小桐子類固醇還原酶基因Jc5β-StR的克隆和表達分析
2017-07-12 18:05:57黃堯瑤文錦芬鄧明華龔明陳浩維
生物技術通報 2017年7期
黃堯瑤文錦芬鄧明華龔明陳浩維
(1. 昆明理工大學現代農業工程學院,昆明 650500;2. 昆明理工大學建筑與城市規劃學院,昆明 650500;3. 云南農業大學園林園藝學院,昆明 650201;4. 云南師范大學生命科學院,昆明 650500)
小桐子類固醇還原酶基因Jc5β-StR的克隆和表達分析
黃堯瑤1文錦芬2鄧明華3龔明4陳浩維1
(1. 昆明理工大學現代農業工程學院,昆明 650500;2. 昆明理工大學建筑與城市規劃學院,昆明 650500;3. 云南農業大學園林園藝學院,昆明 650201;4. 云南師范大學生命科學院,昆明 650500)
以小桐子(Jatropha curcas L.)cDNA為模版,克隆了Jc5β-StR基因的CDS序列,并對其序列進行了生物信息學分析。序列分析表明該基因包含1 173 bp開放閱讀框(ORF),編碼390個氨基酸。預測其編碼蛋白質的相對分子量為44.23 kD,理論等電點為5.22。Blast搜索結果及進化分析結果表明,Jc5β-StR與蓖麻5β-StR蛋白序列一致性最高(87%)且親緣關系最近。Jc5β-StR是一個短鏈還原酶,該蛋白編碼有一個短鏈還原酶家族(SDRs)的黃體酮5β還原酶(5β-POR),還含有SDRs的6個保守結構域。組織表達結果顯示Jc5β-StR在小桐子根、莖、葉、花、果皮和種子中都有表達,在種子中表達量最高,且隨種子發育過程表達量逐漸上升,在種子生長發育的后期(50 d)達到最高值后下降。非生物脅迫(PEG、NaCl、低溫和機械損傷)誘導下,Jc5β-StR的表達量都不同程度上調,推測其參與小桐子非生物脅迫響應。
小桐子;Jc5β-StR;克隆;生物信息學分析;表達分析
短鏈還原酶家族(Short-chain dehydrogenases/ reductases,SDRs)是迄今為止發現的最古老、最大的蛋白質超家族之一,存在各種生物形式中,如古細菌,原核生物,真核生物和病毒等[1-2]。SDR家族成員在脂類、氨基酸、碳水化合物、激素類、異生物質代謝和氧化還原反應傳感機制中發揮重要作用。它們通常催化NADP(H)依賴的反應,其底物包括多元醇、類維生素A、類固醇、脂肪酸的衍生物和異源物質等[3-4]。已報到的SDRs具有廣泛的生物學功能,不僅通過參與許多重要內固醇化合物的合成來調控植物生長發育,如油菜素內酯和強心苷等,還參與植物的脅迫應答。半定量RT-PCR分析證實,在草莓果實發育過程中,草莓短鏈還原酶基因FaSDR的表達量隨著果實的著色快速升高,揭示該基因可能參與草莓果實成熟調控[5]。擬南芥的一個短鏈還原酶基因AtHSD1通過調節油菜素內酯生物合成,來調控植物的生長和發育[6,7]。類固醇5α-還原酶(DET2)也屬于SDR家族一員,是油菜素類固醇物質生物合成途徑的限速酶,GbDET2基因在棉花纖維的起始和快速伸長過程中發揮著重要的作用[8]。從甘藍型油菜中克隆的β-酮脂酰輔酶A還原酶基因BnKCR1和BnKCR2在植株中普遍表達,并在種子與根系中的表達最豐富,在酵母中異源表達這兩個基因恢復了酵母脂肪酸延長酶的活性[9]。胡椒(CaMNR1)和擬南芥(AtSDR1)中的薄荷酮新薄荷醇還原酶(MNR)基因增強了植物對多種病原菌的抗性[10]。
