括 ABRE、ACGT、LTRE、MYB1AT、MYB1LEPR、MYB2、MYBRE、MYC和 SREATMSD,其中 MYB 類水分脅迫與 LTRE 低溫脅迫誘導元件是根部生長發育必不可少的調控元件。此外,還發現如調控花器官部位相關基因表達的應答元件 AP1 和 MYBPLANT,根發育調控和木質部形成相關的基因表達的應答元件 RHERPATEXPA7 和 XYLAT,以及細胞周期蛋白表達的調控元件MYBCOREATCYCB1 等其他有關轉錄起始和特異性表達產物合成控制的應答作用元件,針對這些表達部位明確的應答作用元件結果進行分析,將有利于深入解析 AtWOX10 基因的表達調控規律及其基因功能。

表2 AtWOX10 基因啟動子區的順式作用元件預測結果
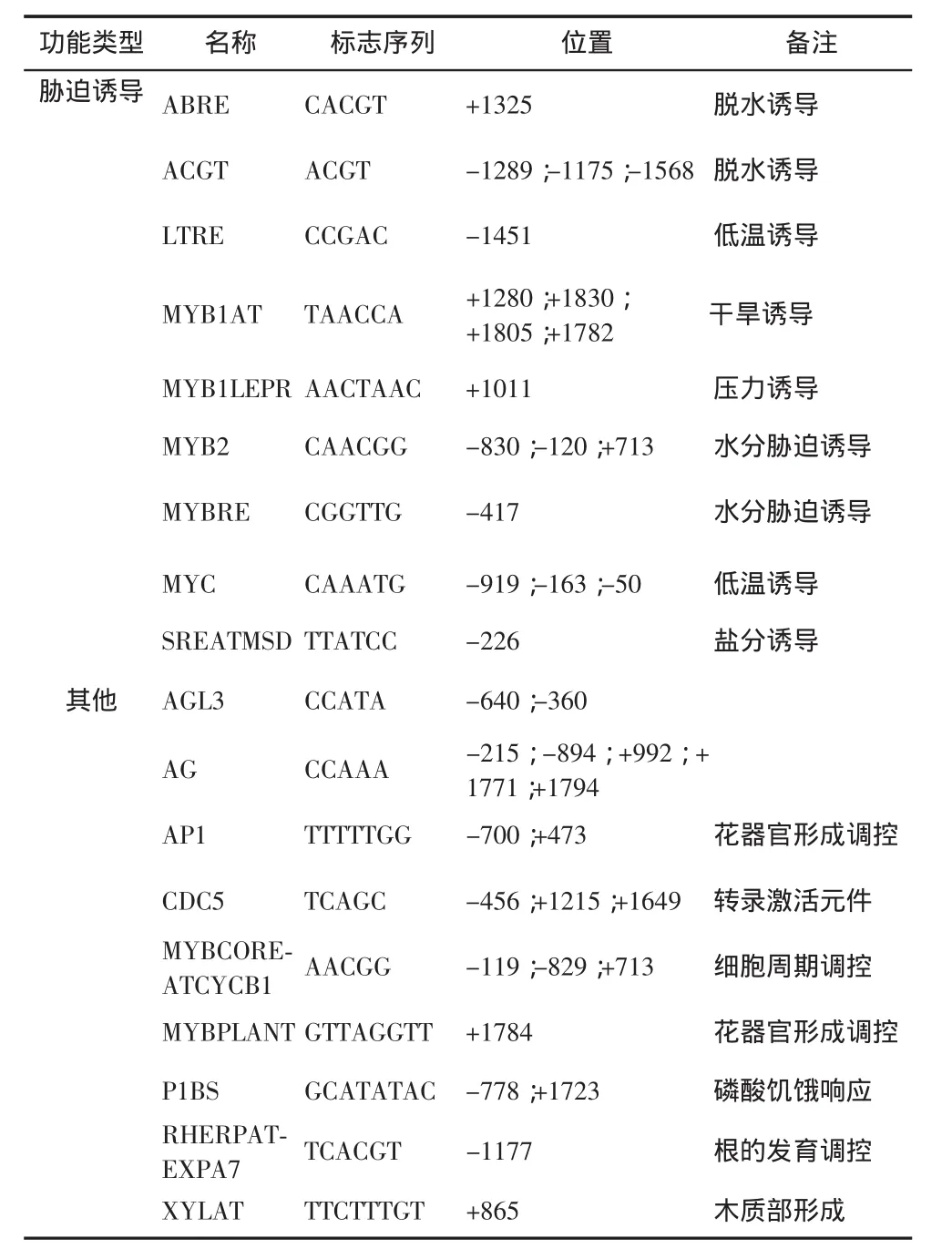
注 :(-)表 示 pAtWOX10 序 列 ,(+)表 示 與 pAtWOX10 互補序列。
3 結論及展望
由上述 AtWOX10 基因啟動子中相關順式作用元件分析結果可知,當外界環境處于干旱、高鹽以及壓力等逆 境脅 迫時,AtWOX10 基因可能會被激活(MYB1LEPR、MYB1AT、SREATMSD);也可能存在對藍色、 白色和紫外光等產生相應的應答機制(-10 promoter element);能夠被諸如 生長 素 、赤 霉素 和 脫落酸等植物生長激素誘導表達(ARF、TGA-element、GARE、DPBF); 同時,AtWOX10 基因啟動子還具有一些特殊的應答作用元件,例如花器官中基因特異性表達的因子 AP1。 綜上所述,AtWOX10 基因轉錄調控可能受到光信號,生長素、赤霉素和脫落酸等植物激素以及脅迫等多種信號途徑的協同作用; 但是,At-WOX10 基因具體的轉錄調控規律有待進一步研究。
本文對 AtWOX10 基因啟動子進行了克隆及序列分析,但關于其表達的具體部位,真實存在哪些與其表達相關的光照、激素和溫度等因素,有待下一步通過構建植物表達載體,并將其轉化至擬南芥中加以驗證;同時,本研究也為 AtWOX10 基因的功能解析提供了參考信息。
[1]Lian G,Ding Z,Wang Q,et al.Origins and evolution of WUSCHEL-related homeobox protein family in plant kingdom[J].The Scientific World Journal,2014:534140.