類固醇5β還原酶5β-StR(3-o-Δ4,5steroid 5β-reductase)也是SDR家族的一員,能夠催化多種類固醇基質,在內固醇化合物生物合成途徑上發揮作用。目前對于植物5β-StR的研究報道較少,Herl等[11]在2009年報道了一個擬南芥內固醇5β還原酶基因At5β-StR,其蛋白序列和三維模型都與毛花洋地黃(Digitalis lanata)黃體酮5β還原酶(Dl5β-POR)相似度較高,5β-POR是一種類固醇化合物—強心苷生物合成途徑上的重要催化酶。半定量RTPCR發現擬南芥At5β-StR在莖和根中表達量最高,并且在甘露醇誘導20 h后的表達明顯增強,推測At5β-StR還參與植物滲透脅迫調控。同年,Edyta等[12]研究報道5β-StR因為其碳碳雙鍵活性,所以催化能力要強于5β-POR。
小桐子(Jatropha curcas L.)又叫做膏桐、小油桐、老胖果、油蘆子、麻瘋樹等。原產熱帶美洲,分布于我國廣東、云南、四川、貴州、廣西等省區[13]。小桐子果實含油率高達60%,可以提練出不含硫、無污染、符合歐四排放標準的生物柴油,是中國重點開發的綠色能源樹種[14]。小桐子不擇土壤,耐干旱瘠薄,又是干熱河谷地區荒山造林的優良樹種,具有廣泛的開發利用前景[15-16]。本研究從小桐子中克隆得到一個類固醇5β還原酶基因Jc5β-StR,進行蛋白質序列和功能結構上的探討,并分析其在不同組織和非生物脅迫下的表達變化,以期為進一步研究5β-StR基因的作用奠定基礎。
1 材料與方法
1.1 材料
供試材料小桐子(Jatropha curcas L.)取自于云南省昆明市昆明理工大學現代農業工程學院實驗基地。取小桐子根、莖、葉、花、果皮和授粉后不同發育階段(10、20、30、40、50和60 d)的種子,立即液氮速凍后保存在-80℃的環境中備用。每種組織從10株小桐子樹上取樣,來源于3株樹上的組織混合作為1個組織樣品,4次重復。
實驗處理材料為穩定成熟期的小桐子幼苗,取長勢基本一致的小桐子幼苗分別放置于NaCl(250 mmol/L)培養液、30% PEG 6000培養液和4℃培養箱中,機械損傷用消過毒的剪刀在每個小桐子葉片上各剪5下,注意不要剪斷葉片主脈。4種處理后的0(對照)、3、6、12和24 h取幼嫩的葉片。各處理3次重復,材料用錫箔紙包好,立即液氮速凍后保存在-80℃的環境中備用。
1.2 方法
1.2.1 RNA的提取和cDNA第一鏈的合成 小桐子RNA用TianGen公司植物總RNA提取試劑盒(RNAprep pure Kit,DP432)提取,cDNA鏈合成用大連寶生物公司(TaKaRa PrimeScript II 1st Strand cDNA Synthesis Kit)試劑盒,操作過程參考說明。產物放置于-20℃備用。1.2.2 基因克隆 Jc5β-StR基因克隆特異引物設計為Jc5β-StR-F(5'-CCTCAGCCACCAACTACTA-3')和Jc5β-StR-R(5'-GTTCAAATCAAGGCACAAT-3'),PCR擴增用大連寶生物公司的Taq酶(TaKaRa Ex Taq Hot Start Version)和5×PCR buffer,cDNA模板加入量為2 μL,操作過程均參考說明。PCR反應程序:94℃預變性5 min,94℃變性30 s,54℃退火30 s,72℃延伸40 s,35個循環,72℃延伸5 min。將PCR產物連接到pMD18-T并雙向測序,每個基因至少5個克隆。
1.2.3 cDNA序列及其編碼氨基酸分析 用NCBI網站中的ORF finder找出基因的開放讀碼框(open reading frame,ORF)和其BLASTp程序進行同源性分析;用DNAMAN軟件進行核酸翻譯和多序列對比,采用MEGA7.0 Neighbor-joining方法構建進化樹進行親緣關系分析。利用ProtParam在線軟件分析氨基酸殘基數目、組成和蛋白質的基本性質等參數。利用ExPASy在線分析蛋白質一級結構和預測蛋白質功能位點。通過在線軟件SOPMA預測蛋白質二級結構。用Modeling程序(http://swissmodel.expasy. org/)對蛋白質的三級結構進行預測。跨膜結構預測用TMHMM-2.0 server,信號肽預測用SignalP。
1.2.4 基因的表達分析 用實時熒光定量PCR方法進行基因表達分析。根據測序結果設計特異引物序列為Jc5β-StR-qF(5'-GATTTGCCCAGATTAGACGCT-3')和Jc5β-StR-qR(5'-CAATCCCTCCTTCTTCGCTAC-3')。內參基因序列設計為β-actin-F(5'-GCAGGCATCCACGAGACTACT-3')和β-actin-R(5'-GTCAGCAATACCAGGGAACATAG-3')。分別對反轉錄后不同組織和不同處理材料葉片cDNA進行實時熒光定量PCR分析。反應體系為:2 μL cDNA模板、12.5 μL Premix TaqTM、0.5 μL Jc5β-StR-qF、0.5 μL Jc5β-StR-qR、加dd H2O至25 μL。PCR反應條件為:94℃預變性3 min,94℃預變性30 s,59℃退火1 min,72℃延伸30 s,72℃延伸5 min,35個循環。根據廠商推薦,在羅氏LightCycler 480系統上進行。每個材料做3次獨立重復,采用2-ΔΔCT方法分析處理數據,在組織表達分析中用表達量最低的組織作為校正子,在種子發育過程中的時序表達分析用第一發育階段(10 d)的樣品做校正子[17]。用SPSS軟件進行顯著性分析,Excel 2013作圖。
2 結果
2.1 Jc5β-StR基因的克隆
用RT-PCR方法對Jc5β-StR進行克隆,瓊脂糖凝膠電泳后在凝膠成像儀上觀察得到一條大約1 200 bp的產物(圖1),對其測序后,用ORF finder軟件分析Jc5β-StR基因編碼氨基酸序列,完整的ORF框編碼區為1 173 bp,編碼蛋白包含390個氨基酸(圖2)。
2.2 蛋白質一級結構分析
通過ProtParam軟件分析表明,Jc5β-StR蛋白相對分子量為44.23 kD,理論等電點為5.22,酸性氨基酸殘基總數(Asp+Glu)為49個,堿性氨基酸殘基總數(Arg+Lys)為34個,不穩定系數為41.40,為不穩定蛋白。原子總數是6 160個,分子式為C2017H3032N520O578S13。脂肪系數為84.31,疏水系數為-0.299,為親水蛋白。