[2]Laux T,Mayer KF,Berger J,et al.The WUSCHEL gene is required for shoot and floral meristem integrity in Arabidopsis[J].Development,1996,122(1):87-96.
[3]Kamiya N,Nagasaki H,Morikami A,et al.Isolation and characterization of a rice WUSCHEL-type homeobox gene that is specifically expressed in the central cells of a quiescent center in the root apical meristem[J].The Plant Journal,2003,35(4):429-441.
[4]HaeckerA,Groβ -HardtR,Geiges B,etal.Expression dynamics of WOX genes mark cell fate decisions during early embryonic patterning in Arabidopsis thaliana[J]. Development,2004,131(3):657-668.
[5]Zhang X,Zong J,Liu J,et al.GenomeWide analysisof WOX gene family in Rice,Sorghum,Maize,Arabidopsis and Poplar[J].JournalofIntegrative PlantBiology,2010,52(11):1016-1026.
[6]Mayer KF,Schoof H,Haecker A,et al.Role of WUSCHEL in regulating stem cellfate in the Arabidopsisshoot meristem[J].Cell,1998,95(6):805-815.
[7]Palovaara J,Hakman I.Conifer WOX-related homeodomain transcription factors,developmental consideration and expression dynamic of WOX2 during Picea abies somatic embryogenesis[J].Plant Molecular Biology,2008,66(5):533-549.
[8]Vandenbussche M,Horstman A,Zethof J,et al.Differential recruitment of WOX transcription factors for lateral development and organ fusion in Petunia and Arabidopsis[J]. The Plant Cell,2009,21(8):2269-2283.
[9]聶麗娜,夏蘭琴,徐兆師,等.植物基因啟動子的克隆及其功能研究進展[J].植物遺傳資源學報,2008,9(3):385-391.
[10]王穎,麥 維 軍 ,梁承鄴,等.高 等 植 物 啟 動 子 的 研 究 進 展[J].西北植物學報,2004,23(11):2040-2046.
責任編輯:胡德明
The Cloning and Sequence Analysis of Promoter of WOX Gene Family in Arabidopsis thaliana
Wang Zhanjun,Huang Shiping,Ou Zulan,Xu Zhongdong,Han Jiupan,Yu Wenjing
(School of Life Sciences,Hefei Normal University,Hefei 230601,China)
In order to study the expression rule of WOX10 with unknown function in Arabidopsis thaliana,the promoter of WOX10 gene in Arabidopsis thaliana was cloned and sequenced.Cis-acting elements were predicted by PlantCARE in the promoter of AtWOX10 gene.The results showed that there were TATA-box,CAAT-box and some cis-acting elements in the promoter of AtWOX10 gene,including a variety of plant hormones like auxin,gibberellin and abscisic acid,light responsive element,and also cisacting elements related to drought,salt and stress.The results can also be useful for future studies of AtWOX10 gene in transcriptional regulation and function.
WOX10 gene;Arabidopsis thaliana;promoter;cis-acting element
Q74
:A
:1672-447X(2017)03-0058-04
2016-09-20
安徽省高校自然科學基金重點項目(KJ2015A186);安徽省自然科學基金面上項目 (1708085MC76);合肥師范學院人才科研啟動基金項目(2013rcjj01);合肥師范學院產學研項目(2014cxy33);合肥師范學院課程建設項目(2013hxk14)
王占軍(1984-),安徽壽縣人,合肥師范學院講師,研究方向為植物細胞分子生物學。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