2.3 蛋白質二級結構及三級結構分析
通過在線軟件SOPMA對蛋白質進行二級結構預測,結果表明Jc5β-StR蛋白含有豐富的α螺旋(Alpha he1ix)占氨基酸殘基總數的40.77%,其次是無規則卷曲(Random coil)占氨基酸殘基總數的33.08%,延伸鏈(Extended strand)占氨基酸殘基總數的17.18%,β-轉角(Beta turn)占氨基酸殘基總數的8.97%(圖3)。
通過Prosite在線數據庫分析,發現Jc5β-StR蛋白序列含有3類功能位點包括2個N-糖基化位點(N-glycosylation site),7個豆蔻酰化位點(N-myristoylation site)和7個酪蛋白激酶Ⅱ磷酸化位點(Casein kinase II phosphorylation site)。登錄http://swissmodel.expasy.org/中的MODELING,對小桐子Jc5β-StR蛋白三級建模,結果顯示Jc5β-StR蛋白質三級結構模型以黃體酮5β還原酶(5β-POR)為模板,序列相似度為74.86%,建模結果如圖4所示。2.4 蛋白質亞細胞定位、跨膜結構和信號肽預測

圖1 Jc5β-StR基因電泳檢測圖
蛋白質亞細胞定位預測Jc5β-StR最有可能定位在細胞質。跨膜結構預測結果表明Jc5β-StR不具有跨膜結構域。信號肽預測結果表明Jc5β-StR不具有信號肽,說明它不是一個分泌蛋白。
2.5 Jc5β-StR序列一致性分析
通過NCBI Blast序列比對發現5β-StR基因在多種植物中均存在,Jc5β-StR與大戟科蓖麻(Ricinus communis)序列一致性最高,為87%;其次與胡桃科核桃(Juglans regia)序列一致性為83%,與梧桐科可可樹(Theobroma cacao)序列一致性為82%;與薔薇科白梨(Pyrus bretschneideri)序列一致性為81%;與蕓香科甜橙(Citrus sinensis)、錦葵科陸地棉(Gossypium hirsutum)、薔薇科蘋果(Malus domestica)和梅(Prunus mume)等序列一致性為80%;與大豆(Glycine max)、辣椒(Capsicum annuum)、油菜(Brassica napus)等其他幾十種植物序列一致性為75%-79%。Jc5β-StR與不同種類植物序列一致性較高,說明該基因在植物進化過程中較保守。
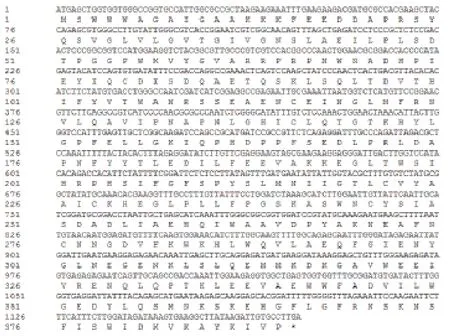
圖2 Jc5β-StR全長序列及其推導的氨基酸序列
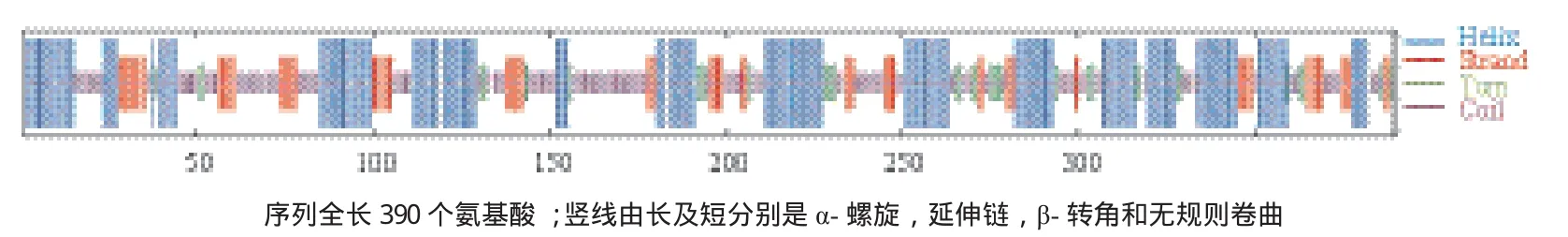
圖3 Jc5β-StR蛋白質二級結構
圖4 Jc5β-StR蛋白質三級結構
Jc5β-StR屬于NADB Rossmann超家族(圖5)。該蛋白編碼一個短鏈還原酶家族(SDRs)的黃體酮5β還原酶(5β-POR),Jc5β-StR也有含有SDRs的6個保守結構域(圖6),表明Jc5β-StR是一個短鏈還原酶。結構域Ⅰ-Ⅲ在所有NADP依賴的SDRs中都存在,是典型的SDRs結構域,余下的3種Ⅳ-Ⅵ結構域,不是所有的SDRs都存在,特別是Ⅴ結構域(NFYYxxED),是5β-POR類的典型結構域[11]。
2.6 Jc5β-StR進化分析
用MEAG7.0軟件對Jc5β-StR蛋白氨基酸序列和其他植物的5β-StR蛋白進行進化分析,結果表明,Jc5β-StR的進化基本符合植物分類學分類,并具有明顯的種屬特性,如屬薔薇科的蘋果和梨組成一個分支,Jc5β-StR與大戟科蓖麻聚在一枝(圖7),說明其與蓖麻親緣關系最近。
2.7 Jc5β-StR表達分析

圖5 Jc5β-StR保守結構域預測
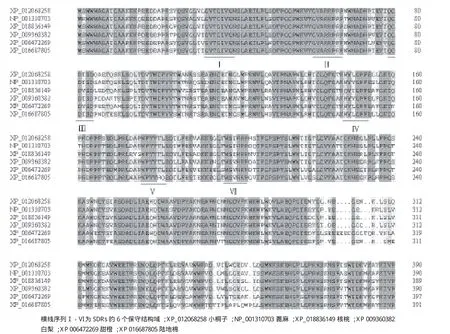
圖6 Jc5β-StR與不同物種5β-StR氨基酸序列對比
以β-actin作為內參,對小桐子Jc5β-StR在不同組織和種子不同發育階段的表達情況進行qRT-PCR分析,分析結果表明,Jc5β-StR在小桐子根、莖、葉、花、果皮和種子中都有表達,在種子和葉片中的表達量顯著高于其他組織。Jc5β-StR在種子發育過程中的定量分析(圖8)表明,該基因在種子整個生長發育期都有表達,在種子生長發育前期其表達水平不斷增加,在種子生長發育的后期(50 d)達到最高值,然后下降。
Jc5β-StR在PEG、NaCl、機械損傷和低溫4℃處理下表達都不同程度提高,PEG處理下表達水平變化顯著,在第12 h達到最高值,為對照樣品表達量的6倍左右。NaCl處理下12 h達到最高值,為對照樣品的1.8倍左右。機械損傷處理下第6 h達到最高值,為對照樣品的3.7倍左右。4℃處理下Jc5β-StR在第3小時表達水平就達到最高值,為對照樣品的2.5倍左右,之后緩慢下降(圖9)。
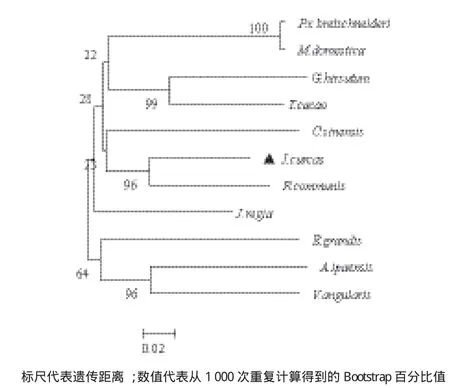
圖7 不同植物5β-StR氨基酸序列進化樹分析
3 討論
本研究通過RT-PCR方法從小桐子中克隆了一個小桐子類固醇5β還原酶基因(Jc5β-StR),分析其ORF大小為1 173 bp,編碼蛋白包含390個氨基酸。預測其相對分子量為44.23 kD,理論等電點為5.22。蛋白質亞細胞定位預測其定位在細胞質,沒有跨膜結構域,也不具有信號肽。Blast結果表明Jc5β-StR屬于NADB Rossmann超家族。其序列與同屬于大戟科的蓖麻5β-StR序列一致性最高,為87%,進化樹分析表明其與蓖麻5β-StR親緣關系最近。該蛋白編碼一個短鏈還原酶家族(SDRs)的黃體酮5β還原酶(5β-POR),也含有SDRs的6個保守結構域,證明Jc5β-StR是一個短鏈還原酶。
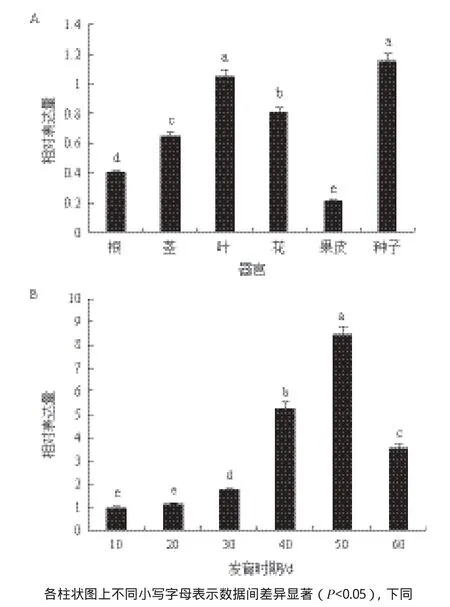
圖8 Jc5β-StR基因在不同組織(A)和種子不同發育時期(B)的表達
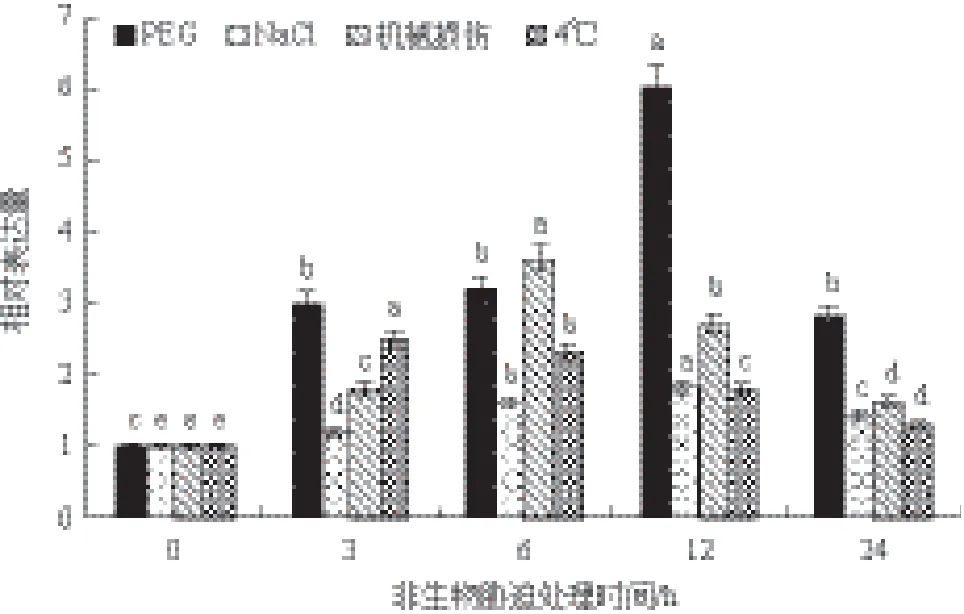
圖9 小桐子Jc5β-StR基因在不同非生物脅迫處理下的表達
類固醇化合物對植物生長發育過程具有調節作用,Mandava報道了甾和皂苷對某些植物的生長過程均有調節作用,特別是甾醇和性激素對一些植物的生長具有促進作用[18]。種子作為小桐子的生殖器官,在發育過程中大量合成和累積類固醇化合物,結合本實驗定量分析結果,Jc5β-StR在小桐子種子中高表達,并且在種子生長發育期表達量上升,種子成熟期(50 d)表達量達到最高值后下降,表達量變化過程與種子生長發育趨勢一致,進一步驗證了5β-StR在類固醇化合物生物合成途徑上的重要作用。保守結構域預測還表明Jc5β-StR也屬于SDRs家族一員,已報到的SDRs廣泛參與植物的初級代謝和次級代謝,在植物生長發育和抗逆機制中發揮重要作用[5-11]。本研究小桐子幼苗在PEG、NaCl、機械損傷和低溫這4種非生物脅迫處理下Jc5β-StR基因表達量都不同程度上調,由此推測Jc5β-StR也參與小桐子非生物脅迫應答。本研究為深入了解5β-StR基因的功能和小桐子的抗逆機制奠定了基礎。
4 結論
本研究報道了小桐子的一個內固醇5β還原酶基因 Jc5β-StR。序列全長1 173 bp,編碼390個氨基酸。該基因編碼的蛋白含有SDRs的6個保守結構域。Jc5β-StR具有組織表達特異性,在種子中表達量最高,還受多種非生物脅迫誘導上調表達。
[1]Jornvall H, Hedlund J, Bergman T, et al. Superfamilies SDR and MDR:from early ancestry to present forms. Emergence of three lines a Zn-metalloenzyme, and distinct variabilities[J]. Biochem Biophys Res Commun, 2010, 396:125-130.
[2]Moummou H, KallbergY, Tonfack LB, et al. The plant short chain dehydrogenase(SDR)superfamily:genome-wide inventory and diversification patterns[J]. BMC Plant Biol, 2012:12:1-17.
[3]Oppermann U, Filling C, Hult M, et al. Short-chain dehydrogenases/ reductases(SDR):the 2002 update[J]. Chem-Biol Interact, 2003, 143:247-253.
[4]Kallberg Y, Oppermann U, Persson B. Classification of the short-chain dehydrogenase/ reductase superfamily using hidden models[J]. Febs Journal, 2010, 277:2375-2386.
[5]楊東曉, 侯柄竹, 沈元月. 草莓果實FaSDR基因的克隆、生物信息學及表達分析[J]. 中國園藝文摘, 2015, 9:19-21.
[6]Li F, Asami T, Wu X, et al. A putative hydroxysteroid dehydrogenase involved in regulating plant growth and development[J]. PlantPhysiol, 2007, 145:87-97.
[7]Baud S, Dichow NR, Kelemen Z, et al. Regulation of HSDl in seeds of Arabidopsis thaliana[J]. Plant Cell Physiol, 2009, 50:1463-1478.
[8]陳全家, 劉超, 倪志勇, 等. 海島棉GbDET2基因的克隆及表達分析[J]. 分子植物育種, 2015, 13(1):119-124.
[9] Puyaubert J, Dieryck W, Costaglioli P, et al. Temporal gene expression of 3-ketoacyl-CoA reductase is different in high and in low erucic acid Brassica napes cultivars during seed development[J]. Biochim Biophys Acta, 2005, 1687:152-163.
[10] Choi HW, Lee BC, Kim NH, et al. A role for a reductase in resistance against microbial pathogens in plants[J]. Plant Physiol, 2008, 148:383-401.
[11] Herl V, Fischer G, Reva VA, et al. The VEP1 gene(At4g24220)encodes a short-chain dehydrogenase/reductase with 3-oxo-Δ4,5-steroid 5β -reductase activity in Arabidopsis thaliana L. [J]. Biochimie, 2009, 91:517-525.
[12]Edyta B, Marina K, Gabriele F, et al. Recombinant Δ4,5-steroid 5β -reductase as biocatalysts for the reduction of activated C=C-double bonds in monocyclic and acyclic molecules[J]. Adv Synth Catal, 2009, 351(17):2787-2790.
[13]余帥勇, 丁貴杰. 能源植物麻瘋樹研究進展[J]. 貴州林業科技, 2009, 37(1):49-54.
[14] 曾覺民. 可大力發展的生物質能源植物——膏桐[J]. 云南林業, 2006, 27(2):21-22.
[15]呂文, 王春蜂, 王國勝, 等. 中國林業生物質能源發展潛力的研究[J]. 中國能源, 2005, 27(11):21-26.
[16]林娟, 周選圍, 唐克軒, 等. 麻瘋樹植物資源研究概況[J].熱帶亞熱帶植物學報, 2004, 12(3):285-290.
[17]Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCTmethod[J]. Methods, 2001, 25:402-408.
[18]Mandava. Planta growth substances[J]. ACS symposium Seris, 1979, 160:164-165.
(責任編輯 狄艷紅)
Cloning and Expression Analysis of Jc5β-StR Gene in Jatropha curcas
HUANG Yao-yao1WEN Jin-fen2DENG Ming-hua3GONG Ming4CHEN Hao-wei11. Faculty of Modern Agricultural Engineering,Kunming University of Science and Technology,Kunming 650500;2. Faculty of Architecture and City Planning,Kunming University of Science and Technology,Kunming 650500;3. Faculty of Landscape and Horticulture,Yunnan
Agricultural University,Kunming 650201;4. School of Life Sciences,Yunnan Normal University,Kunming 650500)
The CDS sequence of Jc5β-StR gene was cloned with Jatropha curcas cDNA as template,and analyzed by bioinformatics analysis. The opening reading frame(ORF)of Jc5β-StR was 1 173 bp encoded 390 amino acids. The predicted molecular weight was 44.23 kD,and theoretical isoelectric point(pI)was 5.22. Blast searching and phylogenetic analysis showed that Jc5β-StR had highest identity(87%)and closest relationships with the 5β-StR of Ricinus communis. Jc5β-StR was a short-chain reductase,containing a progesterone 5β reductase(5β-POR)in a family of short chain reductase(SDRs),and also 6 conserved domains of SDRs. Jc5β-StR expressed in different tissues of root,stem,leave,flower,seed capsule,and seed,the 1highest in seed;moreover,the expression gradually rose with the development of seed,and declined after peak in the later stage of seed development(50 d). The abiotic stress such as NaCl,PEG,4℃,and mechanical damage resulted in the up-regulation of Jc5β-StR expression in varied level,indicating that Jc5β-StR involved in the response to abiotic stress.
Jatropha curcas L.;Jc5β-StR;clone;bioinformatics analysis;expression analysis
10.13560/j.cnki.biotech.bull.1985.2016-1114
2016-12-08
國家自然科學基金項目(31460355)
黃堯瑤,女,碩士研究生,研究方向:分子生物學;E-mail:358680047@qq.com
文錦芬,博士,副教授,研究方向:植物次生代謝產物;E-mail:wenjf888@163.com
猜你喜歡
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
山東工業技術(2016年15期)2016-12-01 05:31:22
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
